Хлорофилл: его свойства и биосинтез
Введение
Белки – высокомолекулярные азотосодержащие органические вещества, молекулы которых построены из остатков аминокислот. Белки составляют до половины и более сухой массы живой клетки.
Состав белков по химическим элементам: C, O, H, N, иногда S. В белках встречаются также элементы Fe, Cu, Zn и др. Но не химические элементы являются «буквами алфавита», из которых складывается всё многообразие «слов» – молекул белков. Такими структурными элементами белков природа выбрала простые соединения -- > >-аминокислоты.
Все белки состоят в основном из 20 > >-аминокислот.
По составу белки делятся на простые и сложные. Простые состоят только из аминокислотных остатков. Сложные белки отличаются от простых наличием простетической группы. Сложный белок, утративший простетическую группу, называют анобелком. Сложные белки подразделяют на классы в зависимости от состава и структуры простетической группы
название сложных белков простетическая группа
металлопротеины атомы металлов
гемопротеины железопорфирин
фосфопротеины фосфатные группы
гликопротеины алигосахариды, простые сахара
протеогликаны полисахариды
лигопротеины липиды
нуклеопротены ДНК или РНК
Белки относятся к высокомолекулярным соединения, в состав которых входят сотни и даже тысячи аминокислотных остатков, объединённых в макромолекулярную структуру. Молекулярная масса белков колеблется от 6000 до 1000000 дальтон и выше в зависимости от количества отдельных полипептидных цепей в составе единой молекулярной структуры белка. Такие полипептидные цепи получили название субъединиц. Их молекулярная масса варьируется в широких пределах: от 6000 до 100000 и более дальтон.
Хлорофилл
Хлорофилл относится к классу белков. Хлорофилл представляет собой «одно из интереснейших веществ на земной поверхности» (Ч. Дарвин), так как благодаря ему возможен синтез органических веществ из неорганических C> > и > >.
Важнейшую роль в процессе фотосинтеза играет зелёный пигмент – хлорофилл. Французские учёные Пелетье и Кавенту (1818) выделили из листьев зелёное вещество и назвали его хлорофиллом (от греч. «хлорос» – зелёный и «филлон» – лист). В настоящее время известно около 10 хлорофиллов. Они отличаются по химическому строению, окраске, распространению среди живых организмов. У всех высших растений содержатся хлорофиллы а и b. Хлорофилл с содержится в диатомовых водорослях, хлорофилл d – в красных водорослях. Кроме того, известны четыре бактериохлорофилла, содержащиеся в клетках фотосинтезирующих бактерий. В клетках зелёных бактерий содержатся бактериохлорофиллы с и d. В клетках пурпурных бактерий – бактериохлорофиилы а и b. Основными пигментами, без которых фотосинтез не идёт, являются хлорофилл а для зелёных растений и бактериохлорофилл для бактерий.
В первые точное представление и пигментах зелёного листа было получено благодаря работам крупнейшего русского ботаника М. С. Цвета. Он выделил пигменты листа в чистом виде и разработал новый хроматографический метод разделения веществ. Метод этот в дальнейшем получил широкое применение, как в биохимии, так и в чисто химических исследованиях.
Хлорофиллы а и b различаются по цвету. Хлорофилл а имеет сине-зелённый оттенок, а хлорофилл b – жёлто-зелёный. Содержание хлорофилла а в листе примерно в три раза больше по сравнению с хлорофиллом b.
Условия образования хлорофилла.
Образование хлорофилла осуществляется в 2 фазы: первая фаза – темновая, во время которой образуется предшественник хлорофилла – протохлорофилл, а вторая – световая, при которой из протохлорофилла на свету образуется хлорофилл. Для образования хлорофилла необходимо наличие железа. При недостатке железа получаются растения, характеризующиеся бледными полосами и слабой зелёной окраской листьев. Образование хлорофилла зависит от температуры. Оптимальная температура для накопления хлорофилла 26-30 С. Как и следовало ожидать, от температуры зависит лишь образование протохлорофилла (темновая фаза). При наличии уже образовавшихся протохлорофиллов процесс зеленения (световая фаза) идёт с одинаковой скоростью независимо от температуры. На скорость образования хлорофилла оказывает влияние содержания воды. Сильное обезвоживание проростков приводит к полному прекращению образования хлорофилла. Особенно чувствительно к обезвоживанию образование протохлорофилла.
Ещё В. И. Палладин обратил внимание на необходимость углеводов для протекания процесса зеленения. Именно с этим связано то, что зеленение проростков на свету зависит от их возраста. После 7-9-дневного возраста способность к образованию хлорофилла у таких проростков резко падает. При опрыскивании сахарозой проростки снова начинают интенсивно зеленеть.

Важнейшее значение для
образования хлорофилла имеют условия
минерального питания. Прежде всего,
необходимо достаточное количества
железа. При недостатке железа даже
листья взрослых растений теряют окраску.
Это явление названо хлорозом. Железо –
необходимый катализатор образования
хлорофилла. Оно необходимо на этапе
синтеза δ-аминолевулиновой
кислоты из глицерина и сукцинил-КоА, а
также синтеза протопорфирина. Большое
значение для обеспечения синтеза
хлорофилла имеет нормальное снабжение
растений азотом и магнием, так как оба
эти элемента входят в состав хлорофилла.
При недостатке меди хлорофилл легко
разрушается. Это, по-видимому, связано
с тем, что медь способствует образованию
устойчивых комплексов между хлорофиллом
и соответствующими белками.
Исследования процесса накопления хлорофилла у растений в течение вегетационного периода показало, что максимальное содержание хлорофилла приурочено к началу цветения. Есть даже мнение, что повышение образования хлорофилла может быть использовано как индикатор, указывающий на готовность растений к цветению. Синтез хлорофилла зависит от деятельности корневой системы. Так, при прививках содержание хлорофилла в листьях привоя зависит от свойств корневой системы подвоя. Возможно, что влияние корневой системы связано с тем, что там образуются гормоны (цитокинины). У двудомных растений большим содержанием хлорофилла характеризуются листья женских особей.
Химические свойства хлорофилла.
По химическому составу хлорофилл представляет сложный эфир дикарбоновой кислоты хлорофинилла. Хлорофинилл представляет собой азотосодержащее металлоорганическое соединение, относящееся к магний-порфиринам. В центре молекуле хлорофилла расположен атом магния, который соединён с четырьмя азотами пиррольных группировок. В пиррольных группировках хлорофилла имеется система чередующихся двойных и простых связей. Это и есть хромофорная группа хлорофилла, обуславливающая его окраску.
Наличие магния обнаруживается легко. Стоит только подействовать на спиртовую вытяжку хлорофилла слабым раствором соляной или какой-нибудь другой кислоты, чтобы определить магний. При этом произойдёт изменение окраски – вытяжка приобретает жёлто-бурый оттенок. Хлорофилл без магния получил название феофитина:
> >
В молекуле феофитина сравнительно легко ввести обратно какой-нибудь металл и восстановить металлоорганическую связь. Для этого к раствору феофитина прибавляют уксуснокислую медь или уксуснокислый цинк и нагревают. Цинк или медь входят в молекулу хлорофилла, и вытяжка становится опять зелёного цвета.
Химическая формула была установлена в 1913 году немецкими биохимиками Р. Вильштеттером и А. Штоллем. Им удалось её установить, последовательно отщепляя от молекулы хлорофилла отдельные её части действием кислот и щелочей, а в дальнейшем и нагреванием под давлением. До этих работ в физиологии растений считалось, что хлорофилл содержит железо, а не магний. Они же окончательно доказали и наличие двух хлорофиллов – а и b.
Эти же работы сделали образование кристаллического хлорофилла. Вильштеттер и Штолль показали, что имеющийся в зелёных листьях фермент хлорофиллаза отщепляет спирт фитол и на его место становится остаток этилового или метилового спирта. Такие соединения получили название хлорофиллидов. Если фитол замещается остатком этилового спирта, то полученное соединение называется этилхлорофиллидом.
Оптические свойства хлорофилла.
Хлорофилл поглощает солнечную энергию и направляет её на химические реакции, которые не могут протекать без энергии, получаемой извне. Раствор хлорофилла в проходящем свете имеет зелёный цвет, но при увеличении толщины слоя или концентрации хлорофилла он приобретает красный цвет.
Хлорофилл поглощает свет не сплошь, а избирательно. При пропускании из семи видимых цветов, которые постепенно переходят друг в друга. При пропускании белого света через призму и раствор хлорофилла на полученном спектре наиболее интенсивное поглощение будет в красных и сине-фиолетовых лучах. Зелёные лучи поглощаются мало, поэтому в тонком слое хлорофилл имеет в проходящем свете зелёный цвет. Однако с увеличением концентрации хлорофилла полосы поглощения расширяются (значительная часть зелёных лучей также поглощается) и без поглощения проходит только часть крайних красных. Спектры поглощения хлорофилла а и b очень близки.
В отражённом свете хлорофилл, кажется вишнёво-красным, так как он излучает поглощённый свет с изменением длины его волны. Это свойство хлорофилла называется флюоресценцией.
Биосинтез хлорофилла
Исходными для синтеза хлорофилла субстратами являются очень простые органические соединения – ацетат и глицин. Процесс синтеза хлорофилла принято подразделять на три этапа.
Первый этап состоит из следующих реакций:
Образование ацетилкофермента А, в котором принимает участие ацетат, кофермент А и АТФ. Реакция катализируется ацилкофермент А-синтетазой.
> >
Образование сукцинилкофермента А из двух молекул ацетилкофермента А. Считается не менее вероятным другой путь: вовлечение ацетата в цикл Кребса и образование в нём сукцината и затем сукцинилкофермента А. Некоторые исследователи считают исходным субстратом биосинтеза хлорофилла именно сукцинилкофермент А, не рассматривая реакции его образования (как не специфичные, осуществляющиеся в связи и с другими метаболическими цепочками).
> >
3. Образование > >-амино-> >-кетоадипиновой кислоты из сукцинилкофермента А и глицина, катализируемого так же, как и следующая реакция, ферментом синтетазой > >-аминолевулиновой кислоты:
> >
Из > >-амино-> >-кетоадипиновой кислоты путём декарбоксилирования образуется > >-аминолевулиновая кислота:
> >
Синтез из двух молекул > >-аминолевулиновой кислоты пирроленинового кольца и затем изомеризация его в пиррольное кольцо с образованием порфобилиногена. Реакция катализируется ферментом дегидразой > >-аминолевулиновой кислоты.
Второй этап включает реакции синтеза из четырёх пиррольных колец одной молекулы протопорфирина.
Из четырех молекул порфобилиногема под влиянием фермента порфобилиноге-дезаминазы синтезируется цепочка тетрапиррана.
Наименее изучен механизм реакции замыкания открытой цепи тетрапиррана в уропорфириноген III.
В результате декарбоксилирования всех четырёх остатков ацетата из уропорфириногена III образуется копропорфириноген III, фермент – уропорфириногендекарбоксилаза.
Происходит декарбоксилирование и дегидрирование двух из четырёх пропионатных остатков, что приводит к появлению винильных радикалов в кольцах и образованию протопорфириногена IX, фермент – копропорфириногендекарбоксилаза.
В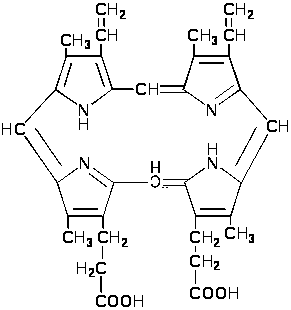
результате дегидрирования протопорфириногена
IX появляется протопорфирин
IX.
Третий этап характеризуется образованием и превращением магнийпорфиринов.
Протопорфирин, взаимодействуя с магнием, превращается в магнийпротопорфирин.
Д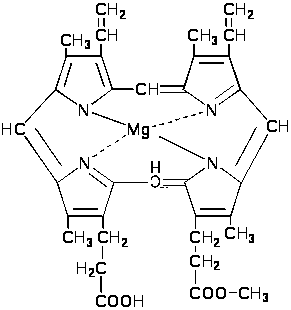
алее
следует ферментативная этерификация
метилом пропионатного остатка и
образование монометилового эфира
магний протопорфирина.
П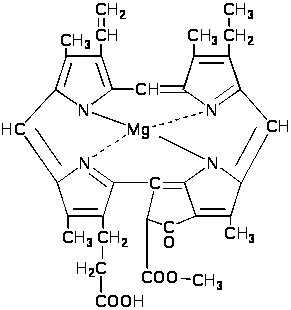
оследний
превращается в протохлорофиллид в
серии реакций, включающих преобразование
метильного производного пропионата.
П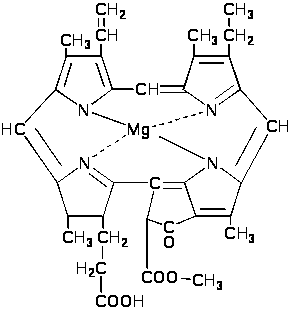
ревращение
протохлорофиллида в хлорофиллид,
заключающееся в гидрировании одной из
двух двойных связей кольца, осуществляется,
как правило, при участии света (в ходе
фотохимической реакции).
Лишь у некоторых низших и голосеменных растений эта реакция может протекать ферментативно в темноте. В хлорофиллид превращается не свободная форма протохлорофиллида, а связанная с белком в единый комплекс – так называемый протохлорофиллидголохром.
Последняя реакция – ферментативная этерификация хлорофиллида фитолом, в результате чего образуется хлорофилл а.
Поскольку синтез хлорофилла – многоэтапный, в нём участвуют различные ферменты, составляющие, по-видимому, полиферментный комплекс. Интересно заметить, что образование многих из этих белков-ферментов ускоряется на свету. Содержание хлорофилла в листе колеблется незначительно. Это связано с тем, что идёт непрерывный процесс разрушения старых молекул и образование новых молекул хлорофилла. Причём эти два процесса уравновешивают друг друга. При этом предполагается, что вновь образовавшиеся молекулы хлорофилла не смешиваются со старыми и имеют несколько иные свойства.
Различные виды хлорофилла.
Различные виды хлорофилла отличаются, как правило, природой заместителей при > >-атомах углерода в пиррольных кольцах порфиринов. Все связи > >-углеродных атомов заняты в формировании порфиринового кольца и поэтому не могут определять специфику отдельных видов хлорофилла. Хлорофиллы бактерий называют бактериохлорофиллами. Их известно четыре. Большинство пурпурных бактерий содержит бактериохлорофилл а, который и обуславливает их способность к фотосинтезу:
Этот пигмент представляет собой порфирин, у которого > >-углеродные атомы имеют следующих заместителей в положениях: 1 – метил, 2 – ацетил, 3 – метил, 4 – этил, 5 – метил, 6-й атом углерода участвует в образовании насыщенного циклопентанового кольца, 7 – сложный эфир пропионовой кислоты и высокомолекулярного ненасыщенного спирта фитола, 8 – метил. Кольца В и D имеют лишь по одной двойной связи. В циклопентановом кольце в положении 9 – кетогруппа, а в 10 – карбоксиметильный радикал > >. Фитол может рассматриваться как производное изопрена или дитерпена с одной двойной связью.
Бактериохлорофиллы с и d зелёных бактерий отличаются от бактериохлорофилла а рядом особенностей. Они не имеют циклопентановного кольца. Карбоксиметильный радикал при атоме 10 отсутствует, а атомы 9 и 10 входят в состав пропила. Вместо фитольного остатка содержат фарнезильный -- > >.
При втором углеродном атоме порфирин имеется вместо ацетильного оксиэтильный радикал > >. У некоторых зелёных бактерий имеется несколько процентов (до 10) бактериохлорофилла а.
Все другие фотосинтезирующие организмы содержат в качестве основного зелёного пигмента хлорофилл а.
От бактериохлорофилла а он отличается тем, что в положении 2 имеет винильный радикал > >, а кольцо В имеет ещё одну двойную связь (за счёт того, что содержит на 2 атома водорода меньше).
Кроме хлорофилла а, все высшие растения и большинство водорослей (за исключением сине-зелёных и красных) содержат хлорофилл. Он отличается от хлорофилла а тем, что при углеродном атоме 3 вместо метильной имеется формильная группа СНО.
У бурых и хризофитовых водорослей, а также динофлагеллят обнаружен хлорофилл с, не имеющий остатка фитола.
Красные водоросли имеют хлорофилл, который отличается от хлорофилла а тем, что вместо винильной группы при углеродном атоме 2 имеется формильный радикал.
Распознавание различных видов хлорофилла осуществляется с помощью спектральных характеристик. Обычно изучают спектр поглощения растворов пигментов, реже – спектр люминесценции.
хлорофилл а хлорофилл b
Хлорофилл d
Хлорофилл а
Хлорофилл b
Министерство высшего и профессионального образования Российской Федерации
Тверской Государственный Технический Университет
Кафедра информатики и прикладной математики
Курсовая работа
«Хлорофилл: его свойства и биосинтез»
Выполнил: студентка группы БТ-111
Николаева Е. С.
Проверил: Семёнов А. В.
Принял: Семёнов А. В.