Ретикулярная формация ствола мозга, вегетативная нервная система
МИНИСТЕРСТВО ЗДРАВООХРАНЕНИЯ УКРАИНЫ
ЛУБЕНСКОЕ МЕДИЦИНСКОЕ УЧИЛИЩЕ
РЕФЕРАТ
по ФИЗИОЛОГИИ
НА ТЕМУ: Ретикулярная формация ствола мозга, вегетативная нервная система. Кора большого мозга
Выполнила: студентка группы Ф-31
Бендрик Татьяна
Лубны 2009
Ретикулярная формация ствола мозга
Особенности строения. Ретикулярная формация представляет собой скопление особых нейронов, волокна которых образуют своеобразную сеть. Нейроны ретикулярной формации в области ствола мозга были описаны в прощлом веке немецким ученым Дейтерсом. В. М. Бехтерев подобные же структуры обнаружил в области спинного мозга. Нейроны ретикулярной формации образуют скопления, или ядра. Дендриты этих клеток относительно длинные, маловетвистые, аксоны, напротив, короткие, имеют много ответвлений (коллатералей). Эта особенность обусловливает многочисленные синаптические контакты нейронов ретикулярной формации.
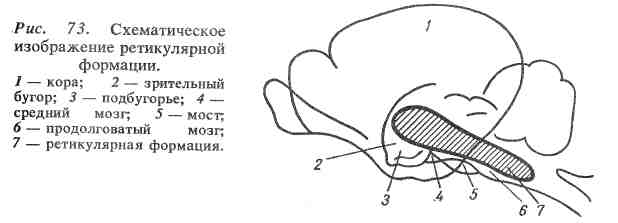
Ретикулярная формация ствола мозга занимает центральное положение в продолговатом мозге, мосту мозга, среднем и промежуточном мозге (рис. 73).
Нейроны ретикулярной формации не имеют непосредственных контактов с рецепторами организма. Нервные импульсы при возбуждении рецепторов поступают к ретикулярной формации по коллатералям волокон вегетативной и соматической нервной системы.
Физиологическая роль. Ретикулярная формация ствола мозга оказывает восходящее влияние на клетки коры большого мозга и нисходящее на мотонейроны спинного мозга. Эти влияния могут быть активирующими или тормозными.
Афферентные импульсы к коре большого мозга поступают по двум путям: специфическому и неспецифическому. Специфический нервный путь обязательно проходит через зрительные бугры и несет нервные импульсы к определенным зонам коры головного мозга, в результате осуществляется какая-либо специфическая деятельность.
Например, при раздражении фоторецепторов глаз импульсы через зрительные бугры поступают в затылочную область коры большого мозга и у человека возникают зрительные ощущения.
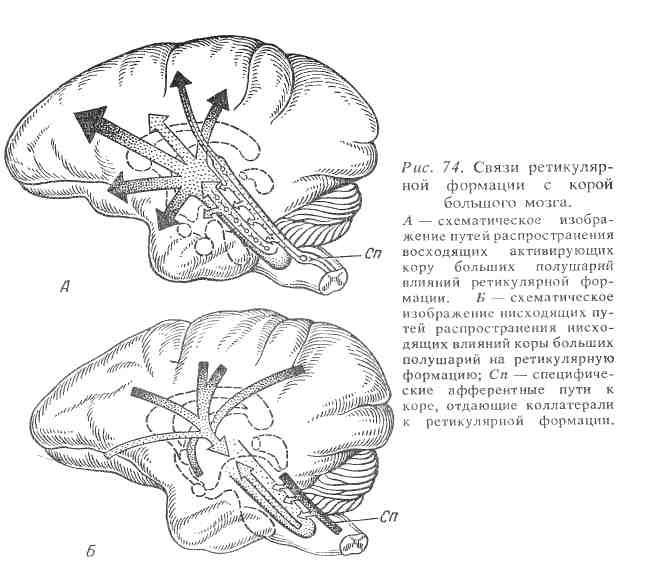
Неспецифический нервный путь обязательно проходит через нейроны ретикулярной формации ствола мозга. Импульсы к ретикулярной формации поступают по коллатералям от специфического нервного пути. Благодаря многочисленным синапсам на одном и том же нейроне ретикулярной формации могут сходиться (конвергировать) импульсы различных значений (световые, звуковые и т. д.), при этом они теряют свою специфичность. От нейронов ретикулярной формации эти импульсы поступают не в какую-то определенную область коры большого мозга, а веерообразно распространяются по ее клеткам, повышая их возбудимость и облегчая тем самым выполнение специфической функции (рис. 74).
В спинном мозге, как уже указывалось, также имеются нейроны ретикулярной формации. Полагают, что они поддерживают на высоком уровне активность нейронов спинного мозга. Функциональное состояние самой ретикулярной формации регулируется корой большого мозга.
Мозжечок
Особенности строения мозжечка. Связи мозжечка с другими отделами центральной нервной системы. Мозжечок — непарное образование; он располагается позади продолговатого мозга и моста мозга, граничит с пластинкой крышки, сверху прикрыт затылочными долями больших полушарий.
Функции мозжечка были изучены на животных, у которых мозжечок удаляли частично или полностью, а также путем регистрации его биоэлектрической активности в покое и яри раздражении.
Двигательные расстройства при удалении мозжечка были описаны известным итальянским физиологом Лючиапи. Основными из них являются: атония — исчезновение или ослабление мышечного тонуса; астения — снижение силы мышечных сокращений (для такого животного характерно быстрое наступление мышечного утомления); астазия — потеря способности к слитным тетаническим сокращениям. У животных наблюдаются дрожательные движения конечностей и головы. Собака после удаления мозжечка не может сразу поднять лапы, животное делает ряд колебательных движений лапой, перед тем как ее поднять. Если поставить такую собаку, то тело ее и голова все время качаются из стороны в сторону.
В результате атонии, астении и астазии у животного нарушается координация движений: отмечается шаткая походка, размашистые, неловкие, неточные движения. Весь комплекс двигательных расстройств при поражении мозжечка получил название мозжечковой атаксии (рис. 75).
Подобные нарушения наблюдаются и у человека при поражении мозжечка.
Через некоторое время после удаления мозжечка все двигательные расстройства постепенно сглаживаются. Если у таких животных удалить моторную область коры большого мозга, то двигательные нарушения вновь усиливаются. Следовательно, компенсация (восстановление) двигательных расстройств при поражении мозжечка осуществляется при участии коры большого мозга, ее моторной области.
Исследованиями Л. А. Орбели показано, что при удалении мозжечка наблюдается не только падение мышечного тонуса (атония), но и неправильное его распределение (дистония).
Л. А. Орбели установил, что мозжечок
влияет и на состояние рецепторного
аппарата, а также на вегетативные
процессы. Мозжечок оказывает
адаптационно-трофическое влияние на
все отделы мозга через симпатическую
нервную систему, он регулирует обмен
веществ в головном мозге и тем самым
способт

 вует
приспособлению нервной системы к
изменяющимся у слови ям существования.
вует
приспособлению нервной системы к
изменяющимся у слови ям существования.
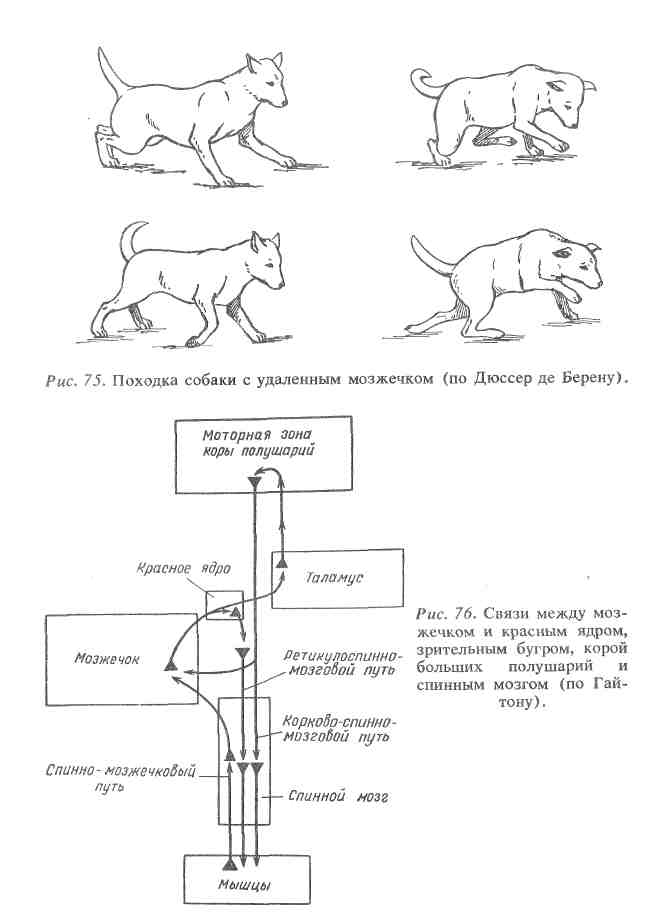
Таким образом, основными функциями мозжечка являются координация движений, нормальное распределение мышечного тонуса и регуляция вегетативных функций. Свое влияние мозжечок реализует через ядерные образования среднего и продолговатого мозга, а также двигательные нейроны спинного мозга. Большая роль в осуществлении влияний мозжечка на функции организма принадлежит двусторонней связи мозжечка с моторной зоной коры большого мозга и ретикулярной формацией ствола мозга (рис. 76).
Вегетативная нервная система
Нервную систему организма животных и человека подразделяют на соматическую (анимальную) и вегетативную.
Соматическая нервная система обеспечивает иннервацию поперечнополосатой мускулатуры и восприятие раздражений.
Вегетативная нервная система обеспечивает иннервацию внутренних органов, желез внешней и внутренней секреции, кровеносных и лимфатических сосудов, трофическую иннервацию (регулирует обмен веществ) скелетной мускулатуры, рецепторов и самой центральной нервной системы.
Анатомические особенности. Вегетативная нервная система имеет центральную и периферическую части. Центральные части расположены очагово, в виде скопления нейронов (ядер вегетативной нервной системы), заложенных в спинном, продолговатом и среднем мозге.
Вегетативная нервная система имеет два отдела: парасимпатический и симпатический.
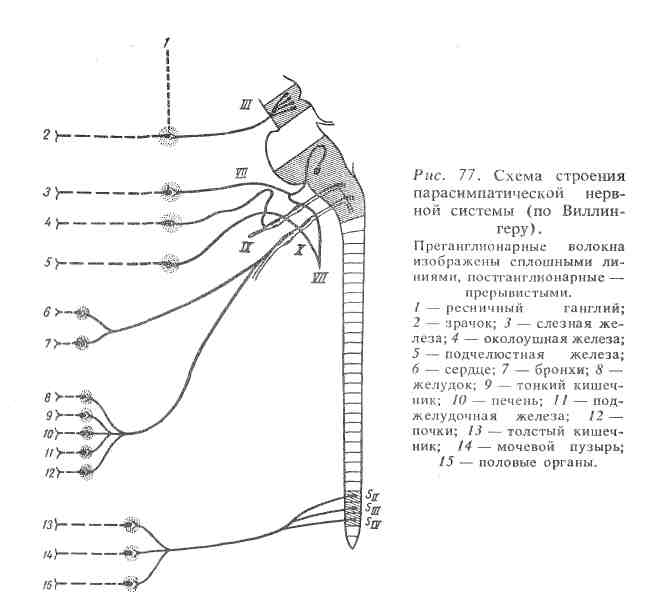
Ядра, находящиеся в среднем мозге (III пара черепных нервов), продолговатом мозге (VII, IX и X пары черепных нервов) и крестцовом отделе спинного мозга (ядра тазовых внутренних нервов), являются центрами парасимпатической нервной системы (рис. 77).
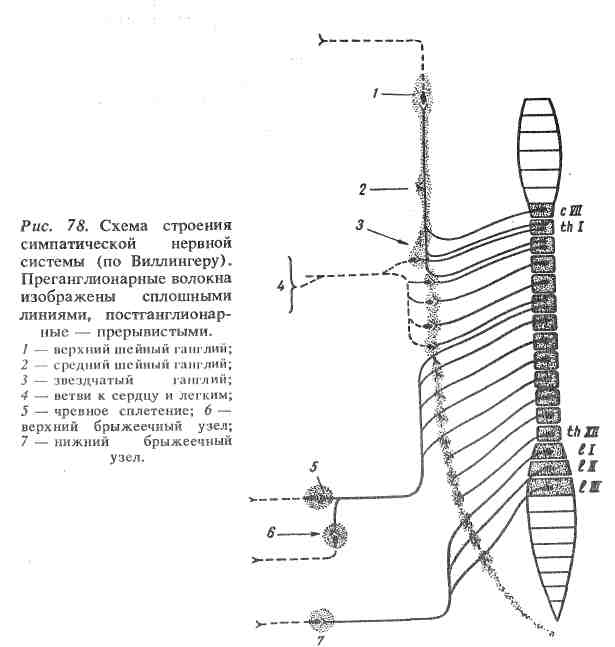
Ядра, расположенные в боковых рогах грудного и поясничного отделов спинного мозга (от I грудного до II— IV поясничного), образуют центры симпатической нервной системы (рис. 78).
От нейронов вегетативной нервной системы, расположенных в центральной нервной системе, отходят на периферию нервные волокна, которые, не дойдя до иннервируемого органа, прерываются в вегетативных ганглиях. Здесь они образуют многочисленные синапсы на нервных клетках ганглия. Нервные волокна, подходящие к ганглию, называются преганглионарными. Нервные отростки, отходящие от ганглиозных клеток, образуют постганглионарн ы е нервные волокна, которые достигают иннервируемого органа.
Ганглии парасимпатической нервной системы располагаются внутри иннервируемого органа или вблизи нег. Ганглии симпатической нервной системы находятся в отдалении от иннервируемых ими органов. Ганглии симпатической нервной системы образуют так называемую симпатическую цепочку, располагающуюся справа и слева от позвоночного столба, и ряд узлов на более далеком расстоянии от него (чревное сплетение, верхний и нижний брыжеечные узлы).
Физиологические особенности. Волокна нервов вегетативной нервной системы в 2—5 раз тоньше волокон соматических нервов. Отсюда различная скорость проведения нервных импульсов. В соматических нервах нервные импульсы распространяются со скоростью до 120—140 м/сек, в парасимпатических— 10—20 м/сек, в симпатических — 0,4—0,5 м/сек. Волокна нервов вегетативной нервной системы менее возбудимы и обладают более продолжительным рефрактерным периодом, чем соматические нервы, поэтому для возбуждения вегетативных нервов необходимо более сильное раздражение.
Парасимпатическая нервная система. От среднего мозга отходят парасимпатические волокна, которые входят в состав глазодвигательного нерва. Эти волокна интернируют круговую мышцу радужной оболочки глаза, при их возбуждении происходит уменьшение просвета зрачка. Из продолговатого мозга выходят парасимпатические волокна, идущие в составе лицевого, языкоглоточного и блуждающего нервов. Парасимпатические волокна, входящие в состав лицевого и языкоглоточного нервов, иннервируют слюнные железы. При возбуждении этих волокон наблюдается обильное выделение слюны. Блуждающий нерв, разветвляясь, иннервирует многие внутренние органы: сердце, пищевод, бронхи, альвеолы легких, желудок, тонкий кишечник и верхний отдел толстого, поджелудочную железу, надпочечники, почки, печень, селезенку. От крестцового отдела спинного мозга отходят волокна тазовых внутренних нервов, которые иннервируют органы малого таза: сигмовидную и прямую кишку, мочевой пузырь, половые органы, за исключением матки.
Симпатическая нервная система. Волокна симпатической нервной системы начинаются от нейронов боковых рогов грудного и поясничного отделов спинного мозга. Симпатическая нервная система иннервирует все органы и ткани организма, в том числе скелетные мышцы и центральную нервную систему.
Симпатический и парасимпатический отделы вегетативной нервной системы оказывают на органы, как правило, противоположное влияние. Например, при возбуждении парасимпатических (блуждающих) нервов ритм сердца замедляется, под влиянием симпатических нервов ускоряется. При повышении активности блуждающих нервов тонус гладкой мускулатуры бронхов повышается, в результате этого просвет их уменьшается. Под влиянием симпатической нервной системы мускулатура бронхов расслабляется и просвет их увеличивается. За счет разнонаправленного влияния двух отделов вегетативной нервной системы на деятельность органов обеспечивается лучшее приспособление организма к условиям существования.
За счет парасимпатической нервной системы осуществляются рефлекторные реакции защитного характера (сужение зрачка при вспышке яркого света); рефлекторные реакции, направленные на сохранение состава и свойств внутренней среды организма (возбуждение блуждающего нерва стимулирует процессы пищеварения и тем самым обеспечивает восстановление уровня питательных веществ в организме). Активация парасимпатического отдела вегетативной нервной системы способствует опорожнению полых органов (желчного пузыря, мочевого пузыря, прямой кишки).
Возбуждение симпатической нервной системы обеспечивает поддержание постоянства состава внутренней среды организма (гомеостаза). Например, при понижении уровня сахара в крови возбуждение симпатической нервной системы приводит к увеличению активности мозгового вещества надпочечников и выделению адреналина. Током крови гормон доставляется к печени и способствует переходу гликогена в глюкозу, которая поступает в кровь, и уровень сахара восстанавливается.
Симпатическая нервная система не только регулирует работу внутренних органов, но и оказывает влияние на обменные процессы, протекающие в скелетных мышцах и в нервной системе, что было впервые установлено Л. А, Орбели (адаптационно-трофическая функция симпатической нервной системы).
Под адаптационно-трофической функцией симпатической нервной системы следует понимать ее влияние на интенсивность обменных процессов и приспособление их уровня к условиям существования организма.
В лаборатории Л. А. Орбели на нервно-мышечном препарате лягушки был проведен следующий опыт. Получали тетаническое сокращение мышцы посредством раздражения двигательного нерва. Продолжая раздражать нерв, доводили мышцу до степени утомления. Раздражение в этих условиях симпатического нерва восстанавливало работоспособность скелетной мышцы. Она вновь приобретала способность реагировать тетаническим сокращением на раздражение двигательного нерва.
В настоящее время установлено, что при возбуждении и торможении всех отделов центральной и периферической нервной системы происходит образование физиологически активных веществ — медиаторов.
В зависимости от того, какой медиатор образуется в окончаниях нервных волокон, принято делить их на холинергические и адренергические. Передача возбуждения в холинергических нервных волокнах осуществляется при помощи ацетилхолина, а в адренергических — норадреналина. Холинергическими являются все преганглионарные нервные волокна (парасимпатические и симпатические), все постганглионарные нервные волокна парасимпатической нервной системы и соматические нервы. Адренергическими являются все постганглионарные симпатические нервы, за исключением нервов потовых желез и симпатических нервов, расширяющих кровеносные сосуды.
Холинергические и адренергические нейроны обнаружены и в центральной нервной системе.
Рецепторы, взаимодействующие с ацетилхолином,
называют холинорецепторами, взаимодействующие с норадреналином — адренорецепторами. Медиатор изменяет структуру молекулы белка рецептора, что приводит к повышению проницаемости постсинаптической мембраны, изменению движения через нее ионов. Вследствие этого в постсинаптической мембране возникает деполяризация или гиперполяризация. Если происходит деполяризация постсинаптической мембраны и этот процесс достигает достаточного (критического) уровня, возбуждение передается на эффекторную клетку. Если же в результате взаимодействия медиатора с рецептором возникает процесс гиперполяризации постсинаптической мембраны, передача возбуждения, тормозится.
После того как медиатор передал возбуждение, он разрушается специфическим ферментом.
Кора большого мозга
Особенности строения коры большого мозга и методы изучения ее функций. Кора большого мозга является высшим и наиболее молодым отделом центральной нервной системы.
У взрослого человека толщина коры в большинстве областей составляет около 3 мм. Площадь ее благодаря многочисленным складкам и бороздам составляет 0,25 м2. Кора состоит из 14—17 млрд. нервных клеток, их отростков и нейроглии; для большинства ее участков характерно шестислойное расположение нейронов.
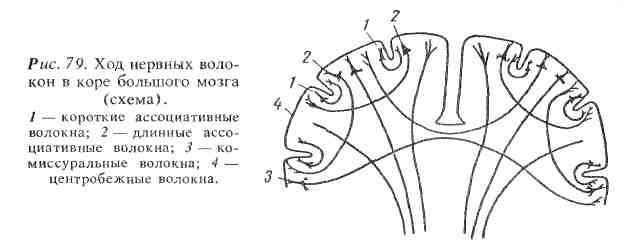
Отростки нервных клеток, коры большого мозга связывают различные ее отделы между собой или с нижележащими отделами центральной нервной системы. Отростки нервных клеток, соединяющие между собой различные участки одного и того же полушария, называются ассоциативными, связывающие чаще всего одинаковые участки двух полушарий — к о м и с-суральными и обеспечивающие контакты коры большого мозга с другими отделами центральной нервной системы и через них со всеми органами и тканями тела — проводящими (центробежными). Схема этих путей приведена на рис. 79.
Клетки нейроглии выполняют ряд важных функций: они являются опорной тканью, участвуют в обмене веществ головного мозга, регулируют кровоток внутри мозга, выделяют нейросекрет, который регулирует возбудимость нейронов коры большого мозга.
Для изучения функций коры большого мозга применяются различные методы: 1) удаление отдельных участков коры больших полушарий оперативным путем. После исчезновения последствий хирургической травмы наблюдают, какие функции в организме нарушаются, а какие — сохраняются; 2) метод раздражения с использованием электрических, химических и температурных раздражителей. Как и предыдущий, этот метод позволяет определить роль различных участков коры в регуляции функций организма; 3) метод отведения биопотенциалов от отдельных зон и нейронов коры. Метод позволяет регистрировать электрическую активность не только поверхностных, но и глубоких структур головного мозга, а также ее изменения под влиянием различных афферентных раздражений; 4) классический метод условных рефлексов, разработанный И. П. Павловым. Достоинство этого метода состоит в возможности исследования высшей нервной деятельности на здоровых животных и людях; 5) клинический метод, позволяющий изучать изменения деятельности отдельных органов и их систем, которые наблюдаются при повреждении коры большого мозга (кровоизлияния, ранения, опухоли мозга).
Функции коры большого мозга: 1) кора осуществляет взаимодействие организма с окружающей средой за счет безусловных и условных рефлексов; 2) она является основой высшей нервной деятельности (поведения) организма; 3) за счет деятельности коры осуществляются высшие психические функции: мышление и сознание; 4) кора регулирует и объединяет работу всех внутренних органов и регулирует обмен веществ.
Таким образом, с появлением: коры большого мозга она начинает контролировать все процессы, протекающие в организме, а также всю деятельность человека, то есть происходит кортиколизация функций. И. П. Павлов, характеризуя значение коры, указывал, что она является распорядителем и распределителем всей деятельности животного и человеческого организма.
Функциональное значение различных областей коры большого мозга. Локализация функций. Роль отдельных областей коры впервые была изучена в 1870 г. немецкими исследователями Фричем и Гитцигом. Они показали, что раздражение различных участков передней центральной извилины и собственно лобных долей вызывает сокращение определенных групп мышц на противоположной раздражению стороне. В дальнейшем была выявлена функциональная неоднозначность различных областей коры. Обнаружено, что височные доли коры связаны со слуховыми функциями, затылочные — со зрительными и т. д. Эти исследования позволили сделать вывод, что разные участки коры больших полушарий ведают определенными функциями. Было создано учение о локализации функций в коре большого мозга.
И. П. Павлов, сочетая у экспериментальных животных (собак) метод экстирпации (удаление) отдельных участков коры большого мозга с методом условных рефлексов, подтвердил основные положения теории о локализации функций в коре. Вместе с тем ряд положений этой теории И. П. Павлов уточнил и внес в нее принципиально новые представления.
По И. И. Павлову деление коры большого мозга на двигательные и воспринимающие (чувствительные, или сенсорные) зоны является неправильным. Метод- условных, рефлексов позволил установить, что вся кора обладает способностью осуществлять анализ и синтез афферентных импульсов, поступающих от различных рецепторов, воспринимающих раздражения внешнего мира и внутренней среды организма. И. П. Павлов показал, что выпадающая функция при удалении участков коры может быть в какой-то степени восстановлена за счет деятельности оставшихся отделов коры большого мозга.
Представления И. П. Павлова о локализации функций в коре большого мозга подтверждаются клиническими наблюдениями. Функции, выпавшие при локальных поражениях мозга (кровоизлияниях, ранениях, опухолях) могут частично или полностью восстанавливаться. Кроме того, результаты электрофизиологических исследований позволяют регистрировать характерные изменения биоэлектрической активности в определенных участках коры при раздражении рецепторов. Моторная зона представлена передней центральной извилиной и расположенными вблизи нее участками лобной области. При ее раздражении возникают разнообразные сокращения скелетной мускулатуры на противоположной стороне. Эта область коры особенно развита у обезьян и человека. Установлено соответствие между определенными зонами передней центральной извилины и скелетной мускулатурой (рис. 80).
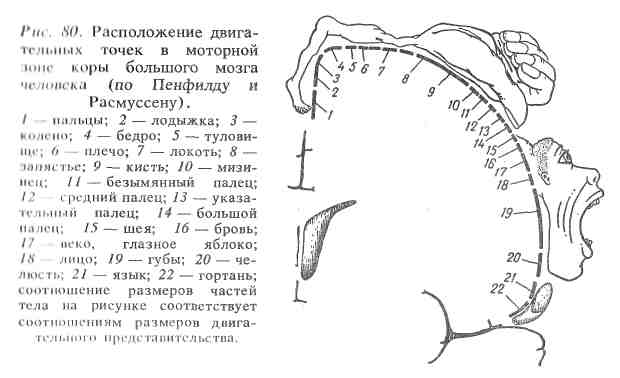
Верхние участки этой зоны связаны с рецепторами мышц нижних конечностей, средние — мышц туловища, нижние — мышц головы.
В зависимости от обширности поражения передней центральной извилины наступают параличи (полное отсутствие движений) или парезы (ослабление движений).
Особый интерес представляет собственно лобная область, которая достигает у человека наибольшего развития. При поражении лобных областей у человека нарушаются сложные двигательные функции, обеспечивающие трудовую деятельность и речь, а также приспособительные поведенческие реакции организма.
Область кожной чувствительности представлена в основном задней центральной извилиной. Клетки этой области воспринимают импульсы от тактильных, болевых и температурных рецепторов кожи. Проекция кожной чувствительности в пределах задней центральной извилины аналогична таковой для двигательной зоны, Верхние участки задней центральной извилины связаны с рецепторами кожи нижних конечностей, средние — с рецепторами кожи туловища и рук, нижние — с рецепторами кожи головы и лица. Раздражение этой области у человека во время нейрохирургических операций вызывает ощущения прикосновения, покалывания, онемения, при этом никогда не наблюдается выраженных болевых ощущений.
Поражение области задней центральной извилины на одной стороне приводит к нарушению кожной чувствительности на противоположной стороне тела. При двустороннем повреждении указанной зоны коры большого мозга наблюдается полная потеря чувствительности (анестезия).
Зрительная область расположена в затылочных долях коры большого мозга обоих полушарий. Эту область следует рассматривать как проекцию сетчатой оболочки глаза.
При поражении затылочной области может нарушиться зрительная память, ориентация в непривычной обстановке и развиваться полная корковая слепота.
Слуховая область локализуется в височных долях коры большого мозга. Сюда поступают нервные импульсы от рецепторов улитки внутреннего уха. При повреждении этой зоны может возникнуть музыкальная и словесная глухота, когда человек слышит, но не понимает значения слов. Двустороннее поражение слуховой области приводит к полной корковой глухоте.
Вкусовая область расположена в лимбической системе (крючок) коры большого мозга. Эта область получает нервные импульсы от вкусовых рецепторов слизистой оболочки полости рта. Поражение этой зоны приводит к потере или искажению вкусовых ощущений.
Обонятельная область располагается в лимбической системе (крючок, гиппокамп) коры большого мозга. Сюда поступают нервные импульсы от обонятельных рецепторов слизистой оболочки носа. Повреждение этой зоны ведет к понижению или потере обоняния.
В коре больших полушарий обнаружено несколько зон, ведающих функцией речи. В лобной области левого полушария располагается, моторный центр речи (центр Брока). У леворуких он локализуется в правом полушарии. При поражении этого центра речь затруднена или даже невозможна. В височной области находится
сенсорный центр речи (центр Вернике). Повреждение этой области приводит к расстройствам восприятия речи: больной не понимает значение слов, хотя способность произносить слова сохранена. В затылочной доле коры большого мозга имеются зоны, обеспечивающие восприятие письменной (зрительной) речи. При поражении этих областей больной не понимает письменный текст.
Теменную область коры большого мозга относят к ассоциативным зонам. Среди нервных клеток теменной области найдено большое количество полисенсорных нейронов, которые способствуют установлению связей между различными зонами коры больших полушарий и играют большую роль в формировании условных рефлексов.
Любая область коры большого мозга находится и в анатомическом, и в функциональном контакте с другими Зонами коры, с подкорковыми ядрами, с образованиями промежуточного мозга и ретикулярной формации, что обеспечивает совершенство выполняемых ими функций.
Использованная литература:
1. Физиология / Под ред. С.А. Георгиевой. – 2-е изд. – Ф48 М.: Медецина, 1986. – 400 с.