Механизмы теплопродукции и теплоотдачи
СОДЕРЖАНИЕ
стр.
Введение ………………………………………………………………….3
Пойкилотермия, гетеротермия, гомойотермия ………………………...4
Принципы регуляции температуры тела, тепловой баланс …………...5
Физиология теморецепторов ……………………………………………6
Центры терморегуляции ………………………………………………...8
а) центры теплоотдачи ………………………………………………...9
б) центры теплопродукции …………………………………………..10
6) Механизмы теплопродукции …………………………………………..10
а) сократительный термогенез ………………………………………11
б) несократительный термогенез ……………………………………12
7) Механизмы теплоотдачи ……………………………………………….12
а) теплопроведение …………………………………………………...13
б) теплоизлучение …………………………………………………….13
в) конвекция …………………………………………………………..14
г) испарение …………………………………………………………..14
8) Обмен веществ ………………………………………………………….16
9) Питание ………………………………………………………………….17
10) Заключение ……………………………………………………………...20
11) Список используемой литературы ……………………………………..23
ВВЕДЕНИЕ
Как бы ни были разнообразны формы проявления жизни, они всегда неразрывно связаны с превращением энергии. Энергетический обмен является особенностью, присущей каждой живой клетке. Богатые энергией питательные вещества усваиваются и химически преобразуются, а конечные продукты обмена веществ с более низким содержанием энергии выделяются из клетки. Согласно первому закону термодинамики, энергия не исчезает и не возникает вновь. Организмы должны получать энергию в доступной для них форме из окружающей среды и возвращать в среду соответствующее количество энергии в форме, менее пригодной для дальнейшего использования.
Около столетия тому назад французский физиолог Клод Бернар установил, что живой организм и среда образуют единую систему, гак как между ними происходит непрерывный обмен веществами и энергией. Нормальная жизнедеятельность организма поддерживается регуляцией внутренних компонентов, требующей затраты энергии. Использование химической энергии в организме называют энергетическим обменом: именно он служит показателем общего состояния и физиологической активности организма.
Обменные (или метаболические) процессы, в ходе которых специфические элементы организма синтезируются из поглощенных пищевых продуктов, называют анаболизмом; соответственно те метаболические процессы, в ходе которых структурные элементы организма или поглощенные пищевые продукты подвергаются распаду, называют катаболизмом.
Живой организм продуцирует тепло, которое идет на нагревание тела. Удельная теплоемкость тела человека (количество тепла, необходимое для нагревания ткани на 1°С) равна в среднем 0,83 ккал/кг на 1 градус (для воды — 1 ккал/кг на градус). Чтобы повысить температуру тела человека массой 70 кг на 1°, следует затратить 58,1 ккал (0,83•70). В среднем человек массой 70 кг в условиях покоя выделяет около 72 ккал/час. Следовательно, если бы не было второго процесса — теплоотдачи, то ежечасно ткани человека нагревались бы на 1,24° (72:58,1). Однако такого не происходит, так как в норме в условиях покоя скорость продукции тепла равна скорости ее потери. Это носит название теплового баланса, в основе которого лежат процессы регуляции теплопродукции и теплоотдачи. Все вместе это называется терморегуляция.
ПОЙКИЛОТЕРМИЯ, ГЕТЕРОТЕРМИЯ, ГОМОЙОТЕРМИЯ
В эволюции системы терморегуляции имеется нижняя ступень, на которой температура тела животного зависит в основном от температуры среды: когда она уменьшается, температура тела тоже падает и наоборот. Такое состояние температуры тела получило название пойкилотермия, а животные — пойкилотермные. Типичным представителем пойкилотермных является лягушка. Зимой температура тела лягушки приближается к нулю. В этом состоянии она все же способна совершать прыжки в длину, но не более 12— 15 см. Летом температура тела ее достигает 20—25°С, а прыгать она может значительно дальше — до 1 м. Обычно в условиях низкой температуры пойкилотермные животные впадают в состояние анабиоза. Существуют микроорганизмы, для которых оптимум температуры среды варьирует от 0°С до минус 60°С, например, микробы, живущие в толще льда, или, наоборот, микроорганизмы, выдерживающие температуру среды от +70°С до + 120°С, например, микробы горячих источников.
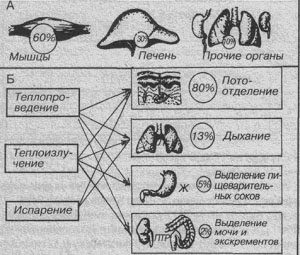
Механизмы теплопродукции и теплоотдачи.
А – роль органов в теплопродукции
Б – роль органов в теплоотдаче
Ряд животных, например, летучая мышь, грызуны, некоторые виды птиц, к примеру, колибри, относится к группе гетеротермных организмов: при одних условиях они пойкилотермные организмы, при других – гомойотермные.
Млекопитающие относятся к гомойотермным организмам (теплокровным), у которых имеет место изотермия, или постоянство температуры организма. Однако изотермия имеет относительный характер: температура тканей, расположенных не глубже 3 см от поверхности тела (кожа, подкожная клетчатка, поверхностные мышцы), или оболочки, — во многом зависит от внешней температуры, в то время как ядро тела, т. е. ЦНС, внутренние органы, скелетные мышцы, расположенные глубже 3 см, имеют сравнительно постоянную температуру, независимо от температуры окружающей среды. Таким образом, теплокровные имеют пойкилотермную оболочку и гомойотермную «сердцевину», или «ядро».
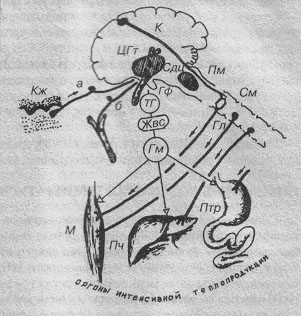
Органы теплопродукции и управление выработкой тепла.
К – кора, Кж – кожа, ЦГт – центры гипоталамуса, Сдц – сосудодвигательный центр, Пм – продолговатый мозг, См – спинной мозг, Гф – гипофиз, ТГ – тиреотропный гормон, Жвс – железы внутренней секреции, Гм – гормоны, М – мышца, Пч – печень, Птр – пищеварительный тракт, а, б – поток дифферентной импульсации.
У человека средняя температура мозга, крови, внутренних органов приближается к 37°С. Физиологический предел колебаний этой температуры составляет 1,5°. Изменение температуры крови и внутренних органов у человека на 2—2,5°С от среднего уровня сопровождается нарушением физиологических функций, а температура тела выше 43°С практически несовместима с жизнью человека.
ПРИНЦИПЫ РЕГУЛЯЦИИ ТЕМПЕРАТУРЫ ТЕЛА,
ТЕПЛОВОЙ БАЛАНС
Температура ядра (тела) определяется двумя потоками — теплообразованием (теплопродукцией) и теплоотдачей (тепловыделением). При термонейтральной, или комфортной зоне (при 27—32°С), существует баланс между теплопродукцией и теплоотдачей. Например, в условиях физиологического покоя в организме продуцируется около 1,18 ккал/минуту (или около 70 ккал в час) и такое же количество тепла отдается в окружающую среду. При низкой температуре среды, несмотря на механизм защиты, возрастает потеря тепла организмом. В этих условиях для сохранения температуры тела организм должен эквивалентно повысить теплопродукцию. Таким образом, возникает новый уровень теплового баланса. Например, при температуре воздуха 10°С теплоотдача достигает 120 ккал/час (в условиях комфорта — 70 ккал/час), поэтому для поддержания температуры тела на постоянном уровне теплопродукция тоже должна возрастать до 120 ккал/час.
При высокой температуре окружающей среды, например, при 40°С, отдача тепла значительно уменьшается, например, до 40 ккал/час (вместо 70 ккал/час в условиях комфортной среды). Для поддержания постоянства температуры тела теплопродукция тоже должна снизиться примерно до 40 ккал/час. Устанавливается новый уровень теплового баланса, который и обеспечивает поддержание температуры тела.
Таким образом, ведущим фактором, определяющим уровень теплового баланса, является температура окружающей среды.
Учитывая, что продукция тепла меняется в зависимости от вида физической активности человека, а величина теплоотдачи во многом зависит от температуры окружающей среды, необходимы механизмы регуляции теплопродукции и теплоотдачи. Они осуществляются с участием специализированных структур мозга, объединенных в центр терморегуляции. Принцип регулирования заключается в том, что управляющее устройство (центр терморегуляции) получает информацию от терморецепторов. На основании этой информации оно вырабатывает такие команды, благодаря которым деятельность объектов управления (рабочие структуры, определяющие интенсивность теплопродукции и теплоотдачи) изменяется так, что возникает новый уровень теплового баланса, в результате которого температура тела сохраняется на постоянном уровне. Система терморегуляции может работать в режиме слежения или по принципу рассогласования — изменилась температура крови, изменяется деятельность объектов управления. Однако в системе терморегуляции предусмотрен и более мягкий способ поддержания постоянства температуры тела, который основан на принципе регуляции по возмущению: улавливается изменение температуры среды, и не дожидаясь, когда она отразится на температуре крови, в системе возникают команды, меняющие работу объектов управления таким образом, что температура крови сохраняется постоянной. Кроме того, система терморегуляции может функционировать и в режиме управления по прогнозированию, т. е. досрочного управления (это условные рефлексы): человек еще только собирается выйти на зимнюю улицу, а у него уже возрастает продукция тепла, необходимого для компенсации теплопотерь, которые произойдут у человека на улице в условиях низкой температуры. Во всех случаях для оптимального регулирования интенсивности теплопродукции и теплоотдачи необходима информация о температуре тела (ядра и оболочки). Она передается в ЦНС от терморецепторов.
ФИЗИОЛОГИЯ ТЕРМОРЕЦЕПТОРОВ
Терморецепторы расположены на различных участках кожи, во внутренних органах (в желудке, кишечнике, матке, мочевом пузыре), в дыхательных путях, слизистых, роговице глаза, скелетных мышцах, кровеносных сосудах, в том числе в артериях, аортальной и каротидной зонах, во многих крупных венах, а также в коре больших полушарий, спинном мозге, ретикулярной формации, среднем мозге, гипоталамусе.
Терморецепторы ЦНС — это, скорее всего, нейроны, которые одновременно выполняют роль рецепторов и роль афферентного нейрона.
Наиболее полно изучены терморецепторы кожи. Больше всего терморецепторов на коже головы (лицо) и шеи. В среднем на 1 мм2 поверхности кожи приходится 1 терморецептор. Кожные терморецепторы делятся на холодовые и тепловые. В свою очередь, холодовые подразделяются на собственно холодовые (специфические), реагирующие только на изменение температуры, и тактильно-холодовые, или неспецифические, которые одновременно могут отвечать и на изменение температуры, и на давление.
Холодовые рецепторы располагаются на глубине 0,17 мм от поверхности кожи. Всего их около 250 тысяч. Реагируют на изменение температуры с коротким латентным периодом. При этом частота потенциала действия линейно зависит от температуры в пределах от 41° до 10°С: чем ниже температура, тем выше частота импульсации. Оптимальная чувствительность в диапазоне от 15° до 30°С, а по некоторым данным — до 34°С.
Тепловые рецепторы залегают глубже — на расстоянии 0,3 мм от поверхности кожи. Всего их около 30 тысяч. Реагируют на изменение температуры линейно в диапазоне от 20° до 50°С: чем выше температура, тем выше частота генерации потенциала действия. Оптимум чувствительности в пределах 34—43°С.
Среди холодовых и тепловых рецепторов имеются разные по чувствительности популяции рецепторов: одни реагируют на изменение температуры, равное 0,1 °С (высокочувствительные рецепторы), другие — на изменение температуры, равное 1°С (рецепторы средней чувствительности), третьи — на изменение в 10°С (высокопороговые, или рецепторы низкой чувствительности).
Информация от кожных рецепторов идет в ЦНС по афферентным волокнам группы А-дельта и по волокнам группы С, в ЦНС она доходит с разной скоростью. Вероятнее всего, что импульсы от холодовых рецепторов идут по волокнам А-дельта.
Импульсация от кожных рецепторов поступает в спинной мозг, где расположены вторые нейроны, дающие начало спиноталамическому пути, который заканчивается в вентробазальных ядрах таламуса, откуда часть информации поступает в сенсомоторную зону коры больших полушарий, а часть — в гипоталамические центры терморегуляции.
Высшие отделы ЦНС (кора и лимбическая система) обеспечивают формирование теплоощущения (тепло, холодно, температурный комфорт, температурный дискомфорт). Ощущение комфорта строится на потоке импульсации от терморецепторов оболочки (в основном — кожи). Поэтому организм можно «обмануть» — если в условиях высокой температуры охлаждать тело прохладной водой, как это бывает при летнем купании в зной, то создается ощущение температурного комфорта.
ЦЕНТРЫ ТЕРМОРЕГУЛЯЦИИ
Терморегуляция в основном осуществляется с участием ЦНС, хотя возможны и некоторые процессы терморегуляции без ЦНС. Так, известно, что кровеносные сосуды кожи могут сами по себе реагировать на холод: за счет термочувствительности гладкомышечных клеток к холоду происходит релаксация гладких мышц, поэтому на холоде вначале происходит рефлекторным спазм, что сопровождается болевым ощущением, а потом сосуд расширяется за счет прямого воздействия холода на гладкомышечные клетки. Таким образом, сочетание двух механизмов регуляции дает возможность, с одной стороны, сохранить тепло, а с другой — не позволить тканям испытывать кислородное голодание.
Центры терморегуляции представляют собой в широком смысле совокупность нейронов, участвующих в терморегуляции. Они обнаружены в различных областях ЦНС, в том числе — в коре больших полушарий, лимбической системе (амигдалярный комплекс, гиппокамп), таламусе, гипоталамусе, среднем, продолговатом и спинном мозге. Каждый отдел мозга выполняет свои задачи. В частности, кора, лимбическая система и таламус обеспечивают контроль за деятельностью гипоталамических центров и спинномозговых структур, формируя адекватное поведение человека в различных температурных условиях среды (рабочая поза, одежда, произвольная двигательная активность) и ощущения тепла, холода или комфорта. С помощью коры больших полушарий осуществляется заблаговременная (досрочная) терморегуляция — формируются условные рефлексы. Например, у человека, собирающегося выйти на улицу зимой, заблаговременно возрастает теплопродукция.
В терморегуляции участвуют симпатическая и соматическая нервные системы. Симпатическая система регулирует процессы теплопродукции (гликогенолиз, липолиз), процессы теплоотдачи (потоотделение, теплоотдачу путем теплоизлучения, теплопроведения и конвекции — за счет изменения тонуса кожных сосудов). Соматическая система регулирует тоническое напряжение, произвольную и непроизвольную фазную активность скелетных мышц, т. е. процессы сократительного термогенеза.
Основную роль в терморегуляции играет гипоталамус. В нем различают скопления нейронов, регулирующих теплоотдачу (центр теплоотдачи) и теплопродукцию.
Впервые существование таких центров в гипоталамусе обнаружил К. Бернар. Он производил «тепловой укол» (механически раздражал гипоталамус животного), после чего повышалась температура тела.
Животные с разрушенными ядрами преоптической области гипоталамуса плохо переносят высокие температуры окружающей среды. Раздражение электрическим током этих структур приводит к расширению сосудов кожи, потоотделению, появлению тепловой одышки. Это скопление ядер (главным образом, паравентрикулярных, супраоптических, супрахиазматических) и получило название «центра теплоотдачи».
При разрушении нейронов задних отделов гипоталамуса животное плохо переносит холод. Электростимуляция этой области вызывает повышение температуры тела, мышечную дрожь, увеличение липолиза, гликогенолиза. Полагают, что эти нейроны, в основном, концентрируются в области вентромедиального и дорсомедиального ядер гипоталамуса. Скопление этих ядер получило название «центра теплопродукции».
Разрушение центров терморегуляции превращает гомойотермный организм в пойкилотермный.
Согласно К. П. Иванову (1983, 1984), в центрах теплопродукции и теплоотдачи имеются сенсорные, интегрирующие и эфферентные нейроны. Сенсорные нейроны воспринимают информацию от терморецепторов, расположенных на периферии, а также непосредственно от крови, омывающей нейроны. К. П. Иванов делит сенсорные нейроны на два вида: 1) воспринимающие информацию от периферических терморецепторов и 2) воспринимающие температуру крови. Информация от сенсорных нейронов поступает на интегрирующие нейроны, где происходит суммация всей информации о состоянии температуры ядра и оболочки тела, т. е. эти нейроны «вычисляют» среднюю температуру тела. Затем информация поступает на командные нейроны, в которых происходит сличение текущего значения средней температуры тела с заданным уровнем. Вопрос о нейронах, которые задают этот уровень, остается открытым. Но, вероятно, такие нейроны есть, и они могут быть расположены в коре, лимбической системе или, что более вероятно, в гипоталамусе. Итак, если в результате сличения выявляется отклонение от заданного уровня, то возбуждаются эфферентные нейроны: в центре теплоотдачи — это нейроны, регулирующие потоотделение, тонус кожных сосудов, объем циркулирующей крови, а в центре теплопродукции — это нейроны, которые регулируют процесс образования тепла. Остается пока не ясным, каждый ли центр (теплоотдачи и теплопродукции) занимается «расчетами» и самостоятельно принимает решения, или существует еще какой-то отдельный центр, где совершается этот процесс.
Центры теплоотдачи. При возбуждении эфферентных нейронов центра теплоотдачи может уменьшаться тонус сосудов кожи. Это осуществляется за счет воздействия эфферентных нейронов центра теплоотдачи («сосудов кожи») на сосудодвигательный центр, который, в свою очередь, влияет на активность спинномозговых симпатических нейронов, посылающих поток импульсов к гладким мышцам сосудов кожи. В итоге, при возбуждении гипоталамических нейронов «сосудов кожи» снижается тонус кожных сосудов, возрастает кожный кровоток и увеличивается отдача тепла за счет теплоизлучения, теплопроведения и конвекции. Усиление кожного кровотока способствует также повышению потоотделения (отдачи тепла путем испарения). Если изменение кожного кровотока недостаточно для отдачи тепла, то возбуждаются нейроны, которые приводят к выбросу крови из кровяных депо и, тем самым, — к повышению объема теплопереноса. Если и этот механизм не способствует нормализации температуры, то возбуждаются эфферентные нейроны центра теплоотдачи, которые возбуждают симпатические нейроны, активирующие потовые железы, эти нейроны гипоталамуса можно условно назвать «поторегулирующие нейроны», или нейроны, регулирующие потоотделение. Симпатические нейроны, активирующие потоотделение, располагаются в боковых столбах спинного мозга (Тh>2>—L>2>), а постганглионарные нейроны локализуются в симпатических ганглиях. Постганглионарные волокна, идущие к потовым железам, являются холинергическими, их медиатором является ацетилхолин, который повышает активность потовой железы за счет взаимодействия с ее М-холинорецепторами (блокатор — атропин).
Центры теплопродукции. Эфферентные нейроны центра теплопродукции тоже можно условно разделить на несколько типов, каждый из которых включает в действие соответствующий механизм теплопродукции.
а) Одни нейроны при своем возбуждении активируют симпатическую систему, в результате чего повышается интенсивность процессов, генерирующих энергию (липолиз, гликогенолиз, гликолиз, окислительное фосфорилирование). В частности, симпатические нервы за счет взаимодействия их медиатора (норадреналина) с бета-адренорецепторами активируют процессы гликогенолиза и гликолиза в печени, процессы липолиза в буром жире.
Одновременно, при возбуждении симпатической нервной системы увеличивается секреция гормонов мозгового слоя надпочечников — адреналина и норадреналина, которые повышают продукцию тепла в печени, скелетных мышцах, буром жире, активируя гликогенолиз, гликолиз и липолиз.
б) В гипоталамусе имеются эфферентные нейроны, которые влияют на гипофиз, а через него — на щитовидную железу: возрастает продукция йодосодержащих гормонов (Т>3> и Т>4>), которые, возможно, за счет разобщения процессов окислительного фосфорилирования повышают поток первичной теплоты, т. е. под их влиянием уменьшается аккумуляция энергии в АТФ, а большая часть энергии рассеивается в виде тепла.
в)
В гипоталамическом
центре теплопродукции имеется также
популяция эфферентных нейронов,
возбуждение которых приводит к появлению
терморегуляционного тонуса (при этом
в скелетных мышцах возрастает тонус,
благодаря чему, примерно на 40—60%
возрастает
теплообразование) или возникают
фазноподобные сокращения отдельных
мышечных
волокон, которые получили
название «дрожь». Во всех этих случаях
команда от эфферентных
нейронов гипоталамуса передается, в
конечном итоге, на альфа-мотонейроны.
Центральный дрожательный путь представляет
собой эфферентный путь, идущий
от гипоталамуса к альфа-мотонейронам
через промежуточные образования, в
частности,
через покрышку среднего мозга
(тектоспинальный путь) и через красное
ядро (руброспинальный
тракт). Детали этого пути до сих пор не
ясны.
МЕХАНИЗМЫ ТЕПЛОПРОДУКЦИИ
Источником тепла в организме являются экзотермические реакции окисления белков, жиров, углеводов, а также гидролиза АТФ. При гидролизе питательных веществ часть освобожденной энергии аккумулируется в АТФ, а часть рассеивается в виде теплоты (первичная теплота). При использовании энергии, аккумулированной в АГФ, часть энергии идет на выполнение полезной работы, часть рассеивается в виде тепла (вторичная теплота). Таким образом, два потока теплоты — первичной и вторичной — являются теплопродукцией. При высокой температуре среды или соприкосновении человека с горячим телом, часть тепла организм может получать извне (экзогенное тепло).
При необходимости повысить теплопродукцию (например, в условиях низкой температуры среды), помимо возможности получения тепла извне, в организме существуют механизмы, повышающие продукцию тепла.
Классификация механизмов теплопродукции:
1.Сократительный термогенез — продукция тепла в результате сокращения скелетных мышц:
а) произвольная активность локомоторного аппарата;
б) терморегуляционный тонус;
в) холодовая мышечная дрожь, или непроизвольная ритмическая активность скелетных мышц.
2.Несократительный термогенез, или недрожательный термогенез (продукция тепла в результате активации гликолиза, гликогенолиза и липолиза):
а)в скелетных мышцах (за счет разобщения окислительного фосфорилирования);
б) в печени;
в) в буром жире;
г) за счет специфико-динамического действия пищи.
Сократительный термогенез
При сокращении мышц возрастает гидролиз АТФ, и поэтому возрастает поток вторичной теплоты, идущей на согревание тела. Произвольная мышечная активность, в основном, возникает под влиянием коры больших полушарий. Опыт человека показывает, что в условиях низкой температуры среды необходимо движение. Поэтому реализуются условнорефлекторные акты, возрастает произвольная двигательная активность. Чем она выше, тем выше теплопродукция. Возможно повышение ее в 3—5 раз по сравнению с величиной основного обмена. Обычно при снижении температуры среды и температуры крови первой реакцией является увеличение терморегуляционного тонуса. Впервые его выявили в 1937 г. у животных, а в 1952 г. — у человека. С помощью метода электромиографии показано, что при повышении тонуса мышц, вызванного переохлаждением, повышается электрическая активность мышц. С точки зрения механики сокращения, герморегуляционный тонус представляет собой микровибрацию. В среднем, при его появлении, теплопродукция возрастает на 20—45% от исходного уровня. При более значительном переохлаждении терморегуляционный тонус переходит в мышечную холодовую дрожь. Терморегуляционный тонус экономнее, чем мышечная дрожь. Обычно в его создании участвуют мышцы головы и шеи.
Дрожь, или холодовая мышечная дрожь, представляет собой непроизвольную ритмическую активность поверхностно расположенных мышц, в результате которой теплопродукция возрастает по сравнению с исходным уровнем в 2—3 раза. Обычно вначале возникает дрожь в мышцах головы и шеи, затем — туловища и, наконец, конечностей. Считается, что эффективность теплопродукции при дрожи в 2,5 раза выше, чем при произвольной деятельности.
Сигналы от нейронов гипоталамуса идут через «центральный дрожательный путь» (тектум и красное ядро) к альфа-мотонейронам спинного мозга, откуда сигналы идут к соответствующим мышцам, вызывая их активность. Курареподобные вещества (миорелаксанты) за счет блокады Н-холинорецепторов блокируют развитие терморегуляционного тонуса и холодовой дрожи. Это используется для создания искусственной гипотермии, а также учитывается при проведении оперативных вмешательств, при которых применяются миорелаксанты.
Несократительный термогенез
Он осуществляется путем повышения процессов окисления и снижения эффективности сопряжения окислительного фосфорилирования. Основным местом продукции тепла являются скелетные мышцы, печень, бурый жир. За счет этого вида термогенеза теплопродукция может возрасти в 3 раза.
В скелетных мышцах повышение несократителыюго термогенеза связано с уменьшением эффективности окислительного фосфорилирования за счет разобщения окисления и фосфорилирования, в печени — в основном, путем активации гликогенолиза и последующего окисления глюкозы. Бурый жир повышает теплопродукцию за счет липолиза (под влиянием симпатических воздействий и адреналина). Бурый жир расположен в затылочной области, между лопатками, в средостении по ходу крупных сосудов, в подмышечных впадинах. В условиях покоя около 10% тепла образуется в буром жире. При охлаждении роль бурого жира резко повышается. При холодовой адаптации (у жителей арктических зон) возрастает масса бурого жира и ее вклад в общую теплопродукцию.
Регуляция процессов несократительного термогенеза осуществляется путем активации симпатической системы и продукции гормонов щитовидной железы (они разобщают окислительное фосфорилирование) и мозгового слоя надпочечников.
МЕХАНИЗМЫ ТЕПЛООТДАЧИ
Основная масса тепла образуется во внутренних органах. Поэтому внутренний поток тепла для удаления из организма должен подойти к коже. Перенос тепла от внутренних органов осуществляется за счет теплопроведения (таким способом переносится менее 50% тепла) и конвекции, т. е. тепломассапереноса. Кровь в силу своей высокой теплоемкости является хорошим проводником тепла.
Второй поток тепла — это поток, направленный от кожи в среду. Его называют наружным потоком. Рассматривая механизмы теплоотдачи, обычно имеют ввиду именно этот поток.
Отдача тепла в среду осуществляется с помощью 4 основных механизмов:
1)испарения;
2)теплопроведения;
3)теплоизлучения;
4)конвекции.
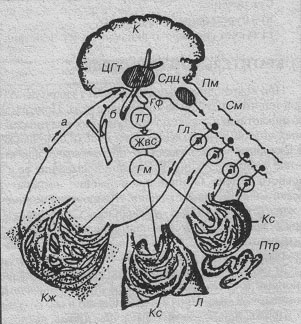
Механизмы теплоотдачи и управление выделением тепла.
К – кора, Кж – кожа, ЦГт – центры гипоталамуса, Сдц – сосудодвигательный центр, Пм – продолговатый мозг, См – спинной мозг, Гф – гипофиз, ТГ – тиреотропный гормон, Жвс – железы внутренней секреции, Гм – гормоны, Птр – пищеварительный тракт, Кс – кровеносные сосуды, Л – легкие, а, б – поток афферентной импульсации.
Вклад каждого механизма в теплоотдачу определяется состоянием среды и скоростью продукции тепла в организме. В условиях температурного комфорта основная масса тепла отдается за счет теплопроведения, теплоизлучения и конвекции и лишь 19—20% — с помощью испарения. При высокой температуре среды до 75—90% тепла отдается за счет испарения.
Теплопроведение — это способ отдачи тепла телу, которое непосредственно контактирует с телом человека. Чем ниже температура этого тела, чем выше температурный градиент, тем выше скорость потери тепла за счет этого механизма. Обычно этот способ отдачи тепла ограничен одеждой и воздушной прослойкой, которые являются хорошими изоляторами тепла, а также подкожным жировым слоем. Чем толще этот слой, тем меньше вероятность передачи тепла к холодному телу.
Теплоизлучение — отдача тепла с участков кожи, не прикрытых одеждой. Происходит путем длинноволнового инфракрасного излучения, поэтому такой вид теплоотдачи еще называют радиационной теплоотдачей. В условиях температурного комфорта за счет этого механизма отдается до 60% тепла. Эффективность теплоизлучения зависит от градиента температуры (чем он выше, тем больше тепла отдается), от площади, с которой происходит излучение, от числа объектов, находящихся в среде, которые поглощают инфракрасные лучи.
Конвекция. Воздух, соприкасающийся с кожей, нагревается и поднимается, его место занимает «холодная» порция воздуха и т. д. Таким способом — за счет тепломассапереноса отдается в условиях температурного комфорта до 15% тепла.
Во всех перечисленных механизмах большую роль играет кожный кровоток: когда его интенсивность возрастает за счет снижения тонуса гладкомышечных клеток артериол и закрытия артериовенозных шунтов — отдача тепла существенно возрастает. Этому также способствует увеличение объема циркулирующей крови: чем больше его значение, тем выше возможность переноса тепла в среду. На холоде происходят противоположные процессы — уменьшается кожный кровоток, в том числе за счет прямого переброса артериальной крови из артерий в вены, минуя капилляры, уменьшается объем циркулирующей крови, меняется и поведенческая реакция: человек или животное инстинктивно занимает позу «калачиком», т. к. в этом случае площадь отдачи тепла уменьшается на 35%, у животных к этому добавляется и реакция — «гусиная кожа» — подъем волос кожи (пилоэрекция), что повышает ячеистость накожного покрова и снижает возможность отдачи тепла.
На долю кистей рук приходится небольшая часть поверхности тела — всего 6%, но их кожей отдается до 60% тепла при помощи механизма сухой теплоотдачи (теплоизлучение, конвекция).
Испарение. Отдача тепла происходит за счет траты энергии (0,58 ккал на 1 мл воды) на испарение воды. Различают два вида испарения, или перспирации: неощущаемую и ощущаемую перспирацию.
а)неощущаемая перспирация — это испарение воды со слизистых дыхательных путей и воды, которая просачивается через эпителий кожного покрова (тканевой жидкости). За сутки через дыхательные пути испаряется в норме до 400 мл воды, т. е. отдается 400x0,58ккал=232ккал/сутки. При необходимости эта величина может быть увеличена за счет так называемой тепловой одышки, которая обусловлена влиянием нейронов центра теплоотдачи на дыхательные нейроны ствола мозга.
В среднем за сутки через эпидермис просачивается около 240 мл воды. Следовательно, за счет этого отдается 240•0,58ккал=139ккал/сутки. Эта величина не зависит от процессов регуляции и различных факторов среды.
Оба вида неощущаемой перспирации за сутки позволяют отдать (400 + 240) • 0,58 = 371 ккал.
б)ощущаемая перспирация (отдача тепла путем испарения пота). В среднем за сутки при комфортной температуре среды выделяется 400—500 мл пота, следовательно, отдается до 300 ккал. Однако при необходимости объем потоотделения может возрасти до 12 л/сутки, т. е. путем потоотделения можно отдать почти 7000 ккал в сутки. За час потовые железы могут продуцировать до 1,5 л, а по некоторым источникам — до 3 л пота.
Эффективность испарения во многом зависит от среды: чем выше температура и ниже влажность воздуха (насыщенность воздуха водяными парами), тем выше эффективность потоотделения как механизма отдачи тепла. При 100% насыщения воздуха парами воды испарение невозможно.
Потовые железы состоят из концевой части, или тела, и потового протока, который открывается наружу потовой порой. По характеру секреции потовые железы делятся на эккриновые (мерокриновые) и апокриновые. Апокриновые железы локализуются, главным образом, в подмышечной впадине, в лобковой области, а также в области половых губ, промежности, околососковом круге молочной железы. Апокриновые железы секретируют жирное вещество, богатое органическими соединениями. Вопрос об их иннервации дискутируется — одни утверждают, что она адренергическая симпатическая, другие считают, что она вообще отсутствует и продукция секрета зависит от гормонов мозгового вещества надпочечников (адреналина и норадреналина).
Видоизмененными апокриновыми железами являются ресничные железы, расположенные в веках у ресниц, а также железы, продуцирующие ушную серу в наружном слуховом проходе, и железы носа (преддверные железы). В испарении, однако, апокриновые железы не участвуют. Эккриновые, или мерокриновые, потовые железы расположены в коже почти всех областей тела. Всего их более 2 млн. (хотя есть люди, у которых они почти полностью отсутствуют). Больше всего потовых желез на ладонях и подошвах (свыше 400 на 1 см2) и в коже лобка (около 300 на 1см2). Скорость потообразования, также как и включение в активность потовых желез, в разных участках тела очень широко варьирует.
По химическому составу пот — это гипотонический раствор: он содержит 0,3% хлористого натрия (в крови — почти 0,9%), мочевину, глюкозу, аминокислоты, аммоний, малые количества молочной кислоты. рН пота варьирует от 4,2 до 7, в среднем рН = 6. Удельный вес — 1,001—1,006. Так как пот — это гипотоническая среда, то при обильном потоотделении больше теряется воды, чем солей, и в крови может происходить повышение осмотического давления. Таким образом, обильное потоотделение чревато изменением водно-солевого обмена.
Потовые железы иннервируются симпатическими холинергическими волокнами — в их окончаниях выделяется ацетилхолин, который взаимодействует с М-холинорецепторами, повышая продукцию пота. Преганглионарные нейроны расположены в боковых столбах спинного мозга на уровне Th>2>—L>2>, а постганглионарные нейроны — в симпатическом стволе.
При необходимости повышения теплоотдачи путем потоиспарения происходит активация нейронов коры, лимбической системы и, главным образом, гипоталамуса. От гипоталамических нейронов сигналы идут к нейронам спинного мозга и постепенно вовлекают различные участки кожи в процесс потоотделения: вначале лицо, лоб, шею, потом — туловище и конечности.
Существуют различные способы активного воздействия на процесс потоотделения. Например, многие жаропонижающие средства, или антипиретики: аспирин и другие салицилаты — повышают потообразование и, тем самым, снижают температуру тела (происходит усиленная теплоотдача путем испарения). Потогонным эффектом обладают также соцветия липы, ягоды малины, листья мать-и-мачехи.
ОБМЕН ВЕЩЕСТВ
Обмен веществ — это процесс метаболизма веществ, поступивших в организм, в результате которого из этих веществ могут образовываться более сложные или, наоборот, более простые вещества.
Человеческий организм, как и организмы других представителей животного и растительного мира, — это открытая термодинамическая система. В нее постоянно поступает поток свободной энергии. Одновременно она отдает окружающей среде энергию, в основном, обесцененную (связанную). Благодаря этим двум потокам энтропия живого организма (степень неупорядоченности, хаоса, деградации) остается на постоянном (минимальном) уровне. Когда же по каким-то причинам поток свободной энергии (негэнтропии) уменьшается (или возрастает образование связанной энергии), то суммарная энтропия организма возрастает, что может привести к его термодинамической смерти.
Согласно термодинамике живых систем, жизнь — это борьба с энтропией, борьба упорядоченности системы с деградацией. Согласно известному уравнению Пригожина, минимальный прирост энтропии имеет место в том случае, если скорость негэнтропийного потока равна скорости энтропийного потока в среду.
Свободная энергия для организма может поступать лишь с пищей. Она аккумулирована в сложных химических связях белков, жиров и углеводов. Для того чтобы освободить эту энергию, питательные вещества вначале подвергаются гидролизу, а потом — окислению в анаэробных или аэробных условиях.
В процессе гидролиза, который осуществляется в желудочно-кишечном тракте, высвобождается незначительная часть свободной энергии (менее 0,5%). Она не может быть использована для нужд биоэнергетики, т. к. не аккумулируется макроэргами типа АТФ. Она превращается лишь в тепловую энергию (первичную теплоту), которая используется организмом для поддерживания температурного гомеостаза.
2-й этап высвобождения энергии — это процесс анаэробного окисления. В частности, таким способом высвобождается около 5% всей свободной энергии из глюкозы при окислении до молочной кислоты. Эта энергия, однако, аккумулируется макроэргом АТФ и используется на совершение полезной работы, например, для мышечного сокращения, для работы натрий-калиевого насоса, но, в конечном итоге, она тоже превращается в теплоту, которая называется вторичной теплотой.
3-й этап — основной этап высвобождения энергии — до 94,5% всей энергии, которая способна высвободиться в условиях организма. Осуществляется этот процесс в цикле Кребса: в нем происходит окисление пировиноградной кислоты (продукт окисления глюкозы) и ацетилкоэнзима А (продукт окисления аминокислот и жирных кислот). В процессе аэробного окисления свободная энергия высвобождается в результате отрыва водорода и переноса его электронов и протонов по цепи дыхательных ферментов на кислород. При этом освобождение энергии идет не одномоментно, а постепенно, поэтому большую часть этой свободной энергии (примерно 52—55%) удается аккумулировать в энергию макроэрга (АТФ). Остальная часть в результате «несовершенства» биологического окисления теряется в виде первичной теплоты. После использования свободной энергии, запасенной в АТФ, для совершения полезной работы она превращается во вторичную теплоту.
Таким образом, вся свободная энергия, которая высвобождается при окислении питательных веществ, в конечном итоге, превращается в тепловую энергию. Поэтому замер количества тепловой энергии, которую выделяет организм, является методом определения энерготрат организма.
В результате окисления глюкоза, аминокислоты и жирные кислоты в организме превращаются в углекислый газ и воду.
Энергетический обмен животного организма (валовый обмен) складывается из основного обмена и рабочей прибавки к основному обмену. Исходной величиной уровня обменных процессов является основной обмен. Указанные стандартные условия определения основного обмена характеризуют те факторы, которые могут влиять на интенсивность процессов обмена веществ у человека. Например, интенсивность обмена веществ подвержена суточным колебаниям, которая возрастает утром и снижается в ночное время. Интенсивность обмена возрастает также при физической и умственной работе. Существенное влияние на уровень обмена оказывает потребление питательных веществ и их дальнейшее переваривание особенно в том случае, если питательные вещества имеют белковую природу. Это явление называют специфически-динамическим действием пищи. Увеличение интенсивности обмена веществ после принятия белковой пищи может продолжаться в течение 12—18 ч. И наконец, если температура окружающей среды становится ниже температуры комфорта, то интенсивность процессов обмена возрастает. Сдвиги в сторону охлаждения приводят к большему усилению обмена веществ, чем соответствующие сдвиги в сторону повышения температуры.
Даже при полном и строгом соблюдении стандартных условий величина основного обмена у здоровых людей может варьировать. Эта вариабельность объясняется различиями в возрасте, поле, росте, массе тела. Как правило, в качестве примерного значения стандартной (основной) интенсивности обмена веществ принимается величина 4,2 кДж/кг • ч [1 ккал/(кг • ч) ]; для человека массой 70 кг соответствующий показатель основного обмена составляет примерно 7100 кДж/сут (1700 ккал/сут).
ПИТАНИЕ
Питание — это процесс усвоения организмом веществ, необходимых для построения и обновления тканей его тела, а также для покрытия энергетических затрат.
В целом эволюция пищевых потребностей животных организмов включала в себя процесс ограничения собственного синтеза ряда соединений с одновременным расширением потребления органических соединений определенных типов. Это привело к выделению целой группы веществ, незаменимых для высших животных и человека, т. е. необходимых для обмена веществ, но не синтезируемых самостоятельно.
Использование пищевых средств, состоящих в основном из сложных соединений растительного или животного происхождения, для энергетических или пластических нужд организма возможно только после гидролиза этих средств и превращения в сравнительно простые соединения, лишенные видовой специфичности. Пищевые потребности разных видов животных различны в зависимости от того, какие пищевые вещества организм способен синтезировать и какие должны поступать извне. И все же в основном различия в пищевых потребностях обусловлены способами переваривания (гидролиза) пищи. Это связано с тем, что у высших животных организмов промежуточные процессы обмена веществ протекают сходным образом.
В обмене веществ (метаболизм) и энергии различают два процесса: анаболизм и катаболизм. Под анаболизмом понимают совокупность процессов, направленных на построение структур организма главным образом через синтез сложных органических веществ; под катаболизмом — совокупность процессов распада сложных органических соединений и использование образовавшихся при этом сравнительно простых веществ в процессах энергообмена. В основе анаболизма и катаболизма лежат соответственно процессы ассимиляции и диссимиляции, которые в организме взаимосвязаны и в нормальном организме сбалансированы.
В целом потребности животных достаточно однородны: они нуждаются в сходных по структуре питательных веществах для энергообмена; в веществах типа аминокислот, пуринов и некоторые липидов для построения сложных белковых молекул и клеточные структур; в специальных катализаторах обмена веществ и стабилизаторах клеточных мембран; в неорганических ионах и соединениях для физико-химических процессов в организме и, наконец, в универсальном биологическом растворителе — воде — для создания среды клеточного обмена веществ.
В конечном итоге в состав пищи высокоорганизованных организмов входят органические вещества, подавляющая часть которых относится к белкам, липидам и углеводам. Продукты их гидролиза — аминокислоты, жирные кислоты, глицерин и моносахара — тратятся на энергообеспечение организма. В процессах энергообмена аминокислоты, жирные кислоты и моносахара взаимосвязаны общими путями их превращения. Поэтому как энергоносители пищевые вещества могут взаимозаменяться в соответствии с энергетической ценностью (правило изодинамии).
Энергетическую (калорическую) ценность пищи оценивают по количеству тепловой энергии, высвобождающейся при сгорании 1 г пищевого вещества (физиологическая теплота сгорания), которую выражают традиционно в килокалориях или по СИ — в джоулях (1 ккал = 4,187 кДж). Расчеты показали, что энергетическая ценность жиров (38,9 кДж/г; 9,3 ккал/г) в два раза выше, чем белков и углеводов (17,2 кДж/г; 4,1 ккал/г). Белки и углеводы имеют одинаковую энергетическую ценность и могут заменяться 1:1 в весовом соотношении.
Для поддержания стационарного состояния организма общие затраты энергии должны покрываться поступлением пищевых веществ, несущих в своих химических связях эквивалентный запас энергии. Если количества поступающей пищи для покрытия энерготрат недостаточно, то энерготраты компенсируются за счет внутренних резервов, главным образом — жира. Если же масса поступающей пищи по энергоносителям превышает расход энергии, то идет процесс запасания жира независимо от состава пищи.
Однако следует всегда помнить, что эти три источника энергии являются и пластическим материалом животного организма. Поэтому длительное исключение одного из трех питательных веществ из пищевого рациона и замена энергетически эквивалентным количеством другого вещества недопустимы.
Под рациональным питанием понимают питание, достаточное в количественном и полноценное в качественном отношении. Основа рационального питания — сбалансированность, т. е. оптимальное соотношение потребляемой пищи. Сбалансированное питание должно включать белки, жиры и углеводы в массовой пропорции, примерно 1:1:4. В качественном отношении пища должна быть полноценной, т. е. содержать белки (включающие незаменимые аминокислоты), незаменимые жирные кислоты (так называемый витамин F), витамины, в большинстве входящие в состав катализирующих систем, и большую группу витаминоподобных веществ, неорганических элементов и воду.
ЗАКЛЮЧЕНИЕ
Жизнь сопряжена с непрерывным расходом энергии, которая необходима для функционирования организма. С точки зрения термодинамики, живые организмы относятся к открытым системам, так как для своего существования они непрерывно обмениваются с внешней средой веществами и энергией. Источником энергии живых организмов служат химические превращения органических веществ, поступающих из окружающей среды. Превращение этих веществ из сложных в простые и приводит к высвобождению энергии, заключенной в химических связях. Извлечение энергии из химических связей осуществляется главным образом с затратой молекулярного кислорода (аэробный обмен); окислению в ряде цепей предшествует бескислородное расщепление (анаэробный обмен).
Основным аккумулятором энергии для использования ее в клеточных процессах является аденозинтрифосфат (АТФ). С помощью энергии АТФ обеспечивается возможность синтеза белка, деления клеток, поддержания их осмотического градиента, мышечного сокращения и др. Согласно первому закону термодинамики, химическая энергия АТФ, пройдя через промежуточные стадии, в конечном итоге превращается в тепловую, которая и теряется организмом. Поэтому интенсивность энергообмена организма является суммой энерготрат на функцию клеточных систем, аккумулированной энергии и потерь ее в виде теплоты.
Жизнь организма зависит от протекания химических реакций с превращением всех видов энергии в тепловую. Скорость химических реакций, а следовательно, и энергообмена зависит от температуры тканей. Теплота как конечное превращение энергии способна переходить из области более высокой температуры в область более низкой. Температура тканей определяется соотношением скорости метаболической теплопродукции их клеточных структур и скорости рассеивания образующейся теплоты в окружающую среду. Следовательно, теплообмен между организмом и внешней средой является неотъемлемым условием существования животных организмов. Для поддержания нормальной (оптимальной) температуры тела у животных организмов имеется система регуляции теплообмена со средой.
Животные организмы подразделяются на пойкилотермные и гомойотермные. Пойкилотермные (стоящие на более низких ступенях филогенетической лестницы) обладают несовершенными, но все же достаточно эффективными механизмами терморегуляции. Эти механизмы включают химическую систему температурной компенсации, позволяющую удерживать устойчивый энергообмен при значительных перепадах температуры тела, терморегуляцию поведением (выбор оптимальной температуры среды) и температурный гистерезис (способность захватывать теплоту из внешней среды быстрее, чем ее терять).
Гомойотермия — более позднее приобретение эволюции животного мира. К истинно гомойотермным животным относят птиц и млекопитающих, так как эти животные способны поддерживать постоянную в пределах 2°С температуру тела мри сравнительно широких колебаниях температуры внешней среды.
В основе гомойотермии лежит более высокий, чем у пойкилотермных животных, уровень энергообмена за счет усиления роли тиреоидных гормонов, стимулирующих работу клеточного натриевого насоса. Высокий энергообмен привел к формированию совершенных механизмов регуляции тепловой энергии в организме.
Ряд животных относится к группе гетеротермных организмов: при одних условиях они пойкилотермные организмы, при других – гомойотермные.
Для поддержания постоянной температуры тела гомойотермные животные обладают химической и физической терморегуляцией. Физическая терморегуляция осуществляется изменением теплопроводности покровных тканей тела (изменение кровотока кожи, пилоэрекция, испарение влаги с поверхности тела или ротовой полости).
Химическая терморегуляция осуществляется путем увеличения теплообразования в организме. Выделяют два основных источника химической терморегуляции (регулируемого теплообразования): сократительный термогенез за счет произвольной активности локомоторного аппарата, терморегуляционного тонуса и дрожи мышц и несократительный термогенез за счет бурой жировой ткани, специфико-динамического действия пищи и др.
Управление теплообменом осуществляется активностью тepморецепторов, информация от которых поступает в центр терморегуляции гипоталамуса, управляющий реакциями химической и физической терморегуляции.
Длительное пребывание в условиях высокой или низкой температуры окружающей среды приводит к существенным изменениям свойств организма, повышающих его устойчивость к действию соответствующих температурных факторов.
Построение и обновление тканей тела, а также покрытие энерготрат организма должны обеспечиваться адекватным питанием. В обмене веществ и энергии различают два процесса: анаболизм и катаболизм. Под анаболизмом понимают совокупность процессов, направленных на построение структур организма главным образом через синтез сложных органических веществ. Катаболизм — это совокупность процессов распада сложных органических веществ с целью высвобождения энергии. В основе анаболизма и катаболизма лежат соответственно процессы ассимиляции и диссимиляции, которые взаимосвязаны и сбалансированы.
Пищевые потребности животных достаточно однородны: необходимые вещества для энергообмена (белки, жиры, углеводы), вещества для построения сложных белковых молекул и клеточных структур (аминокислоты, пурины, липиды, углеводы), специальные катализаторы обмена (витамины) и стабилизаторы клеточных мембран (антиоксиданты), неорганические ионы и универсальный биологический растворитель — вода.
Энергетическую ценность пищи определяют по количеству тепловой энергии, высвобождаемой при сгорании 1г пищевого вещества (физиологическая теплота сгорания).
Под рациональным питанием понимают питание, достаточное в количественном и полноценное в качественном отношении. Основа рационального питания — сбалансированность, т. е. оптимальное соотношение потребляемой пищи. Сбалансированное питание должно включать белки, жиры и углеводы в массовой пропорции, примерно 1:1:4. В качественном отношении пища должна быть полноценной, т. е. содержать белки (включающие незаменимые аминокислоты), незаменимые жирные кислоты (так называемый витамин F), витамины, в большинстве входящие в состав катализирующих систем, и большую группу витаминоподобных веществ, неорганических элементов и воду.
СПИСОК ИСПОЛЬЗУЕМОЙ ЛИТЕРАТУРЫ
1) Мак-Мюрей В. Обмен веществ у человека. М., 1980.
2) Нортон А., Эдхолм О. Человек в условиях холода. М., 1957.
3) Общий курс физиологии человека и животных / под ред А. Д. Ноздрачева. М., 1991. кн. 2.
4) Основы физиологии / под ред. П. Стерки. М., 1984.
5) Слоним А. Д. Эволюция терморегуляции. Л., 1986.
6) Физиология терморегуляции: Руководство по физиологии / под ред. К. П. Иванова. Л., 1984.
7) Физиология человека / под ред. Н.А.Агаджаняна, В.И.Циркина. СПб., 1998.
8) Физиология человека / под ред. Р. Шмидта, Г. Тевса. М., 1986. Т. 4.