Біохімія трансгенної картоплі в умовах України
МІНІСТЕРСТВО АГРАРНОЇ ПОЛІТИКИ УКРАЇНИ
СУМСЬКИЙ ДЕРЖАВНИЙ АГРАРНИЙ УНІВЕРСИТЕТ
АГРОНОМІЧНИЙ ФАКУЛЬТЕТ
Кафедра захисту рослин
Завідувач кафедрою_____________________
А.К.Мішньов
ДИПЛОМНА РОБОТА
на тему: «Біохімія трансгенної картоплі в умовах
України»
Для одержання кваліфікації спеціаліста
за спеціальністю 7.130105 – Захист рослин»
Виконавець _________________ (Загорулько О.)
Науковий керівник ______________ (Чіванов В.Д.)
Консультант з екологічної
експертизи ________________ (Жатова Г.О.)
Консультант з питань
техніки безпеки _________________ (Шандиба О.Б.)
Суми - 2001
ЗМІСТ
Стор.
Вступ..................................................................................................................
1. Огляд літератури…………………………………………………………….
Загальна характеристика фізико-хімічних властивостей глікоалкалоїдів
рослин родини Solanaceae................................………………………………
Глікоалкалоїди рослин картоплі та їх будова………………………….
Глікоалкалоїди як токсичні сполуки……………………………………
Роль глікоалкалоїдів в явищі імунітета до бактеріальних, грибкових захворювань і шкідників та їх значення в селекції нових сортів
картоплі……………………………………………………………………
2. Природні умови господарства......................................................................
Методика проведення досліджень..............................................................
Результати досліджень.................................................................................
Охорона праці...............................................................................................
Охорона навколишнього середовища.......................................................
Висновки..........................................................................................................
Список використаної літератури...............................................................
ВСТУП
Підвищений “тиск” на організм людини ксенобіотичних сполук природного та штучного походження обумовлює необхідність поглибленого вивчення таких сполук, і в першу тих із них, котрі входять до складу найбільш поширених харчових продуктів, зокрема картоплі. Глікоалкалоїди картоплі являють собою групу вторинних метаболітів, яким притаманні найрізномантніші види біологічної активності [ ]. Так, дослідами in vitro доведена здатність цих сполук спричинювати хромосомні аберації, порушувати цілісність ліпосом та природніх біомембран тощо [ ]. Останнє свідчить на користь того, що глікоалкалоїдам притаманна потенційна мутагенна і, можливо, канцерогенна активність [ ]. Виходячи з наведеного, цілком природньою є підвищена зацікавленість в дослідженні глікоалкалоїдів з боку фахівців в галузях медичної токсикології і генетики людини, тим більше, що в останні роки на світовий ринок виходять нові сорти продовольчої картоплі, отриманої як шляхом “класичної” селекції за участі дикоростучих видів роду Solanum [ ], так і за допомогою генетичної інженерії (New Leaf, Monsanto, США). Обидва підходи не виключають біосинтезу в тканинах рослин картоплі, зокрема бульбах, окрім характерних для культурної картоплі глікоалкалоїдів -Соланіну та -Чаконіну “мінорних” глікоалкалоїдів (солакаулін, соласолін, соламаргін, деміссін, томатін, лептін І і ІІ, лептінін-І і ІІ тощо), які ведуть походження з дикоростучих попередників, або синтезуються de novo [ ]. Останнім притаманні токсикологічні характеристики, відмінні від добре відомих аналогічних показників -Соланіну та -Чаконіну. Якщо прийняти до уваги встановлений факт щодо синергетичного багаторазового підсилення біологічної активності суміші двох різних за хімічною структурою глікоалкалоїдів [ ], то стає цілком обгрунтованою необхідність досліджень кількісних та якісних показників глікоалкалоїдів продовольчої картоплі паралельно з селекцією та молекулярною інженерією, успішність яких залежить від наявності нових високочутливих методів фізико-хімічного аналізу. Складність аналізу сумарної фракції глікоалкалоїдів з рослин картоплі пов’язана, зокрема з тим, що субфракція “мінорних” глікоалкалоїдів не перевищує за масою 0,5-5% [ ].
Останнім часом в аналізі біомолекул набули широкого розповсюдження методи м’якоіонізаційної мас-спектрометрії, зокрема плазмено-десорбційна мас-спектрометрія (PDMS). Зважаючи на це, нами зроблена спроба проаналізувати фракцію глікоалкалоїдів бульб та квітів картоплі трансгенних сортів NewLeaf 6 Russet Burbank та NewLeaf 6 Atlantic за допомогою PDMS мас-спектрометрії. Таким чином, метою наших досліджень був докладний аналіз глікоалкалоїдів бульб та квітів картоплі трансгенних сортів NewLeaf 6 Russet Burbank та NewLeaf 6 Atlantic новітнім мас-спектрометричним методам.
ОГЛЯД ЛІТЕРАТУРИ
Загальна характеристика фізико-хімічних властивостей глікоалкалоїдів рослин родини Solanaceaе
Рослини картоплi культурних та диких видів мiстять серед численних органiчних сполук групи вторинних метаболітів глiкоалкалоїди [ ], найважливiшими з яких є a-соланiн, a-чаконiн, деміссин, лептіни I,II,III, соламарін, коммерсонін [ ]. Фiзiологiчнi функцii глiкоалкалоїдiв пов'язанi з явищем стiйкостi рослин картоплі проти фiтопатогенiв i шкiдникiв [ ]. Крiм того, глiкоалкалоїди беруть безпосередню участь в бiохiмiчних процесах, які притаманні тканинам листкiв, пагонiв, квiток, бульб рослин картоплi (цвiтiння, видова сумiснiсть, утворення насiння); обумовлюють разом з iншими вторинними метаболiтами смаковi якостi та придатнiсть картоплi для переробки [ ], а також мають певне значення як токсичні, ембріотропні і канцерогеннi сполуки природнього походження [ ]. Зважаючи на це, багато перспективних за врожайнiстю та iншими господарсько-корисними якостями сортiв картоплi не були рекомендованi для широкого використання саме завдяки пiдвищеному вмiсту глiкоалкалоїдiв [ ]. На думку вiтчизняних та зарубiжних [ ] дослiдникiв, кiлькiснi та якiснi характеристики глiкоалкалоїдiв нових сортiв i гiбридiв картоплi заслуговують на поглиблене вивчення. Одним з найважливіших питань, пов’язаних з біохімією та фізіологією глікоалкалоїдів, яке і досі не знайшло однозначного вирішення, є встановлення корелятивних зв’язків між параметрами метаболізму глікоалкалоїдів та ступенем стійкості сортів картоплі проти шкідників і фітопатогенів [ ]. Іншим дискусійним питанням є невизначеність зв’язків між кількісним накопиченням глікоалкалоїдів в тканинах рослин картоплі різних сортів, зокрема бульбах і умовами вирощування (погодно-кліматичні умови протягом конкретного вегетаційного сезону, тип грунтів, удобрення тощо), а також умовами зберігання (температура, освітленість, вологість тощо)[ ]. На думку провідних вчених дискусійні та суперечливі результати в цьому напрямку дослідження отримані тому, що, по-перше, на інтенсивність біосинтезу вторинних метаболітів взагалі і глікоалкалоїдів зокрема, різноспрямовано впливає велика кількість факторів зовнішнього середовища і це, разом з підвищеною лабільністю та чутливістю ферментних систем метаболізму глікоалкалоїдів до зазначених факторів обумовлює суттєві розбіжності, про які згадувалось вище; і, по-друге, більшість ранніх робіт були проведені без урахування різноманітності глікоалкалоїдів за хімічною структурою – в межах цих досліджень визначали тільки їх сумарну кількість ваговим або колориметричним методами [ ].
Проте, глікоалкалоїди картоплі являють собою досить велику групу сполук, до складу якої входять «мажорні» (a-соланiн та a-чаконiн у культурних видів картоплі) і численні «мінорні» (b-соламарін, деміссидін, 5b-соланідан-3a-ол, томатіденол) компоненти. Крім того, тканини рослин картоплі містять спеціалізовані ферментні системи, які модифікують інтактні молекули глікоалкалоїдів з утворенням численних проміжних сполук, фізіологічна роль яких вичерпно не з’ясована (b-соланін, b-чаконін, g-соланін, g-чаконін тощо)[ ]. Таким чином, дослідження статичних і динамічних кількісних та якісних параметрів глікоалкалоїдів рослин картоплі різних сортів в процесах вирощування в умовах конкретних кліматичних зон, зберігання та переробки, а також під впливом фітопатогенів, мають певні перспективи за умов використання з метою аналізу цих сполук нових високоінформативних фізико-хімічних методів досліджень [ ]. В Сумському ДАУ та Інституті прикладної фізики НАНУ (м.Суми) в 1994-2000 рр. в межах проекту «М’якоіонізаційна мас-спектрометрія в агробіології» провели цикл досліджень глiкоалкалоїдів рослин картоплi за допомогою новiтнього фiзико-хiмiчного методу аналізу полярних, термолабільних, нелетючих органiчних сполук – плазмено-десорбційної часопрольотної плазмено-десорбцiйної мас-спектрометрiї з iонiзацiєю уламками подiлу 252Cf (МС-ПДМС)[ ].
Алкалоїди являють собой велику групу рослинних речовин різного хімічного складу, яким притаманна специфічна дія на організм ссавців та людини. Алкалоїди є азотовміськими гетероциклічними сполуками, що містять в своєму складі атом азоту. Загальною властивостю даної групи речовин є їх надзвичайно сильна фізиологічна дія на організм тварин та людей. Більшість алкалоїдів діють на нервову систему: в невеликих дозах здійснюють збуджуючий вплив, в великих - пригнічують, а в ще більших є сильнодіючими отрутами [ ]. Фізіологічне значення алкалоїдів в життєдіяльності самих рослин досі не з`ясовано. До цього часу не вдалося ще з великою достовірностю показати місце алкалоїдів в життєво важливих процесах обміну речовин, з іншого боку немає неспростовних доказів того, що алкалоїдоносні види рослин можуть існувати без алкалоїдів [ ].
В ряді робіт відзначається вплив алкалоїдів на обмін речовин у рослин, виникнення квітів, формування органел та органів рослин, ріст клітин, поділ ядер і т.д. Було доведено, що алкалоїди в клітинах рослин якимось чином ізольовані від внутрішнього середовища клітини. Так, розчини нікотину, нанесені на рослинні тканини табаку, чинять всебічну негативну фітотоксичну дію навіть при концентрації, рівній їх концентрації в тканинах. Доведено, що високий вміст алкалоїдів часто пов`язаний зі зниженням сили росту рослин. Інакше кажучи, у рослин в період вегетації та спокою існує тісний взаємозв`язок: «білковий азот - алкалоїди». Цими причинами, на думку Горяєва, пояснюється зміна складу алкалоїдів, а в деяких випадках і їх повна відсутність в тканинах алкалоїдних рослин (Solsola richteri. Girgensonia Oppositiflora. Anabasis turkestanika) [ ].
1.1. 1. Глікоалкалоїди рослин картоплі та їх будова.
Глікоалкалоїди, які містяться в бульбах картоплі, раніше називали загальним терміном «соланін». В наш час встановлено, що так званий «соланін» являє собою суміш якнайменше 6 різних глікозідних сполук: a-, b-, g-соланіна та a-, b-, g-чаконіна. Ці сполуки мають один і той же аглікон («корове ядро») – соланідин і відрізняються між собою складом глікозидного ланцюжка (Рис.1). Трисахаріди - a-соланін і a-чаконін є основними формами і складають в тканинах картоплі біля 95% усієї суми глікоалкалоїдів групи соланіна [ ]. Крім a-, b-, g-соланіна та a-, b-, g-чаконіна, відомі інші глікоалкалоїди пасльонових: солакаулін, соласолін, соламаргін, демісін, томатін, лептін, лептінін-1, лептінін-2 та інші. Всі гликоалкалоїди складаються з двох основних складових, виникаючих при кислотному гідролізі цих речовин – аглікона, який і є власне «алкалоїдом», і декількох моносахарів, які знаходяться в молекулі глікоалкалоїда в вигляді глікозідного ланцюга, зв`язаного з агліконом [ ]. Глікоалкалоїди культурної та дікої картоплі (Solanum) можна систематизувати в окремі групи. До групи соланідіна відносяться: a-, b-, g-соланін та a-, b-, g-чаконін, солакаулін і глікоалкалоїд, виділений з групи Akaulia. Іншу групу алкалоїдів складають сполуки, агліконом яких є соласодін. Сюди входять соласонін і соламаргін. Соласонін і соламаргін є основними алкалоїдами Solanum nigrum, Solanum laciniatum більшості інших видів Solanum
[ ].
Соласодамін знайдений в листках Solanum auriculatum і Solanum magrinatum. В групу, яка містить в якості аглікона демісідін, входить демісин, знайдений у Solanum demissum і глікоалкалоїд, виділений із Solanum polyadenium. Лептіни – глікоалкалоїди,
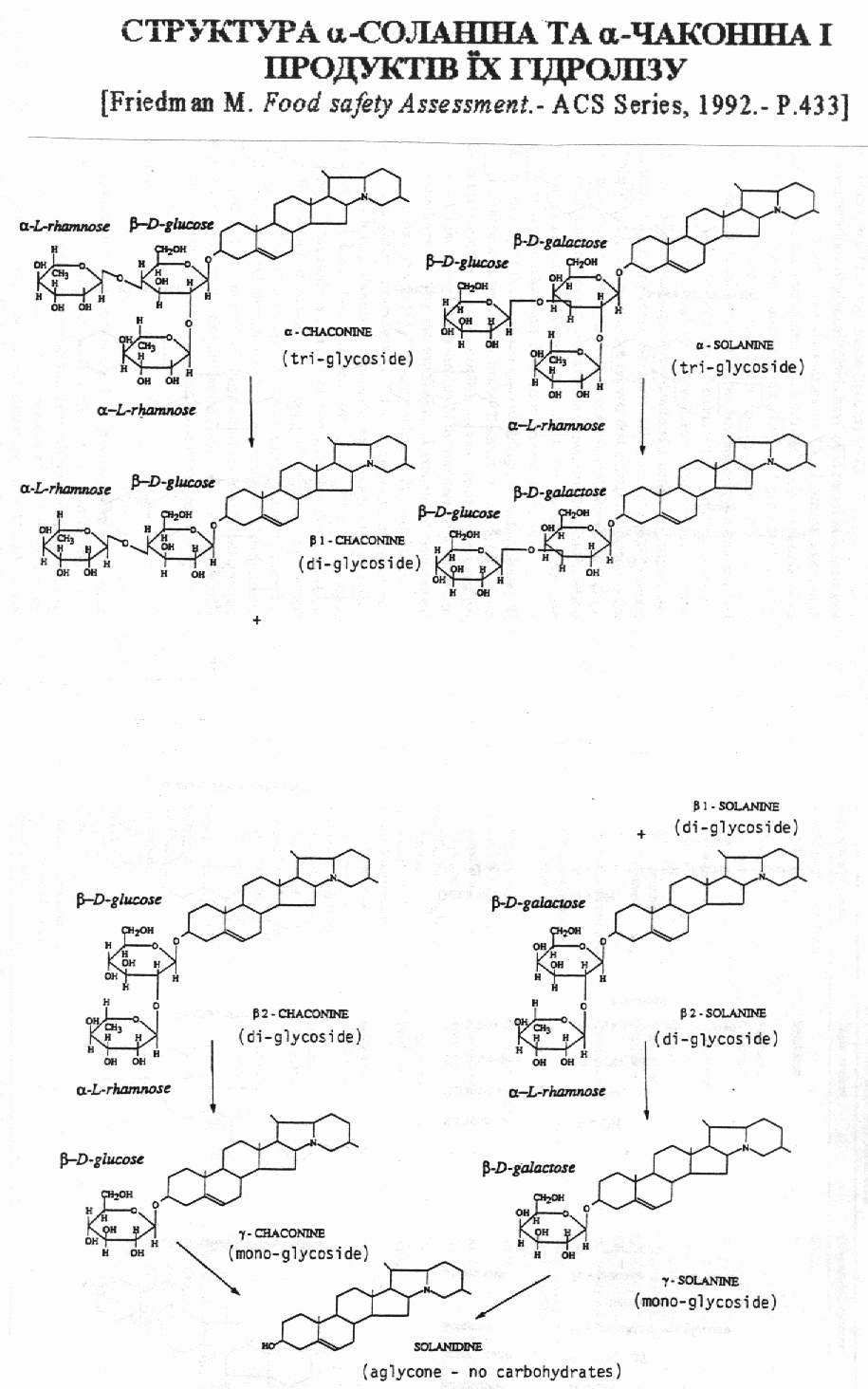
Рис.1. Хімічна структура агліконів, знайдених в рослинах картоплі [ ].
знайдені Куном і Левом, виявлені у Solanum chacoense. Групи Eutuberosa, Andigena, і Commersonia містять тільки - соланін, ці види містять також чаконін. Ці два глікоалкалоїди зустрічаються завжди разом, що доведено хроматографічними дослідженнями. Демісин присутній в рослинах групи Demissa і Acaulia, за виключенням Solanum acaulia, в складі якого знайдено солакаулін. Демісин знайдений також у Solanum jamesii. У форм, які не дають бульб, крім -соланіна, який і тут частіше всього зустрічається, найбільшу групу складають форми, які містять томатін. І тільки єдина форма Solanum poliadenium, яка створює бульби, містить томатін. Інші алкалоїди зустрічаються тільки в деяких форм, нприклад, соласодамін – у Solanum sodomaneum i Slanum aviculare.
Всі алкалоїди картоплі належать до стероїдної групи похідних циклопентанопергідрофенантрена і складаються з алкалоїда (аглікона) та цукру, зв’язаного з ним глікозідним зв’язком. Аглікон має стероїдну будову, містить атоми азоту, який утворює третинну або вторинну аміногрупу. Прикладом першого типу являється соланідін, другого-томатідін і соласодін. Глікозідна частина являє собою моно-, ді-, три-, і тетрасахаріди; до їх складу може входити глюкоза (Glu), галактоза (Gal), рамноза (Rha) і ксілоза (Xyl). Наприклад, -соланін складається з аглікона-соланідіна і трисахаріда, який при гідролізі кислотою розпадається на галактозу, глюкозу і рамнозу. Соланін має розгалужений сахарний ланцюжок: безпосередньо з агліконом зв`язана галактоза, а глюкоза і рамноза складають бокові ланцюги. Подібну будову мають глікозідні частини глікоалкалоїдів демісіну, томатіну, -чаконіну, соласодіну і соламаргіну [ ].
Під впливом гідролізу глікоалкалоїди розпадаються на цукор і аглікон, фізичні і хімічні властивості якого і вказують на велику схожість його зі стероїдами. Стероїди утворюють різноманітну групу важливих природних речовин. Сюди відносяться стеріни, жовчні кислоти, гормони розмноження, вітаміни групи Д, глікозиди, різні алкалоїди і інші речовини. Базовою структурою усіх стероїдів є стеран (циклопентанопергідрофенантрен).
До класу стероїдів належать також фітостерини: сіто-, стігма-, і кампестерін, холестерін, ергостерін, жовчні кислоти.
Сполуки стероїдів з сахаром належать до групи глікозідів. В цю групу, крім глікоалкалоідів, входять «сердечні глікозіди», які містяться в деяких рослинах із родини Strophanthus і Digitalis. Стероїдні саконіни також являють собою глікозіди агліконів, котрі э похідними циклопентанопергідрофенантрена. Сапоніни не містять в своєму складі атом азоту [ ].
Одним з доказів схожості аглікона глікоалкалоідів зі стероїдами є те, що їх можливо перетворити в метилциклофенантрен. Ця сполука синтезується тим же шляхом із стероїдів. При третьому атомі вуглецю аглікона знаходиться група, яка може сполучатися глікозідно з моносахаридами. Остаточно стероїдну будову аглікона вдалося довести, коли похідне аглікона отримали з відомих стероїдів і, навпаки, похідні стероїдів- із аглікона глікоалкалоїдів; наприклад, із безазотної речовини криптогеніна - соласодін, із соласодіна - стероїдний сапогенін - діосгенін. Веліка схожість з сапонінами виявляється і в будові сахарної частини глікоалкалоідів. Загальною властивістю сапонинів і глікоалкалоідів являється їх гемолітична дія [ ].
Таким чином, основними глікоалкалоїдами культурної картоплі є - соланін та -чаконін. Всі глікоалкалоїди різних видів картоплі складаються з двох основних частин: аглікона (алкалоїда), який має стероїдну будову, і молекули сахару, в склад якої входять моно-, ді-, три-, або тетрасахариди.
1.1.2. Глікоалкалоїди як токсичні сполуки
Глікоалкалоїди соланін та чаконін в невеликій кількості розглядається як цілком можливі компоненти бульб картоплі, при надмірній кількості вони погіршують смакові якості картоплі. У бульб з`являється гіркуватий смак, при вживанні таких бульб в харчування віникають чуття зуду в горлянці [ ]. Встановлена визначена кореляція між наявністю глікоалкалоїдів і органолептичними смаковими якостями вареної картоплі. При дуже низькому вмісті глікоалкалоїдів в бульбах (порядку 1-5 мг%) смак вареної картоплі трохи гірший, ніж при середньому (5-10 мг%). Але збільшення їх вмісту вище за 26 мг% на сиру речовину дуже погіршує смак вареної картоплі і стає небезпечним для здоров`я людини [ ].
Бульби картоплі, які містять ніж 20 мг% глікоалкалоїдів, не рекомендується вживати в харчування, осбливо коли картопля вареться зі шкірочкою. Почищені бульби менш небезпечні, так як значна частина глікоалкалоїдів видаляється зі шкірочкою, а частина переходить в воду, при варинні ж неочищених бульб цього не відбувається вз`язку з слабкою проникністю шкірочки. За даними М.Г.Шевченко [ ], нйбільшу небезпеку для здоров`я становить нерозчинний в воді соланін, зосереджений в основі пророслх вічок.
Роль глікоалкалоїдів в явищі імунітета до бактеріальних, грибкових захворювань і шкідників та їх значення в селекції нових сортів картоплі.
Глікоалкалоїди привертають увагу селекціонерів-картоплярів в зв`язку з їх токсичною дією на бактеріальні та грибкові захворювання картоплі, а також в зв`язку з токсичною дією на колорадського жука [ ].
Встановлено, що соланін затримує ріст бактерій, а 0,1-0,5% -ний розчин його викликає відмирання протоплазми. Виділені фунгіцидні якості глікоалкалоїдів в відношенні ряду патогенних грибів, таких як Microsporum audoine, Trichophyton gypseum, Fusarium oxуsporum, зооспор Phytophthora infestans, Helminthosporum carbonum.
В Інституті біохімії РАН досліджували фунгіцидні властивості глікоалкалоїдів і інших сполук картоплі, наділених антибіотичними якостями, а також вивчали дію механічних пошкоджень і зараження Phytophtora infestans на утворення і розподіл глікоалкалоїдів в тканинах бульб [ ]. Виявилось, що глікоалкалоїдам -соланіну та -чаконіну притаманна більша фунгіцидна активність ніж фенолам (кофейній і хлорогеновій кислотам). Так, в концентрації 0,1 мг/мл останні не впливали на кінетичні показники росту фітофтори, тоді як глікоалкалоїди пригнічували його майже на 70%. Зооспори цього паразита гинули вже при концентрації -соланіна 5 мкг/мл.
Соланин, чаконін і соланідін інгібують радіальний рост Alternaria solani на картоплі. По ступеню інгібування на першому місці стоїть соладінін, потім чаконін і соланін. При концентрації 500 мг/л соланін викликає 50% інгібування росту після 96-годинного періоду інкубації при 24оС. Встановлено наявність збільшення чутливості листків до розвітку пошкодження яке супроводжується зменшенням концентрації глікоалкалоїдів в листках в міру їх старіння. Наприклад, 30-денні листки сорту Катадин містять 950 мг/кг глікоалкалоїдів і стійкі до паразиту, тоді як старіючі 90-денні листки містять тільки 150мг/кг глікоалкалоїдів і значно уражаються грибом [ ].
Проте подальші дослідження показали, що хоча глікоалкалоїди і наділені фунгітоксичністю, вони все ж таки не є вирішальним фактором в явищі фітоімунітета картоплі [ ]. Взагалі після інфікування в бульбах картоплі накопичується хлогенова та кофейна кислоти, скополетин, -соланін, -чаконін, рішитин, фітуберин. Всі зазначені сполуки, за виключенням рішитіна та фітуберіна, присутні в шкірочці бульби в кількостях рівних або більших, аніж синтезованих за умов інфікування патогеном очищеної картоплі. Хлорогенова та кофейна кислоти знайдені в усіх частинах рослини. Вони продукуються в бульбах картоплі в результаті пошкоджень та інфекції багатьма патогенними та не патогенними організмами і накопичуються (або присутні) в деяких здорових тканинах рослини картоплі в кількостях, які можуть пригнічувати мікроорганізми. Механічне пошкодження чи інфекція викликає швидке окислення цих речовин, а продукти окислення токсичні для багатьох мікроорганізмів. На ранніх стадіях інфекції після інокуляції бульб деякими расами фітофтори накопичуються поліфеноли. В тканинах, інокульованих несумісними расами фітофтори, або в зрізах накопичується тільки хлорогенова кислота.
Глікоалкалоїди -соланін і -чаконін зосереджуються біля пошкоджених ділянок бульби [ ]. Про те, що глікоалкалоїди не є вирішальним фактором в імунітеті картоплі, свідчать результати дослідів, проведених на раневій перидермі бульб. В зараженій фітофторою раневій тканині картоплі глікоалкалоїди не накопичуються. Звідси виходить припущення, що фітофтора наділена здатністю руйнувати глікоалкалоїди.
В раневій тканині, зараженій несумісною расою фітофтори, не відбувається накопичення глікоалкалоїдів, не виявляються фітоaлексини рішитин і любімін. В непошкодженій паренхимі бульб не синтезуються ні глікоалкалоїди, ані фітоалексіни. В раневій перидермі весь мевалонат використовується на синтез глікоалкалоїдів. В відмерлій тканині, навпаки. Утворення глікоалкалоїдів майже не відбувається, а мевалонат використовується для синтезу рішитіна та любіміна. В зараженій тканині, як і в раневій, глікоалкалоїди хоча і утворюються, але розкладаються фітофторою [ ].
В США проводили роботи по виведенню сортів картоплі, наділених полігенною стікістю до фітофтори, при цьому стикалися з труднощами, обумовленими отриманням гібридів з високим вмістом в бульбах глікоалкалоїдів. Дослідження, проведені в селекційному центрі штата Західна Віргінія, на 15 клонах картоплі, наділених різною ступеню полігенної стійкості до фітофтори і трьох стандартних сортах, не підтвердили наявність взаємозв`язку між зазначеною стійкістю і вмістом в бульбах глікоалкалоїдів. В результаті прийшли до висновку, що можливе створення нових сортів картоплі з полігенною стійкістю до фітофтори і звичайним, або зниженим вмістом глікоалкалоїдів в бульбах. Встановлена висока ступінь кореляції (r=0,82) між вмістом глікоалкалоїдів в листках та бульбах випробуваного селекційного матеріалу [ ].
Дослідженнями Куна було встановлено, що непоїдання листків дикого виду картоплі Solanum demissum колорадським жуком обумовлена наявністю в них глікоалкалоїда демісіна [ ]. Це спонукало вітчизняних ботаніків організувати експедицію в країни Південної Америки для пошуків диких видів картоплі, стійких до колорадського жука. Під час експедиції було знайдено біля 10 таких видів, частина з них використовується в селекції при виведенні нових сортів картоплі.
Глікоалкалоїди бульбоносних видів дикої картоплі належать головним чином до групи соланіну чи демісину. Найбільша кількість стійких до колорадського жука видів картоплі – S.commersonii, S.chacoense, S.gibberulosum, S.shikii, S.dolichostigma відноситься до Commersonia, але демісин в них відсутній. Високостійкими до колорадського жука виявились також S.polyadenum, S.jamesii, котрі містять малу кількість демісину. Дикі види картоплі, які характеризуються низьким вмістом демісину, за виключенням видів Acaulia, виявились надто стійкими до колорадського жука [ ].
ПРИРОДНІ УМОВИ ГОСПОДАРСТВА
Виробнича практика була проведена в учбово-навчальному комплексі (УНВК) Сумського державного аграрного університету (СДАУ) який знаходится в м. Суми (південна частина). УНВК розташований у північній частині Лівобережного Українського Лісостепу, яка в цілому характеризується помірним кліматом. Літо – тепле, із значною кількістю опадів. Зима – не дуже холодна, з відлигами.
Згідно багаторічних даних Сумської метеостанції, середньорічна температура району, де розташований УНВК, дорівнює 60 С (табл. 1). Найбільш холодними місяцями є січень і лютий, найбільш теплими – липень, при чому абсолютний мінімум температур повітря -360 С спостерігається в січні, а максимум +380 С - у серпні. Останні весняні заморозки у повітрі спостерігаються у ІІІ декаді квітня, в окремі роки – на початку травня, а перші осінні заморозки – на початку жовтня, з
Середня багаторічна температура повітря за місяцями
|
Місяці |
І |
ІІ |
ІІІ |
IV |
V |
VI |
VII |
VIII |
ІХ |
Х |
ХІ |
ХІІ |
Середньо- річна |
|
Середньо-місячна температура повітря |
-7,8 |
-7,6 |
-2,6 |
6,3 |
14,6 |
17,4 |
19,6 |
18,3 |
13,0 |
6,6 |
-0,1 |
-5,5 |
6,0 |
к
 оливаннями
11.09. – 27.10. Довжина безморозного періоду
складає приблизно 155 днів. Зима сніжна,
характеризується нестійкою погодою,
поряд із низькими температурами -250
- -300 спостерігаються відлиги з
температурами +40 - +50.
оливаннями
11.09. – 27.10. Довжина безморозного періоду
складає приблизно 155 днів. Зима сніжна,
характеризується нестійкою погодою,
поряд із низькими температурами -250
- -300 спостерігаються відлиги з
температурами +40 - +50.
Часті відлиги взимку при глибокому промерзанні грунту іноді призводить до застою талих вод і утворенню льодяної кірки, що негативно впливає на перезимівлю озимих. Висота снігового покрову досягає 22 см. Розподілення його на більшій частині території нерівномірне: сніг накопичується у балках, ложбинах стоку.
Максимальна глибина промерзання грунту на рівних ділянках сягає 135 см, мінімально – 40 см, середня – 88 см.
Середньорічна кількість опадів становить 510 мм (табл. 2).
Середня багаторічна кількість опадів за місяцями
|
Місяці |
І |
ІІ |
ІІІ |
IV |
V |
VI |
VII |
VIII |
ІХ |
Х |
ХІ |
ХІІ |
За рік |
|
Опади в мм |
28 |
23 |
26 |
40 |
43 |
67 |
68 |
53 |
45 |
42 |
33 |
37 |
510 |
З таблиці видно, що найбільша кількість опадів випадає у літньо-весняний період, що співпадає з максимальним ростом сільськогосподарських культур і сівбою озимих і сприяє їх розвитку.
Взимку переважають північно-східний та північно-західний вітри. Напрямок їх часто змінюється, що призводить до різних змін температури. За останні роки спостерігаються суховійні південно-східні вітри, які сильно висушують грунт і знижують врожай основних сільськогосподарських культур.
Рельєф УНВК рівнинний, але західна частина дещо полога і засівається багаторічними травами.
Грунти, які представлені на дослідному полі, -це типові чорноземи, важкосуглинисті, середньогумусні на льосовидному суглинку. Грунт характеризується високим вмістом крупнопилови часток (0,01-0,05 мм), частка яких сягає 55% і більше. Вміст часток, які належать до фракції розміром 0,001 мм і менше сягає 25-30%, що обумовлює високу ємність поглинання, яка досягає 28-30 мг/екв на 100 г грунту. Реакція грунту практично нейтральна. Значення рН коливається в межах 6,5-7,5/6,5-7,6 у верхніх горизонтах і 7,3-7,5 у нижніх (карбонатних).
Розорювана частина грунту приблизно до 30 см., яка містить у собі необхідні елементи живлення (N, P, K) : азоту нітратного 2,2-3,; азота амонійного 11,2-10,6; фосфора 157, калія 70 мг на 100 г грунту, відсоток гумусу 4,0%.
УНВК заснований в 1987 р. для проведення науково-дослідної роботи та для демонстрації різних технологій вирощування сільськогосподарських культур. До складу УНВК входять комплекс по оздоровленню насінневої картоплі методами меристеми, обласне об’єднання “Сортнасіняовоч” і науково-дослідне об’єднання “Еліта” з лабораторією картоплі і дослідним господарством. До складу господарства також входять дослідні поля в с.Постольне, Гамаліївка та Косівщина, де вирощуються озима пшениця, овес, гречка, картопля, ячмінь, овес, гречка, горох, багаторічні трави, капуста, морква, столові буряки, цибуля. Урожайність деяких культур в 2000 році: озима пшениця 35 ц/га; овес 41 ц/га; гречка 20 ц/га; картопля 150 ц/га; соняшник 15 ц/га; яра пшениця 13 ц/га; горох 13 ц/га; буряки цукрові 460 ц/га; кукурудза на зерно – 42 ц/га.
Дослідні поля УНВК займають 43 га сільськогосподарських угідь. Сівозміна займає майже всі 43 га. Схема сівозміни така:
Овес 10 га.
Озима пшениця 5 га.
Картопля (насіння )0,7.
Ячмінь .
Гречка 4,6 га.
Цукровий буряк 10 га.
Із сівозміни виділена ділянка 2,5 га, розміщена на схилі, яка засівається багаторічними травами для тварин віварія.
Крім того в межах УНВК вирощували в 2000 р.такі культури:
Багаторічні трави 2,4 га
Молодий сад 3 га
Томати 0,2 га
Цибуля 0,25 га
Буряки столові 0,07 га
Морква 0,07 га
Кріп 0,01 га
Петрушка 0,01 га
Капуста 0,75 га
Огірки 0,05 га.
На дослідних ділянках проводять експериментальні роботи кафедри селекції (0,7 га) і рослинництва (0,3 га).
Для роботи на дослідних полях УНВК закріплений підрозділ і повний комплекс тракторів та сільськогосподарського обладнання. Для проведення дослідів виділено ділянки площею в 0,1 га. В склад підрозділу входять: завідуючий дослідним полем і два інженери. Для зберігання і ремонту техніки за дослідним полем закріплений спеціальний бокс, а для зберігання зернової продукції - ангар. Техніка УНВК представлена тракторами Т-150, Т-70, Т-150К, МТЗ-82, Т-25, Т-16.
3. МЕТОДИКА ПРОВЕДЕННЯ ДОСЛІДЖЕНЬ
В експериментi використовувались бульби та квітки картоплi сортів NewLeaf 6 Russet Burbank та NewLeaf 6 Atlantic (зразки люб’язно надані проф.А.А.Підгаєцьким, Інститут картоплярства УААН, смт.Немішаєве, Київська обл.). Зразки бульб: висічки сумарною масою 500 мг вiдбирали з 3 місць бульб картоплi (верхівка, середина та пуповинна частина). Маса зразків квіток дорівнювала 200-300 мг. Висічки бульб та зразки квіток мілко нарізували, перемішували і ретельно розтирали на холоді (0С) в порцеляновій чашці. В розтерту масу додавали 1 мл 5% оцтової кислоти. Ретельно перемішували одержану масу і залишали її стояти протягом 2-х годин. Після цього фільтрували масу, відбирали рідку фазу і підсушували її в струмені теплого повітря. Одержанi рідкофазні зразки центрифугували (300 g), вiдбирали алiквоти рідкої фази об'ємом 20 мкл і наносили на металеву (Au) поверхню зразконесучого диска приладу "МСБХ" (біохімічний мас-спектрометр «МСБX» виробництва ВAТ SELMI, Суми, Україна) з наступною реєстрацією мас-спектра (кiлькiсть вiдлiкiв 40000-200000); прискорююча напруга + 10 кВ)(контроль).
Аналiз одержаного мас-спектра проводили за допомогою сервiсної програми статистичної i математичної обробки спектрiв "МСБХ4" (НДI радіаційної технiки та автоматизації, Москва). Мас-спектри мiстили пiки квазимолекулярних iонiв (КМI) типу [М+Н]+, де М - молекулярна маса аналізованої речовини (глiкоалкалоїду) в атомних одиницях маси (Да), Н – протон, зокрема, -чаконіну з молекулярною масою 851,0 Да вiдповiдає пiк КМI з молекулярною масою 852,0; -соланiну - з масою 868,0.
Виходячи з спiввiдношень iнтенсивностi КМI глiкоалкалоїдiв проводили визначення кiлькостi останнiх в зразках. Для кожного зразка екстракту глiкоалкалоiдiв квіток рослин картоплi проводили по три вимiрювання з наступним усередненням результатiв за допомогою сервiсної програми.
Глікоалкалоїди для отримання калібрувальної кривої отримували з етиольованих ротків картоплі шляхом багаторазової обробки: екстрагування 2% оцтовою кислотою, осадження 25% водним рочином аміаку з підігріваням на водяній бані до випадання осаду, центрифугування, відділення осаду від надосадкової рідини, розчинення осаду в 2% оцтовій кислоті і т.д. до тих пір, поки не будуть отримані чисті кристали глікоалкалоїдів.
Визначення рівня стiйкостi сортiв картоплi до фiтопатогенiв в модельних системах in vitro визначали за методикою Кожушко Н.С., Чіванова В.Д. [ ]. Модельна система для визначення кінетичних параметрів деструкції глікоалкалоїдів складалась з 500 мг тканини бульби картоплі гомогенізованої в 5 мл середовища інкубації - 0,17% розчинi оцтової кислоти (pH 5,4-5,5), доповненої змiшаною польовою культурою грибiв Phytophthora infestans та Fusarium oxysporum spp.[ ]. За контроль правила гомогенізована тканина бульб в стерильному середовищі інкубації без домішок. Iнкубували модельнi системи при 37оC в термостаті, вiдбираючи зразки (100 мкл) на початку інкубації і в подальшому через 1, 2, 3, 4, 5 та 6 дiб. Одержанi зразки центрифугували, вiдбирали алiквоти рідкої фази об'ємом 20 мкл, які аналізували як вказано вище.
3. РЕЗУЛЬТАТИ ДОСЛІДЖЕНЬ
Мас-спектрометрія PDMS досить давно з успіхом використовується з метою аналіза кількісного та якісного складу глікоалкалоїдів рослин картоплі різних сортів [ ].
Так, в якості прикладу наводимо типовий мас-спектр свіжого екстракту з гомогенату бульб картоплі сорту Невська (рис.2). Порівняльна інтенсивність піків КМІ іонів a-соланіну та a-чаконіну в цьому мас-спектрі відповідає реальному кількісному співвідношенню цих сполук в тканинах бульб картоплі даного сорту – відомо, що для всіх культурних видів картоплі відношення a-чаконін : a-соланін дорівнює 2(3) : 1. Останнє вірно у тому разі, якщо в тканинах рослини картоплі даного сорту гідролітичні ферменти є помірно активними, тобто не піддають швидкій деструкції глікоалкалоїди шляхом відщеплення кінцевих залишків моносахаридів.
Крім інтенсивних піків КМІ, що відповідають a-соланіну та a-чаконіну в мас-спектрі присутні також «мінорні» пікі КМІ, які належать b-чаконіну (молекулярна маса 706 Да) та аглікону глікоалкалоїдів – соланідіну (молекулярна маса 398 Да)(рис.3). Зазначені сполуки виникають внаслідок перебігу процесів ферментативноі\хімічної деструкції інтактних молекул глікоалкалоїдів, які супроводжуються вiдщепленням вiд молекули a-чаконiну
L-Rha
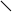 Solanidine -
D-Gal (Mолекулярна маса 851
Да)
Solanidine -
D-Gal (Mолекулярна маса 851
Да)
L-Rha
залишку рамнози (L-Rha) з утворенням b-чаконіну:
Solanidine - D-Gal - L-Rha (Mолекулярна маса 705 Да).
Подальша гідролітична деструкція b-чаконіну призводить до появи в середовищі інкубації вільного соланідину з молекулярною масою 398 Да. Незначна інтенсивність піків КМІ, що відповідають продуктам деструкції свідчить про те, що внутрішньотканинні ферменти і в першу чергу рамнозидаза, яка відщеплює кінцевий залишок рамнози від молекули глікоалкалоїду, знаходяться в інтактних тканинах рослин картоплі в неактивному стані. Як відомо, лише нативні молекули глікоалкалоїдів, проявляють притаманні їм потужні фунгіцидні властивості щодо фітопатогенів [ ]. Відповідно підвищення активності гідролітичних ферментів, індуковане механічними або хімічними чинниками, призводить залежно від глибини деструкційного процесу до часткової чи повної втрати глікоалкалоїдами фунгіцидної активності. Одним з таких чинників є фітопатогени: добре відомо, що бiохiмiчнi аспекти взаємодii фiтопатогенних грибiв з рослинними клiтинами передбачають як ключову компоненту детоксифiкуючий вплив спецiалiзованих ферментних систем грибiв на "захиснi" бiомолекули уражених рослин, зокрема глікоалкалоїди. Так, Phytophthora infestans dB та Fusarium oxysporum Schl. пiддають гiдролiтичнiй деструкцiї глiкоалкалоїди картоплi в процесi iнвазiї, спричинюючи таким чином втрату ними фунгiцидних властивостей [ ]. Дійсно, після 6-ти діб інкубації екстракта гомогенату з бульб картоплі сорту Невська в присутності фітопатогенів мас-спектр значно змінюється, а саме: інтенсивність піків КМІ a-соланіна і a-чаконіна різко знижується з паралельним зростанням інтенсивності відповідних піків КМІ, що належать b-чаконіну і соланідину (рис.3).
Якщо негативний вплив фітопатогенів «накладається» на притаманну конкретному сорту картоплі підвищену активність рамнозидази та споріднених ферментів, то сумація зазначених чинників створює оптимальні умови для повної нейтралізації фунгіцидної активності глікоалкалоїдів за короткий проміжок часу.
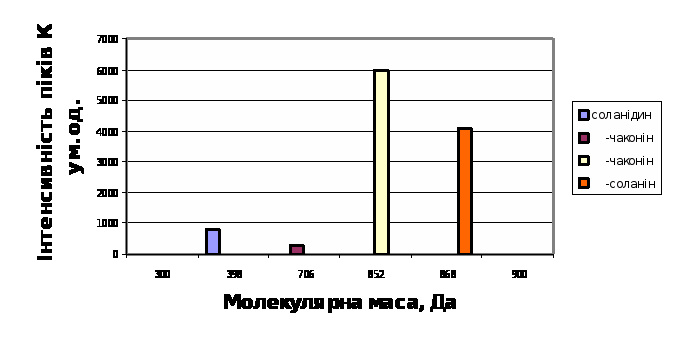
Рис.2. Мас-спектр екстракту з гомогенату тканин бульб картоплі сорту
Невська (контроль). Тут і на подальших рисунках інтенсивність
піків КМІ надана в умовних одиницях (ум.од.) – кількостях
відліків (стартів).
Рис. 3. Мас-спектр екстракту з гомогената тканин бульб картоплі сорту
Невська після 6-ти добової інкубації з фітопатогенами (Phytophthora
infestans dB і Fusarium spp. (дослід).
Навпаки, сорти картоплі, рослинам яких притаманна низька активність гідролітичних ферментів, протистоять фітопатогенам на протягом довшого проміжку часу, і є, таким чином, порівняно стійкими щодо фітопатогенів. Зважаючи на те, що ступінь активності ферментних систем є для сортів і гібридів генетично обумовленою ознакою, ми висловили припущення про наявність безпосереднього зв’язку між швидкістю деструкції молекул глікоалкалоїдів в тканинах рослини, обумовленою сумарною активністю ферментних систем рослини і фітопатогенів і ступенем стійкості окремих сортів картоплі до цих фітопатогенів. Нами досліджені кінетичні параметри ферментативної гідролітичної деструкції глікоалкалоїдів тканин бульб картоплі сортів Свiтанок київський, Невська, Молодiжна, та трансгенної картоплі NewLeaf 6 Russet Burbank та NewLeaf 6 Atlantic які значно різняться між собою за ступенем стійкості до фітопатогенів. Зокрема, Світанок київський порівняно стійкий до Phytophthora infestans dB та багатьох інших фітопатогенів; Невська, - середньостійка, а Молодіжна є сортом, малостійким до фітопатогенів. Щодо рослин трансгенної картоплі, то в літературі є свідчення на користь зниженого ступеня стійкості сортів NewLeaf 6 Russet Burbank та NewLeaf 6 Atlantic по відношенню до збудників хвороб.
Попередні експерименти з встановлення кількісних та якісних параметрів глікоалкалоїдів, притаманних рослинам трансгенних сортів картоплі, показали, що принципові відмінності останніх за зазначеними параметрами від класичних сортів відсутні (рис.4 і 5).
Як видно з рис.4 і 5, в тканинах квіток рослин картоплі сорту NewLeaf 6 Atlantic (рис.4) та NewLeaf 6 Russet Burbank (рис.5) присутні глікоалкалоїди α-соланін (868 а.о.м.) та α-чаконін (852 а.о.м.).
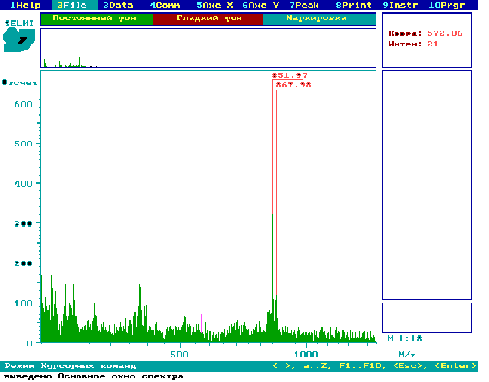
Рис.4. PDMS мас-спектр екстракту з квіток рослин картоплі сорту NewLeaf 6
Atlantic.
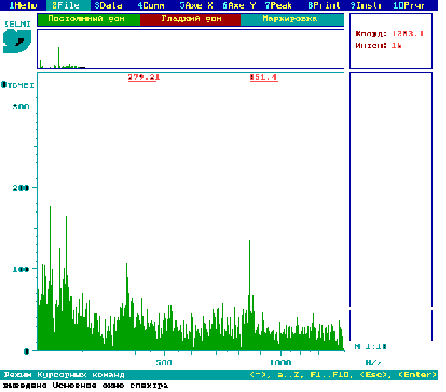
Рис.5. PDMS мас-спектр екстракту з квіток рослин картоплі сорту NewLeaf 6 Russet
Burbank.
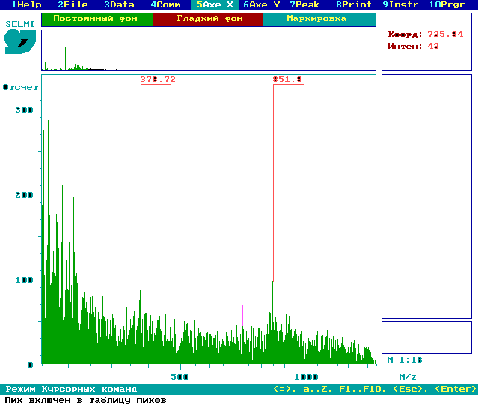
Рис.6. PDMS мас-спектр екстракту з бульб рослин картоплі сорту NewLeaf 6 Russet
Burbank.
Відносно вмісту глікоалкалоїдів в бульбах картоплі трансгенних сортів автор наводить інформацію про те, що ця група сполук присутня в бульбах трансгенної картоплі орієнтовно в тих же самих кількостях, як і в бульбах класичних сортів (рис.6).
Подальші експерименти показали вірність припущення про наявність прямого зв’язку між активністю гідролітичних ферментів рослин картоплі різних сортів та ступенем стійкості картоплі до фітопатогенів.
Із рис.7 легко бачити, що найвища швидкість гідролітичної деструкції як a–соланіна, так і a–чаконіна в середовищі інкубації притаманна малостійкому сортові картоплі Молодіжна; порівняно стійкий сорт Світанок київський характеризуються повільним зменшенням кількості нативних молекул глікоалкалоїдів в середовищі на протязі інкубації (рис.8). Середньостійкому сорту Невська притаманні проміжні кінетичні параметри гідролітичної деструкції глікоалкалоїдів, які є своєрідною комбінацією із відповідних параметрів стійкого та нестійкого до фітопатогенів сортів (рис.9).
Трансгенній картоплі сортів NewLeaf 6 Russet Burbank та NewLeaf 6 Atlantic притаманна підвищена в порівнянні з класичними сортами швидкість гідролітичної деструкції під впливом фітопатогенів (рис.10 і 11).
Таким чином з рис.7-11 випливає, що динамiка накопичення в модельнiй системi в процесi iнкубацii b-чаконiна безпосередньо корелює з рiвнем стiйкостi досліджених сортів картоплi по вiдношенню до фітопатогенів. Умовний ряд стiйкостi зазначених сортiв до збудників фітофторозу та фузаріозу, побудований на пiдставi мас-спектрометричних досліджень, в певному наближенні співпадає з аналогічним рядом, отриманим за допомогою класичних методів:
Свiтанок киiвський Невська Молодiжна NewLeaf 6 Atlantic > NewLeaf 6 Russet Burbank
¬ Рівень стійкості до фітопатогенів
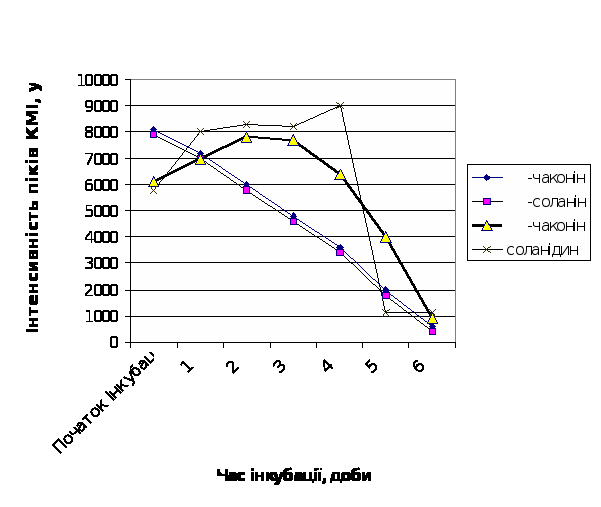
Рис.7. Динаміка деструкції глікоалкалоїдів в тканинах бульб картоплі
сорту Молодіжна.
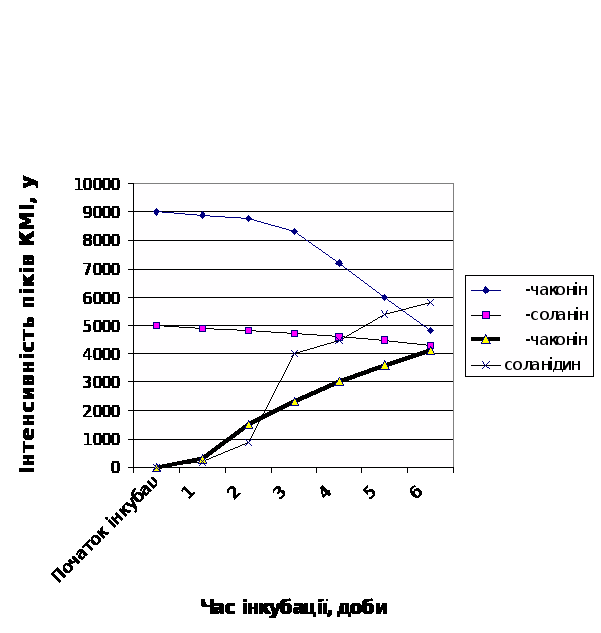
Рис.8. Динаміка деструкції глікоалкалоїдів в тканинах бульб картоплі
сорту Світанок київський.
Рис.9. Динаміка деструкції глікоалкалоїдів в тканинах бульб картоплі
сорту Невська.
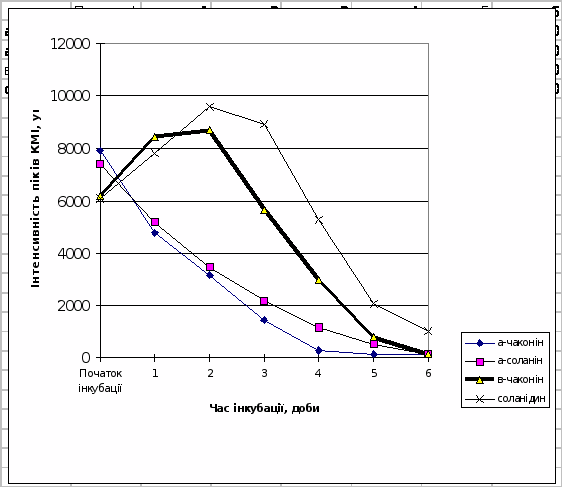
Рис.10. Динаміка деструкції глікоалкалоїдів в тканинах бульб картоплі
сорту NewLeaf 6 Russet Burbank.
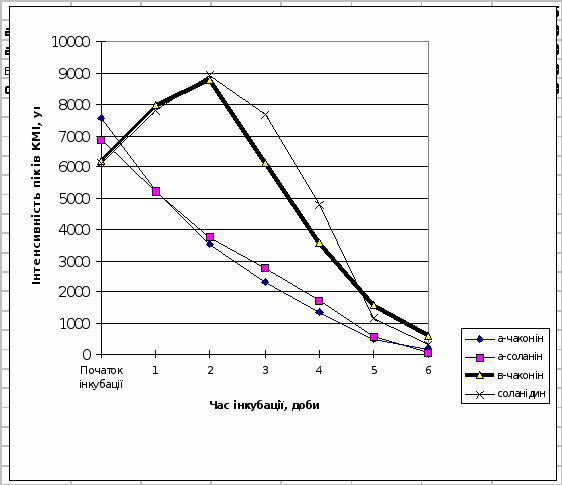
Рис.11. Динаміка деструкції глікоалкалоїдів в тканинах бульб картоплі
сорту NewLeaf 6 Atlantic.
ОХОРОНА ПРАЦІ
Охорона праці - це система законодавчих актів, соціально-економічних, організаційних, технічних, гігієнічних і лікувально-профілактичних заходів і засобів, спрямовані на створення безпечних умов, збереження здоров’я і працездатності людини в процесі праці. Крім того, охорона праці може розглядатися як науково обгрунтована соціально-технічна галузь досліджень, що вивчає теоретичні і практичні питання безпеки праці, запобігання виробничого травматизму, професійних захворювань і отруєнь, аварій, пожеж і вибухів на виробництві.
Законодавство про працю регламентується законодавчими актами, основними з яких є Конституція України, Кодекс законів про працю, Закон України «Про охорону праці».
В сільському господарстві необхідно створювати безпечні умови праці, грунтуючись на існуючих ДСТах, інструкціях та законах. Забезпечення задовільних умов праці по господарству в цілому залежить від керівників:
по галузях – від керівників відповідних підрозділів (агронома, інженера, зооінженера тощо);
по ділянках – від керівників нижчих ланок (бригадир, виконроб тощо).
З метою організації та контролю робіт в господарстві є посада інженера по охороні праці, щорічно складаються комплексні плани по організації служби охорони праці в господарстві. Головне в організації охорони праці – це своєчасне проведення інструктажів: вступного, первинного, повторного (через 6 місяців), позапланового та цільового.
Треба приймати до уваги, що найбільша кількість травм та пошкоджень відбувається при проведенні робіт, пов’язаних з ремонтом та поточним обслуговуванням сільськогосподарської техніки, навантажувальних та розвантажувальних роботах, внесенні мінеральних добрив, використанні пестицидів та роботах по збиранню врожаю.
Стан з охороною праці в УНВК наведені нижче(табл.5):
5. Показники травматизму по УНВК СДАУ
|
Показники травматизму |
Роки |
||
|
1998 |
1999 |
2000 |
|
|
Середньомісячна кількість працівників Кількість нещасних випадків, в тому числі із летальним фіналом Виділено коштів на охорону праці, грн. ( в якості матеріальної допомоги) |
13 2/0 548 |
9 1/0 705 |
15 3/0 843 |
В УНВК є різноманітні сільськогосподарські машини, які використовуються при вирощуванні картоплі: ДТ-75, МТЗ-80, Т-16, Т-150, К-701, котрі агрегатуються з культиваторами КРН-4,2, КПС-4, боронами БЗСС-1,0, котками ккш-6. При проведенні збиральних робіт застосовують комбайн ККУ-4, а при садінні-СКС-4. Особливе місце при плануванні заходів по охороні праці повинно приділятися роботам по застосуванні пестицидів та хімічних добрив. Пестициди в УНВК вносять за допомогою обприскувача ОПШ-15.
При роботі з причіпними та напівпричепними знаряддями для боронування та оранки, лущіння стерні, садіння, міжрядних обробок та інших процесів, необхідно суворо дотримуватись правил техніки безпеки.
Небезпечний виробничий фактор [ ] – це такий, дія якого на працюючого в певних умовах призводить до травм, або раптового погіршення стану здоров’я.
Наведемо небезпечні та шкідливі чинники, які притаманні технологіям вирощування картоплі (Табл.6).
6.Аналіз небезпечних та шкідливих виробничих чинників в технологіях вирощування картоплі.
|
Технологічні операції |
Небезпечні та шкідливі виробничі чинники |
|
Обробіток грунту |
Небезпека травмування навісними машинами під час регулювання, ремонту та очищення піднятої гідросистеми трактора. Небезпека при з’єднанні причіпних сільськогосподарських машин до трактора. При роботі агрегату певну шкоду організму завдають підвищені рівні шуму, вібрації та пилу. |
|
Внесення мінеральних добрив |
Особливо шкідливі азотні добрива: аміачна вода здатна спричинити опік шкіри, отруєння випарами аміаку, що потрапляють у повітря в кількостях, що значно перевищують ГДК. Також є небезпека отримати травму при навантажуванні та розвантажуванні добрив, при регулюванні та роботі агрегатів, що розкидують добрива. |
|
Садіння |
Небезпека при перевезенні мінеральних добрив та бульб; при регулюванні та навантажуванні саджалок. |
|
Обробіток посівів картоплі пестицидами |
Небезпека при приготуванні та перевезенні робочих рідин пестицидів, при регулюванні машин та обприскувачів. Існує реальна небезпека отруєння за умов перебільшення ГДК в навколишньому повітрі. |
|
Збирання врожаю |
Небезпека при налагодженні, регулюванні та процесі очищення від бадилля робочих органів картоплезбиральних комбайнів. Небезпека за умов присутності людей в робочій зоні картоплезбиральних машин, при навантаженні та розвантаженні бульб картоплі. |
Для створення кращих умов в господарстві необхідно усунути шкідливий вплив на організм людей різних виробничих факторів. При підготуванні машинно-тракторного агрегату необхідно перевірити комплектування і технічну справність машин [ ]. Не дозволяється робота машин на непідготовлених полях. При груповій роботі машин і агрегатів необхідно призначити старшого з числа працюючих. Забороняється при оранці, культивації, лущенні, боронувані, сідати на баластні ящики, знаходитися на рамі культиватора та в зоні роботи борін та дисків, а також очищати вручну робочі органи машин від бур’янів [ ]. При проведенні різних механізованих робіт на механізаторів та робітників в значній мірі впливають шуми та вібрація. Це призводить до швидкого стомлення, загальної слабкості, роздратування тощо. При неможливості знизити рівень виробничого шуму нижче встановлених гігієнічних норм застосовують протишумові засоби індивідуального захисту: навушники, вкладиші, шоломи, костюми [ ]. Для захисту від вібрації застосовують огороджуючі засоби, віброізоляцію, віброгасники. Для індивідуального захисту використовують взуття на вібропоглинаючій платформі, віброзахисні рукавиці.
Для роботи з пестицидами та мінеральними добривами не можуть бути допущені люди молодше 18 років, вагітні жінки, молоді матері та персонал, який не пройшов медичний огляд. При внесенні мінеральних та органічних добрив необхідно виконувати загальні правила техніки безпеки при роботі з розкидачами, транспортними засобами, завантажувачами. Забороняється: організовувати навантаження-розвантаження пилоподібних добрив з підвітреного боку; допускати до роботи з водним аміаком людей без засобів індивідуального захисту. Тривалість робочого дня при роботі з фосфорорганічними сполуками – 4 години (з доробкою 2 години на роботах, не пов’язаних з хімікатами).
Обов’язкове забезпечення працюючих на період робіт спецодягом, взуттям, респіраторами, захисними окулярами, рукавицями [ ].
Перед проведенням робіт робітники повинні пройти інструктаж, бути ознайомленими з препаратами та їх властивостями, а також мати спеціальну кваліфікацію.
Обробіток проводять лише відремонтованими та відрегульованими спеціальними машинами для обприскування (ОП-2000, ОПШ-15). Після проведення робіт обов’язково проводять знешкодження залишків пестицидів на обладнанні, тарі, одязі тощо.
Забороняється в місцях роботи з пестицидами зберігати продукти, воду, фураж, предмети домашнього вжитку, палити та харчуватись під час роботи, використовувати в господарстві тару з під пестицидів.
Перед застосуванням хімічних препаратів повідомляють населення найближчих населених пунктів. Всі роботи з пестицидами в спекотні пори року проводити виключно в ранкові часи та увечері, при більш низькій температурі повітря. Категорично забороняється перевозити людей на автомобільних і тракторних причіпах, автокранах, автонавантажувачах [ ].
Основні засоби захисту від ураження електричним струмом. Відповідно до правил влаштування електроустановок, їх необхідно надійно ізолювати, передбачити можливість випадкового дотику до зазначених установок. Обов’язково перевірити функціонування попереджуючої сигналізації. Відсутність можливості безпосереднього контакту з частинами установок та механізмів, що знаходяться під струмом, досягається використанням спеціального огороджування, розміщенням токопровідної мережі на недосяжній для працюючих висоті та використанням блокуючих пристроїв. До ізолюючих пристроїв належать ізолюючі штанги, ізолюючі лещата, діелектричні рукавиці та боти, гумові килимки, інструментарій із діелектричними ручками.
ОХОРОНА НАВКОЛИШНЬОГО СЕРЕДОВИЩА
Екологія – це наука про взаємодію тварин, рослин та мікроорганізмів між собою і біотичним середовищем, про зв’язки в надорганізмових системах – екосистемах, про структуру та функціонування екосистем (Ениоло В., 1998). Організація раціонального використання природніх ресурсів, надійного захисту навколишнього середовища, забезпечення вірних взаємовідносин людської спільноти та біосфери, що грунтується на науковій основі-одна з нагальних соціально-політичних проблем. Саме через це охорона природи – комплексне і довгострокове завдання, яке має безпосереднє відношенння до виробничих сил, науки, культури, та інших аспектів діяльності людини. З появою людини на планеті, велику роль у глобальній екосистемі стали відігравати взаємовідносини “суспільство-природа”. Особливо швидко посилюється вплив суспільства на природу в зв’язку з розвитком машинного виробництва. Завдяки цьому масштаби впливу суспільства на природу поширювались так швидко, що людство перетворилось у потужну геологічну силу, яка здійснює вплив на багато природних процесів. Необхідно відзначити, що проблеми навколишнього середовища не слід зводити тільки до негативного впливу людини на природу. Щорічно світова промисловість скидає в річки понад 160 м3 шкідливих стоків, щорічно в грунти вноситься 500 млн.т. мінеральних добрив і близько 4 млн.т. пестицидів, більша частина яких осідає в грунтах та виноситься поверхневими водами в річки, озера, моря та океани, в дуже значних кількостях накопичуюються в водосховищах для питної води для великих промислових міст.
Найбільш розповсюджений тип грунтів в УНВК – чорнозем типовий, малогумусний. В господарстві вирощуються такі культури: яра і озима пшениця, ячмінь, гречка, цукровий буряк, картопля. Для нормального росту і розвитку культурних рослин щільність грунту повинна становити 1,1-1,3 г/см3. При зростанні або зменшенні об’ємної маси грунту на 0,1-0,2 г/см3 порівняно з нормальною, врожай значно знижується. Ущільнений грунт погано вбирає і фільтрує вологу, що при зливах призводить до стоку та ерозії. На ущільнення грунту впливає також кількість прооджень сільгоспмашин та агрегатів. При підвищенні щільності грунту знижується загальна аерація, збільшується обсяг недоступної для рослин вологи, знижується її рухомість. У сучасному землеробстві хімізація сприяє максимальному використанню як природних факторів продуктивності рослин (грунтової родючості), так і агротехнічних (сівозміни, обробіток грунту, удобрення, боротьба з шкідниками тощо). До основних засобів хімізації при вирощуванні картоплі належать мінеральні добрива і пестициди. Несприятливий вплив добрив на навколишнє середовище може бути різним, але в основному внаслідок таких причин:
Надходження поживних елементів добрив з грунту у підгрунтові води з поверхневих вод може призвести до посиленого росту водоростей і утворення планктону.
Неправильне використання мінеральних добрив може погіршити кругообіг і баланс поживних речовин, агрохімічні властивості родючості грунту. Застосування азотних добрив при вирощуванні сільськогосподарських культур підвищує кислотність грунту.
Порушення живлення рослин мінеральними речовинами (мікро- і макроелементами) призводить до різних захворювань рослин, погіршує санітарний стан посівів.
Порушення технології застосування добрив, недосконалість якісних показників мінеральних добрив можуть зменшувати продуктивність сільськогосподарських культур і якість продукції, та призводити до накопичення в ній нітратів.
Однією з умов успішної рекультивації земель є вірне визначення товщини шару грунту, який потрібно знімати. Рекомендується знімати найбільш родючу частину грунтового покриву товщиною 45-50 см. Великим споживачем води є сільське господарство. 31 га посівів кукурудзи за вегетаційний сезон витрачают близько 3 тис.т., пшениці – 1,5; капусти – 3. У тваринництві при виробництві 1 т. м’яса витрачається 20 тис.м3 води. На підприємствах переробки сільськогосподарської продукції на виробництво 1 т. цукру потрібно 100 л. води. Значна кількість води витрачається при підтриманні санітарно-гігієнічних умов на теплиці для очищення приміщень та їх дезинфекції, підготування субстратів, миття посуду, тари, апаратури тощо. Внаслідок росту населення і розвитку продуктивних сил забезпечення людства водою стало екологічною проблемою. Скинуті у природні водоймища стічні води згубно діють на живі організми та значно погіршують гідрохімічний режим. Такі води необхідно очищати механічними, хімічними, фізико-хімічними та біологічними методами.
У зоні тваринницьких комплексів основними проблемами, які мають екологічне значення, є евтрофікація водоймищ, можливе накопичення патогенних мікроорганізмів, забруднення атмосферного повітря сірководнем, аміаком, молекулярним азотом та іншими сполуками. На атмосферу суттєво впливає використання і зберігання гною. При зберіганні його в ємкостях у великих кількостях випаровується в повітря аміак та його похідні. Тваринницькі комплекси забруднюють поверхневі водоймища, підземні води й грунт. При цьому в природних водоймищах гнойова рідина викликає масове отруєння водних організмів. Таким чином, існує необхідність розробки шляхів утилізації та раціонального використання відходів тваринництва. Гній зберігають у польових буртах і гноєсховищах. Сховища розміщують на відстані 3-50 м від тваринницьких приміщень і не менше, як за 200 м від помешкань. Основними джерелами забруднення атмосфери є природні, промислові і побутові процеси. Атмосфера має здатність до самоочищення, але в багатьох випадках ця потенційна можливість вже вичерпалась. Очищення повітря від пилу дає лише частковий ефект. Необхідні кардинальні заходи, на кшталт переходу до екологічно чистих видів енергії.
Для захисту навколишнього середовища в Україні прийнята низка законів:
“Закон України про охорону навколишнього середовища” (15.06.1991 р.);
“Закон України про охорону атмосферного повітря” (Київ, 1992 р.);
“Земельний кодекс України” (03.03.1993 р.);
“Водний кодекс України” (06.06.1991 р.).
Порушення цих законів може тягти за собою карну відповідальність.
На озброєнні сільського господарства є методи і засоби, які можуть успішно застосовуватись і застосовуються на практиці охорони природи.
В УНВК немає единої програми з охорони природи і тому постійне розширення виробничих потужностей обумовлює збільшення забрудненості відкритих водоймищ і грунтових вод, оскільки при будівництві тваринницьких об’єктів порушуються природоохоронні нормативи.
На землях УНВК мають місце прояви несприятливих природних та антропогенних процесів, які негативно впливають на стан грунтового покриву і стан сільськогосподарського виробництва в цілому. В ряді місць загальні кордони ріллі не зпівпадають з характером рельєфу території та ступінню еродованості грунтового покриву. Значна роздрібненість рельєфу, надмірна розораність, вирощування просапних культур на еродованих землях, а також значні відхилення від рекомендованої при попередньому землекористуванні системи природоохоронних заходів, сприяють розвитку водної ерозії.
В УНВК тваринницька галузь представлена ВРХ та молодняком ВРХ. Як вже зазначалося, продуктивність пасовищ є достатньою, але випасання тварин на сіножатях треба розпочинати після заготівлі сіна, а також обмежувати випасання в період посухи.
Лісосмуги представлені листяними породами. Конструкція лісосмуг - щільна. Вони знаходяться у задовільному стані. Територія господарства має насадження вздовж доріг та на вулицях робітничого селеща. На господарчому дворі, території промбази, дитячого садка та біля правління є газони та квітники.
Влітку добові витрати на 30% більші, ніж взимку. Джерелами стічної води є МТФ, житлові будинки, промислова база господарства. Від водоймищ ці об`єкти в середньому віддалені на незначну відстань - 400-500 м.
В робітничому селищі є колодязі, які віддалені від гноєсховищ, складів пестицидів та мінеральних добрив на значну відстань (понад 1,5 км). Якість води в них - добра, рівень води - 2-2,5 м, паводками не затоплюються.
Звалище має значну площу і організоване на місці кар`єру, тому значної загрози навколишньому середовищу не становить. Скотомогильники - огорожені, відалені від селищ, МТФ.
Основними стаціонарними джерелами забруднення повітря є котельня, а також звалище та автомобілі і трактори. Гази та аерозолі є основними типами речовин, що забруднюють атмосферу.
Населенний пункт господарства - не газифікований, тому повітря забруднюється більше. Сільськогосподарська техніка в цілому експлуатується екологічно безпечно, але трапляються випадки впливу паливно-мастильних матеріалів.
Основними заходами зменшення забруднення повітря в господарстві є газифікування населенного пункту, а також регулювання двигунів сільськогосподарських машин і механізмів.
На території УНВК мешкають дикі тварини: зайці, лисиці, куниці, різні птахи, комахи. Рослинніть - різнотравна - злакова. В травосуміші домінує злакова рослинність: топконіг лучний, вівсяниця борозчата, пирій повзучий та інші. Серед різнотрав`я, що займає 25-35% площі, трапляються подорожник середній, тисячолистник звичайний ті інші. Господарство має направлення зерно-м`ясо-молочне. В господарстві проводиться сівооранка зелених насаджень.
Як видно з наведеного матеріалу, стан охорони довкілля в господарстві - задовільний. Але сітуацію можна покращити:
1) проведення насадження на берегах водоймищ, крутих схилах балок та ярів дерев та кущів;
2) обмежити використання пестицидів, особливо сильнодіючих. Побудувати складські приміщення для зберігання пестицидів у відповідності до санітарних норм і вимог;
3) застосовувати у виробництві сучасну техніку.
ПРОПОЗИЦІЇ ВИРОБНИЦТВУ
Новітній метод м’якоіонізаційної мас-спектрометрії (PDMS) рекомендовано використовувати у великих спеціалізованих НДІ, що спеціалізуються на картоплярстві з метою дослідження трансгенних сортів картоплі та прогнозування рівня їх резистентності щодо фітопатогенів.
ВИСНОВКИ
Показана перспективність використання плазмено-десорбційної мас-спектрометрії в селекційній практиці з метою аналіза глікоалкалоїдів рослин картоплі трансгенних сортів;
М’якоіонізаційною мас-спектрометрією PDMS доведено, що кількісний та якісний склад глікоалкалоїдів в тканинах квіток та бульб картоплі трансгенних сортів NewLeaf 6 Russet Burbank і NewLeaf 6 Atlantic суттєво не відрізняється від аналогічних параметрів, притаманних сортам картоплі, отриманим за допомогою класичної селекції (Світанок київський, Невська, Молодіжна);
Надані теоретичні підстави притаманній картоплі трансгенних сортів NewLeaf 6 Russet Burbank і NewLeaf 6 Atlantic зниженій стійкості щодо фітопатогенів – глікоалкалоїди, як типові захисні вторинні метаболіти, деструкуються з найвищою швидкістю під впливом фітопатогенів саме в тканинах рослин картоплі трансгенних сортів.
СПИСОК ВИКОРИСТАНОЇ ЛІТЕРАТУРИ
Власенко М.Ю. Короткi пiдсумки та завдання дослiджень iз фiзiологii та бiохiмii картоплi. – Картоплярство. – 1994. – Вип.25. – С.14.
Власюк П.А., Власенко Н.Е., Мацко В.Н. Химический состав картофеля и путиулучшения его качества. - К.: Наукова думка, 1979. - 196 с.
Гусева А.Р., Пасешниченко В.А. Ферментативное расщепление гликоалкалоидов картофеля// Биохимия, 1957. - Т.22, N5. - C.843 - 845.
Иванов Б.И.. Пожарная безопасность в химических лабораториях. – М.: Химия,
1988. – 112 с.
Максимов М.Т., Оджагов Г.О. Радиоактивные загрязнения и их измерение. –
Инструкция по технике безопасности при хранении, транспортировке и применении пестицидов в сельском хозяйстве. – М.: Агропромиздат, 1985. – 56 с.
Инструкция по технике безопасности для сотрудников, аспирантов и студентов, работающих в лабораториях. – ССГІ: Суми, 1987. – 22 с.
Кинтя П.К. Строение и биологическая активность стероидных гликоалкалоидов ряда спиростана и фуростана. - Кишинев: Штиинца, 1987. - 141 с.
Кожушко Н.С., Чіванов В.Д. Экспресс-метод определения устойчивости картофеля к фитопатогенам в модельной системе// Современные технологии, экономика и экология в промышленности, на транспорте и в сельском хозяйстве: Сб. научн. ст. по материалам 5-й межд. научно-метод. конф. - К.: ИСМО, Алушта, 1998.
Конева И.М., Васюкова Н.И., Озерецковская О.Л. Возможный механизм индуцированной устойчивости картофеля к возбудителю фитофтороза // Пpикл.биохимия и микробиология.-T.30, N1. - C.161 - 166.
Липсиц Д.В. Биохимические основы болезнеустойчивости картофеля.- М.: ВНИИТЭИСХ, - 321 с.
Лукнер М.К. Вторичный метаболизм у микроорганизмов, растений и животных. - М.: Мир, 1979. - 550 с.
М.: Энергоатомиздат, 1986. – 224 с.
Методические рекомендации по проведению исследований с картофелем / Под
ред. Н.А.Пика. – К.: УкрНИИКХ, 1983. – 216 с.
14. Михайлов В.Н. и др. Охрана труда в сельском хозяйстве: Справочник.-М.: Агропромиздат, 1983.-99 с.
Новосельська А.П., Мицько В.М., Холодило І.В. Сортові особливості біохімічного складу бульб картоплі// Картоплярство. – 1995. – Вип.26. – С.42-45.
Оверчук В.И., Мицко В.Н. Гликоалкалоиды картофеля// Физиология и биохимия культ. растений. - 1972. -Т.4, N 5. - C. 492 - 498.
Оверчук В.И., Мицко В.Н. Гликоалкалоиды картофеля-. М.: ВНИИ ТЭИСХ, 1975. - 328 с.
Озерецковская О.Л., Давыдова М.А., Васюкова Н.И., Метлицкий Л.В. Гликоалкалоиды в здоровом и поврежденном клубне картофеля// ДАН СССР, Сер.биол., 1971. -T.196, N5. - C. 1470 - 1474.
Підгаєцький А.А. Використання генофонду картоплі для інтрогресії цінних генів при створенні вихідного селекційного матеріалу.// Автореф. … дис. док. сільськогосподарських наук. - К., 1993. – 44 с.
Прокошев С.М. Биохимия картофеля. - М.: Изд-во АН СССР, 1947. - 226 с.
Сикилинда В.А., Кирюхин В.П. Иследования гликоалкалолидов картофеля// Селекция и семеноводство картофеля. - 1975. N 21.- C. 5-11.
Справочник картофелевода// Под ред. А.И.Замотаева. - М.: В.О.Агропромиздат, 1987. - 351 с.
Фізіологія та біохімія картоплі / А.А.Кучко, М.Ю.Власенко, В.М.Мицько. – К.: Довіра, 1998. - 335 с.
Физиолого-биохимические и биофизические методы диагностики степени устойчивости растений к патогенам и другим факторам// Под ред. М.Е.Ладыгиной.- М.: Изд-во МГУ, 1992. - 96 с.
Харборн Д.И. Введение в экологическую биохимию. - М: Мир, 1985. - 312 с.
Чиванов В.Д. Определение содержания гликоалкалоидов и активности рамнозидазы в тканях картофеля методом времяпролетной плазменно-десорбционной масс-спектрометрии// Материалы Международной научной конференции «Актуальные проблемы современного картофелеводства"(Минск-Самохваловичи, 26-28.02.97 г.), с 89.
Чіванов В.Д. М’якоіонізуюча мас-спектрометрія в дослідженні глікоалкалоїдів картоплі. // Картоплярство. – К.: Урожай, 1999. - Вип. 29. – С.63-72.
Чіванов В.Д., Єрьоменко I.A. Вивчення динамiки деструкцii глiкоалкалоiдiв картоплi пiд дiєю фiтопатогенних грибiв мас-спектрометричним методом// Матерiали конференцii молодих вчених та спец-тiв "Науковi основи ведення картоплярства України в ринкових умовах" (22-24 травня 1996 р.), Немiшаєве. С.17.
ГОСТ (12.0.303-74) ССТБ. Опасные и вредные производственные факторы.
ГОСТ (12.4.011-75) ССТБ. Средства защиты работающих. Классификация.
ОСТ (46.3.1.108-81) ССТБ. Обработка почвы и посев зерновых культур. Требования безопасности.
ОСТ (46.0141-83) ССТБ. Процессы производственные в сельском хозяйстве. Общие требования безопасности.
ОСТ(46.3.1.161-84) ССТБ. Обработки пестицидами почвы и посевов.
ГОСТ (12.2.019-86) ССТБ. Трактора и машины самоходные сельскохозяйственные. Общие требования безопасности.
ГОСТ 12.3.041-86 ССВТ Применение пестицидов для защиты растений. Требования безопасности.
ГОСТ 46.0.126-82 ССБТ Организация обучения охраны труда в сельском хозяйстве. Общие положения.
Бурякова С.А. Охрана труда в сельском хозяйстве. - М.: Колос, 1991.- С. 22.
Винокуров Е.Н. Охрана труда в сельском хозяйстве. - К.: Урожай, 1992.- 120 с.
Гаврилов М.К. Охрана труда при интенсивной технологии. - М.: Колос, 1989.- 110 с.
Abdell D.C., Sporns P. Rapid quantitation of potato glycoalkaloids by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. // Journal of Agricultural and Food Chemistry.-1996.-V.44.- P.2292-2296.
Allen E.N., Kuс I. Steroid alkaloids in the disease resistance of white potato tubers// Phytopathology, 1964. - N.54. - P. 886 - 889.
Chazi V., Matthess D.P. Quantitative inhibitory effect of steroidal alkaloids: relative involvement of aglycones and sugar moieties on lettuce seed radicle elongation// Bot.Ganz. - 1990. - V.151, N 1. - P. 38 - 40.
Chen S., Derrick P.J., Mellon F.A., Price K.R. Analysis of glycoalkaloids from potato shoots and tomatoes by four sector tandem mass spectrometry with scanning-array detection. Comparison of positive ion and negative ion methods. // Analytical Biochemistry.-1994.-V.218.-P.157-69.
Chivanov V.D., Eriomenko I.A. PDMS Investigation into the Potato Cultivare Resistance to Рhytopathogenic Fungi // Abstr.14th Intern. Mass Spectrometry Conference, (25-29 August 1997).- Tampere, Finland.
Deahl К.А. Study of tle relationship of lateblight resistance to glycoalkaloid content in fifteen potato clones// Amer.Potato J. - 1973. -V.50. -P. 248 - 253.
Evans S., Buchanan R., Hoffman A. et al. Structural characterization of a glycoalkaloids at the femtomole level by means of four sector tandem mass spectrometry and scanning-array detection. // Organic Mass Spectrometry -1993.- V.28.-P. 289-290.
Friedman M., McDonald G.M. Potato Glycoalkaloids: Chemistry, Analysis, Safety, and Plant Physiology.// Critical Reviews in Plant Sciences.-1997.-V.16, №1.-P.55-132.
Hegnauer R. The taxonomic significance of alkaloids// Chemical Plant Taxonomy/ Ed. T.Swain.- N.Y.: Acad. Press, 1963.- P.389-427.
Highet R.J., Wheeler J.M. The study of alkaloid structures by spectral methods// The Alkaloids.- V.24./ Ed. A.Brossi.- N.Y.: Acad. Press, 1985. - P.287-348.
Holland H.L., Taylor G.J. Transformations of steroids and the steroidal alkaloid, solanine, by Phytophthora infestans// Phytochemistry.- 1979.- V.18. - P.437.
Houben R.I., Brunt K. Determination of glycoalkaloids in potato tubers by reversed-phase high-performance liquid chromatography // J.Chromatogr. A. - 1994.- V. 661. - P. 169 - 174.
Macfarlane R.D. et al. 252-Cf-Plasma Desorption Mass Spectrometry. II – A Perspective of New Directions// Biol. Mass Spectrometry. – 1994. – V.23. – 117-130.
Osmann S. et al. Glycoalkaloid composition of wild and cultivated tuber-bearing Solanum species of potenial value in potato breeding programs// J.Agr.Food Chem., 1978. – V.26. –N5. – P.1246-1248.
Paxton J.D., Groth J. Constraints on Pathogens Attacking Plants// Crit. Rev. in Plant Sci. – 1994. – V.13, –N 1. – P.77-95.
Price K.R., Mellon F.A., Self R., Fenwick G.R., Osman S.F. Fast bombardment mass spectrometry of Solanum glycoalkaloids and its potential for mixture analysis // Biomed. Mass Spectrom.-1985.-V.12.- P.79-85.
Ripperger H., Schreiber K. Solanum steroid alkaloids// The Alkaloids. -V.8.-/ Ed. R.G.A.Rodrigo.- N.Y.: Acad. Press, 1981.- P.81-192.
Roddick J.G., Melchers G. Steroidal glykoalkaloid content of potato, tomato and their somatic hybrids// Theor. Appl. Genet. – 1985. –V.80.- P.655-660.
Roddick J.G., Rijnenberg A.L.Synergistic interaction between the potato glycoalkaloids -solanine and -chaconine in relation to lysis of phospholipid/sterol liposomes. // Phytochemistry. – 1987. – V.26. – P.1325-1328.
Roddick J.G., Rijnenberg A.L., Osman S.F. Synergistic interaction between potato
glycoalkaloids -solanine and -chaconine in relation to destabilization of cell membranes:
ecological implications// J. Chem. Ecol. - 1988.- V.14.-P.889-902.
Sanford L.L., Deahl K.L., Sinden S.L. Glycoalkaloid content in foliage of hybrid and backross populations from a S.tuberosum ´ S.chacoense// Potato Journal, 1994. – V.71. – N4. – P.225-236.
Sharma R.P., Salunkhe D.K. Solanum Glycoalkaloids// In: Toxicant of Plant Origin. Vol.1. Alkaloids// Ed.P.R.Cheeke, Boca Raton, 1989.- P. 179 - 236.
Sinden S.L., Webb R.E. Effect of environment on glycoalkaloid content of six potato varieties at 39 locations// U.S. Dep. Agric. Tech. Bull. - 1974. -P.1472.
Sundqvist B.U.R. Modern Mass Spectrometry in Biological Sciences. - N.Y.:
Marcel Dekker, - P. 144 - 263.
Swaaij A.C. Effect of growth conditions on glycoalkaloid in potato tubers// Potato
Research, 1992. - V.35, N1. – P.68-69.
Swain A.P., Fitzpatrick T.J., Talley E.A., Herb S.F., Osman S.F. Enzymatic-hydrolysis of alpha-chaconine and alpha-chaconine// Phytochemistry.-1978.-V.17.-P.800.
The Biochemistry and Physiology of Plant Disease // Eds. R.N.Goodman, Z.Kiraly,
K.R.Wood. – Columbia.: University of Missouri Press, 1986. – 380 р.
Van Gelder W.M.J. Steroidal Glycoalkaloids in Solanum: Consequences for Potato
Breeding and for Food Safety//In. Handbook of Natural Toxins. – V.6. – Toxicology of
Plant and Fungal Compounds/Ed. R.F.Keeler, A.T.Tu. – N.Y.: Marcel Dekker, 1991. –
P.101-134.
Van Gelder W.M.J., Tuinistra L.G.M.Th., van der Greef J., Scheffer J.J.C. Characterization of novel steroidal alkaloids from tuber of Solanum species by combined gas chromatography-mass spectrometry. Implications for potato breeding. // J.Chromatogr.- 1989.-V.482.-P.13-22.
Zitnak A.The significance of glycoalkaloids in the potato plant// Proc. Can. Soc. Hortic. Sci., 1964.- V.3.-P.81.
Вельмишановний голово державної екзаменаційної комісії! Шановні члени державної екзаменаційної комісії! Колеги!
Тема моєї роботи: “Глікоалкалоїди трансгенних сортів картоплі: особливості метаболізму в умовах Північно-Східної України”. Важливість докладного дослідження в зазначеному напрямку обумовлене зокрема тим, що трансгенні сорти картоплі «Новий лист» виявили в умовах України підвищену чутливість до фітопатогенів і, внаслідок цього незадовільну придатність до зберігання протягом зимового періоду в картоплесховищах. З іншого боку, добре відомо, що одними з біохімічних компонентів рослин, що заслуговують на увагу, є саме глікоалкалоїди - сполуки, фізіологічна функція яких полягає у захисті рослин картоплі від фітопатогенів. Ми вважали, що глікоалкалоїди трансгенних сортів картоплі (або ферменти, які відповідають за метаболічні перетворення глікоалкалоїдів) мають деякі особливості, які можливо зкоррелювати з притаманною зазначеним сортам зниженою резистентністю щодо фітопатогенів.
Виходячи з наведеного, нами проведене дослідження стероїдних глікоалкалоїдів в квітках та бульбах рослин картоплі трансгенних сортів за допомогою мас-спектрометрії. Трансгенна картопля сортів NewLeaf 6 Russet Burbank і NewLeaf 6 Atlantic люб’язно надана професором Підгаєцьким з Інституту картоплярства НААН.
Мас-спектрометрія PDMS досить давно з успіхом використовується з метою аналіза кількісного та якісного складу глікоалкалоїдів рослин картоплі різних сортів. В нашому університеті розроблені оригінальні методики, за допомогою яких можна визначити, які саме глікоалкалоїди містяться в тканинах картоплі, а також встановити ступінь резистентності конкретних сортів щодо розповсюджених хвороб на основі параметрів кінетики деструкції, тобто руйнації, глікоалкалоїдів внаслідок дії ферментів.
Експерименти з встановлення кількісних та якісних параметрів глікоалкалоїдів, притаманних рослинам трансгенних сортів картоплі, показали, що принципові відмінності останніх за зазначеними параметрами від класичних сортів відсутні (рисунки).
Як видно з рисунку, в тканинах квіток рослин картоплі сорту NewLeaf 6 Atlantic та NewLeaf 6 Russet Burbank присутні глікоалкалоїди α-соланін (з масою 868 а.о.м.) та α-чаконін (з масою 852 а.о.м.), так само, як і в тканинах квіток звичайної картоплі. Кількість цих речовин в тканинах квіток трансгенних рослин також відповідає кількості глікоалкалоїдів в квітках пересічних культурних сортів. Але, не тільки сумарна кількість глікоалкалоїдів впливає на ступінь стійкості певних сорту картоплі, але важливим чинником є активність ферментних систем як самої рослини, так і фітопатогену, що руйнують молекули глікоалкалоїдів. Зокрема, відщеплення залишку цукру рамнози від молекули α-чаконіну призводить до втрати фунгіцидних властивостей. Таким чином, неважко зробити висновок про те, що чим активніші будуть відповідні ферменти, тим швидше буде зменшуватися в тканинах картоплі кількість нативних молекул глікоалкалоїдів і
Нами досліджені кінетичні параметри ферментативної гідролітичної деструкції глікоалкалоїдів тканин бульб картоплі сортів Свiтанок київський, Невська, Молодiжна, та трансгенної картоплі NewLeaf 6 Russet Burbank та NewLeaf 6 Atlantic які значно різняться між собою за ступенем стійкості до фітопатогенів. Зокрема, Світанок київський порівняно стійкий до Phytophthora infestans dB та багатьох інших фітопатогенів; Невська, - середньостійка, а Молодіжна є сортом, малостійким до фітопатогенів. Щодо рослин трансгенної картоплі, то в літературі є свідчення на користь зниженого ступеня стійкості сортів NewLeaf 6 Russet Burbank та NewLeaf 6 Atlantic по відношенню до збудників хвороб.
Показана перспективність використання плазмено-десорбційної мас-спектрометрії в селекційній практиці з метою аналіза глікоалкалоїдів рослин картоплі трансгенних сортів;
М’якоіонізаційною мас-спектрометрією PDMS доведено, що кількісний та якісний склад глікоалкалоїдів в тканинах квіток та бульб картоплі трансгенних сортів NewLeaf 6 Russet Burbank і NewLeaf 6 Atlantic суттєво не відрізняється від аналогічних параметрів, притаманних сортам картоплі, отриманим за допомогою класичної селекції (Світанок київський, Невська, Молодіжна);
Надані теоретичні підстави притаманній картоплі трансгенних сортів NewLeaf 6 Russet Burbank і NewLeaf 6 Atlantic зниженій стійкості щодо фітопатогенів – глікоалкалоїди, як типові захисні вторинні метаболіти, деструкуються з найвищою швидкістю під впливом фітопатогенів саме в тканинах рослин картоплі трансгенних сортів
ВІДГУК
на дипломну роботу студентки агрономічного факультету (спеціальність “Захист рослин”)
Загорулько О. “Глікоалкалоїди трансгенних сортів картоплі: особливості метаболізму в умовах Північно-Східної України”
Робота Загорулько О. присвячена актуальній темі - дослідженню метаболізму глікоалкалоїдів картоплі трансгенних сортів. Важливість докладного дослідження в зазначеному напрямку обумовлене зокрема тим, що трансгенні сорти картоплі «Новий лист» виявили в умовах України підвищену чутливість до фітопатогенів і, внаслідок цього незадовільну придатність до зберігання протягом зимового періоду в картоплесховищах. З іншого боку, добре відомо, що одними з біохімічних компонентів рослин, що заслуговують на увагу, є саме глікоалкалоїди - сполуки, фізіологічна функція яких полягає у захисті рослин картоплі від фітопатогенів. Виходячи з наведеного, автор вважає, що глікоалкалоїди трансгенних сортів картоплі (або ферменти, які відповідають за метаболічні перетворення глікоалкалоїдів) мають деякі особливості, які можливо зкоррелювати з притаманною зазначеним сортам зниженою резистентністю щодо фітопатогенів.
Виходячи з наведеного, Загорулько О. проведене дослідження стероїдних глікоалкалоїдів в квітках та бульбах рослин картоплі трансгенних сортів.
Дипломниця Загорулько О. сумлінно ставилася до виконання дипломної роботи, самостійно провела частину досліджень, опрацювала експериментальний матеріал і проаналізувала його.
Зважаючи на це, вважаємо, що дипломна робота Загорулько О. заслуговує на позитивну оцінку, а її авторка - на присвоєння кваліфікації «Вчений агроном по захисту рослин».
Доцент кафедри захисту
рослин, к.б.н. В.Д.Чіванов
РЕЦЕНЗІЯ
на дипломну роботу студентки агрономічного факультету (спеціальність “Захист рослин”)
Загорулько О. “Глікоалкалоїди трансгенних сортів картоплі: особливості метаболізму в умовах Північно-Східної України”
В останній час увага як фахівців в галузі сільського господарства, так і пересічних споживачів, прикута до проблеми трансгенних рослин та питань, пов’язаних з потенціальною токсичністю та мутагенністю продуктів харчування, виготовлених з вищезазначених рослин. Більшість дослідників вважає, що біохімічні параметри трансгенних рослин суттєво змінюються внаслідок введення до геному останніх чужорідних генів. Не виключено, що стан метаболізму таких типових вторинних метаболітів рослин картоплі, як стероїдні глікоалкалоїди, теж змінюється у трансгенних рослин. Виходячи з вищенаведеного, важко переоцінити актуальність дипломної роботи Загорулько О. Предметом дослідження є стан метаболізму глікоалкалоїдів картоплі трансгенних сортів картоплі «Новий лист» (“Монсанто”). Дипломницею Загорулько О. проведене докладне дослідження стероїдних глікоалкалоїдів в квітках та бульбах рослин картоплі трансгенних сортів. В роботі використані сучасні методи дослідження, а саме біологічна мас-спектрометрія.
Робота викладена на 58 сторінках комп’ютерного текста, містить усі необхідні розділи, ілюстрована 8 рисунками та 10 таблицями.
До недоліків дипломної роботи можна віднести невдалі літературні кліше, помилки в тексті тощо.
Виходячи з наведеного, вважаємо, що дипломна робота може бути допущена до захисту і заслуговує на позитивну оцінку за умов відповідної якості доповіді, а її авторка - на присвоєння кваліфікації «Вчений агроном по захисту рослин».