Макрофаги перитонеального экссудата как модель фагоцитоза и нарушений фагоцитарной активности

 Макрофаги
Макрофаги
С

 вободные
Резидентные
вободные
Резидентные
Перитонеальные Печеночные
Легочные
Активация – не только повышение активности и усиление метаболизма, цитотоксичности, но и увеличение числа вовлеченных в процесс клеток.
 Макрофаги
Макрофаги
Активированные 5% Интактные 95%
Активация
Специфическая Неспецифическая
(с помощью Тх1 и АТ) (разл. фарм. препараты, ЛПС, токсины)
Модель на перитонеальных МФ


 Среда 199
(пит.в-ва,
Среда 199
(пит.в-ва,
а/б, T=37°)
Регистрация данных
Прямой визуальный подсчет
Оценка хемотаксиса методом Бойдена
НТС-тест
Хемилюминесценция
Радиометрия
Ферментативные методы
Иммунологические методы



 Цитотоксичность
Цитотоксичность
Б ЦЖ,
Циклофосфамид (Активация)
ИЛ-1,
ФНО, Ростовые факторы, PG
E2
ЦЖ,
Циклофосфамид (Активация)
ИЛ-1,
ФНО, Ростовые факторы, PG
E2
Атипичные
клетки не
чувствительны
к данным агентам
Опыт с хитозаном
Макрофаг Т-лимфоцит


 Усиление
контактного взаимодействия тимоцитов
с макрофагами
ИЛ-2, ИФγ Активация
МФ
Усиление
контактного взаимодействия тимоцитов
с макрофагами
ИЛ-2, ИФγ Активация
МФ
Содержание
Краткий экскурс в историю…………………………………………………………………………………………….. 2
Современное состояние учения о фагоцитозе……………………………………………………………..5
Макрофаги перитонеального экссудата как модель
фагоцитоза и нарушений фагоцитарной активности…………………………………………….13
Получение модели………………………………………………………………………………………………………….14
Методы регистрации результатов…………………………………………....................................................14
Некоторые моделируемые процессы
СНИЖЕНИЕ БАКТЕРИАЛЬНОЙ АКТИВНОСТИ ПЕРИТОНЕАЛЬНЫХ
МАКРОФАГОВ МЫШЕИ В УСЛОВИЯХ СОЧЕТАННОГО
ПРИМЕНЕНИЯ СТАФИЛОКОККОВОГО ЭНТЕРОТОКСИНА ТИПА А И ЭНДОТОКСИНА………………………………………………17
ОТМЕНА УСИЛИВАЮЩЕГО ФАГОЦИТОЗ ДЕЙСТВИЯ ОПСОНИНОВ
С ПОМОЩЬЮ ФРАГМЕНТОВ АНТИТЕЛ ПРОТИВ Fc-РЕЦЕПТОРОВ МАКРОФАГОВ……………………………...................................18
УСИЛЕНИЕ С ПОМОЩЬЮ ХИТОЗАНА РЕАКЦИИ
КОНТАКТНОГО ВЗАИМОДЕЙСТВИЯ МАКРОФАГА С ТИМОЦИТАМИ in vitro…………………………………………………………..19
АКТИВАЦИЯ ФАГОЦИТАРНЫХ КЛЕТОК И КЛЕТОЧНОГО
ИММУНИТЕТА СИНТЕТИЧЕСКИМИ ПОЛИЭЛЕКТРОЛИТАМИ………………………………………………………………………………20
АКТИВАЦИЯ МАКРОФАГОВ ПОД ВЛИЯНИЕМ СИНТЕТИЧЕСКОГО АНТИОКСИДАНТА……………………………………………… 22
>ФАГОЦИТАРНАЯ АКТИВНОСТЬ МАКРОФАГОВ >
>ПЕРИТОНЕАЛЬНОГО ЭКССУДАТА МЫШЕЙ ДЕЙСТВИИ ПРЕПАРАТОВ ПЛАТИНЫ………………………………………………………>>23>
ИЗУЧЕНИЕ ФАГОЦИТАРНОЙ АКТИВНОСТИ ПЕРИТОНЕАЛЬНЫХ МАКРОФАГОВ В
ОТНОШЕНИИ YERSINIA PESTIS С ДЕФЕКТНЫМИ И ПОЛНОЦЕННЫМИ FRA-ГЕНАМИ…………………………………………………25
ВЛИЯНИЕ МОДИФИКАТОРОВ БИОЛОГИЧЕСКОГО ОТВЕТА ПРИРОДНОГО
ПРОИСХОЖДЕНИЯ НА ФУНКЦИОНАЛЬНУЮ АКТИВНОСТЬ МАКРОФАГОВ……………………………………………………………….26
ПЕРИТОНЕАЛЬНЫЕ МАКРОФАГИ КАК МОДЕЛЬ
ДЛЯ ИЗУЧЕНИЯ АТЕРОГЕННОГО ПОТЕНЦИАЛА СЫВОРОТКИ КРОВИ……………………………………………………………………...29
ВЛИЯНИЕ ГАМК, ГОМК И ГЛУТАМИНОВОИ
КИСЛОТЫ НА ФУНКЦИОНАЛЬНУЮ АКТИВНОСТЬ ФАГОЦИТОВ……………………………………………………………………………32
Заключение………………………………………………………………………. ……………………………………………………………33
Некоторые другие модели изучения фагоцитоза……………………………………………………………… 34
Литература…………………………………………………………………………………………………………………………………………36
Краткий экскурс в историю
Более 100 лет прошло с момента открытия фагоцитарной теории, созданной нашим великим натуралистом, лауреатом Нобелевской премии И. И. Мечниковым. Открытие, осмысление явления фагоцитоза и формулирование в общих чертах основ фагоцитарной теории было сделано им в декабре 1882 г. В 1883 г. он изложил основы новой фагоцитарной теории в докладе «О целебных силах организма» в Одессе на VII съезде естествоиспытателей и врачей и опубликовал их в печати. Были впервые высказаны основные положения фагоцитарной теории, которые И. И. Мечников развивал в последующем на протяжении всей своей жизни. Хотя сам факт поглощения живыми клетками других частиц был описан многими натуралистами задолго до ученого, однако только он дал блестящее толкование огромной роли фагоцитов в защите организма от болезнетворных микробов.
Много позже к 70-летнему юбилею ученого коллега и друг И. И. Мечникова Эмиль Ру напишет: «Сегодня, мой друг, Вы наблюдаете доктрину фагоцитоза со спокойным удовлетворением отца, дитя которого сделало хорошую карьеру в мире, но сколько беспокойств оно Вам доставило! Его появление вызвало протесты и сопротивление и в течение двадцати лет Вам пришлось сражаться за него». Доктрина фагоцитоза «...одна из наиболее плодотворных в биологии: она связала явление иммунитета с внутриклеточным пищеварением, она объяснила нам механизм воспаления и атрофии; она оживила патологическую анатомию, которая, не будучи в состоянии дать приемлемое объяснение, оставалась чисто описательной... Ваша эрудиция такая обширная и верная, что она служит всему миру».
И. И. Мечников утверждал, что «...иммунитет в инфекционных болезнях должен быть приписан активной целлюлярной деятельности. Среди клеточных элементов фагоциты должны занять первое место. Чувствительность и подвижность, способность поглощать твердые тела и вырабатывать вещества, могущие разрушать и переваривать микробов — вот главные факторы деятельности фагоцитов. Если эти свойства в достаточной мере развиты и парализуют патогенное действие микробов, тогда животное от природы иммунно... когда фагоциты не обнаруживают наличия всех или одного из этих свойств в достаточной степени, то животное восприимчиво к инфекции...». Вместе с тем, если бактериальные продукты вызывают у фагоцитов отрицательный хемотаксис или, если при положительном хемотаксисе фагоциты не поглощают бактерий или поглощают, но не убивают их, — также развивается смертельная инфекция. Решение фундаментальных проблем сравнительной эмбриологии и биологии, приведшее к крупнейшим открытиям ученого, позволило И. И. Мечникову установить, что «фагоцитоз чрезвычайно распространен в животном мире... как на самой низшей ступени животной лестницы, например, у простейших, так и ...у млекопитающих животных и человека... фагоциты представляют собой мезенхимальные клетки».
И. И. Мечников был в то же время первым, кто занялся сравнительным изучением явления фагоцитоза. Внимание ученого было обращено не только на традиционные лабораторные объекты, но и на таких представителей мира животных, как дафнии, морские звезды, крокодилы, обезьяны. Сравнительное изучение фагоцитоза было необходимо И. И. Мечникову для доказательства всеобщности явлений поглощения и разрушения чужеродного материала фагоцитирующими мононуклеарами, широкого распространения в природе изучаемой им формы иммунологической защиты.
Клеточная теория Мечникова сразу наткнулась на сопротивление. Прежде всего она была предложена в то время, когда большинство патологов видели в воспалительной реакции, а также в связанных с ней микрофагах и макрофагах не защитную, а вредоносную реакцию. В то время считали даже, что, хотя фагоцитирующие клетки действительно способны поглощать болезнетворные микроорганизмы, это приводит не к разрушению возбудителя, а к переносу его в другие части тела и распространению болезни. Также в тот период времени интенсивно развивалась гуморальная теория иммунитета, основы которой были заложены П.Эрлихом. Были открыты антитела и антигены, были выявлены механизмы гуморальной устойчивости организма против некоторых патогенных микроорганизмов и их токсинов (дифтерия, столбняк и др.). Как это ни странно, но два таких открытия не могли некоторое время ужиться вместе. Позднее в 1888 г. Наттолл нашел в сыворотке нормальных животных вещества, токсичные для некоторых микроорганизмов, и показал, что такие антибактериальные свойства значительно повышаются в результате иммунизации животного. В дальнейшем было обнаружено, что в сыворотке имеются два разных вещества, совместное действие которых приводит к лизису бактерий: термостабильный фактор, затем идентифицированный как сывороточные антитела, и термолабильный фактор, названный комплементом, или алексином (от греч. aleksein - защищать). Ученик самого Мечникова Борде описал лизис эритроцитов гуморальными антителами и комплементом, и большинство исследователей стали соглашаться с Кохом, что победу одержали гуморалисты. Мечников и его ученики отнюдь не собирались сдаваться. Были поставлены простые опыты, в которых микробы, помещенные в маленький мешочек из фильтровальной бумаги, защищающий их от фагоцитов, сохраняли свою вирулентность, хотя буквально купались в тканевой жидкости, богатой антителами. В Англии сэр Элмрот Райт и С. Р. Дуглас попытались примирить различия между этими двумя школами в своих капитальных исследованиях процесса опсонизации (от греч. opsonein - делать съедобным). Эти ученые утверждали, что клеточный и гуморальный факторы являются одинаково важными и взаимозависимыми в том отношении, что гуморальные антитела, специфически реагируя со своей мишенью - микроорганизмом, подготавливают его к фагоцитозу макрофагами.
В 1908 г. Шведская академия удостоила Нобелевской премии по медицине совместно Мечникова - основателя клеточного направления и Эрлиха - олицетворявшего гуморалистские идеи того времени. Они были удостоены премии в качестве «признания их работ по иммунитету».
Заслуга Мечникова состоит не только в создании им гениальной теории. Еще ранее он начал изучать заразные болезни человека и домашних животных: вместе со своим учеником Н. Ф. Гамалеей он изучал туберкулез, чуму рогатого скота, искал способы борьбы с вредителями сельского хозяйства. К 1886 г. относится одно из важнейших событий в истории русской медицины. Летом этого года в Одессе начала работать созданная Мечниковым и его талантливым учеником Н. Ф. Гамалеей первая русская бактериологическая станция. Он создал в России крупнейшую научную школу микробиологов. Выдающиеся ученые Н. Ф. Гамалея, Д. К. Заболотный, Л. А. Тарасевич и многие другие были учениками И. И. Мечникова. Илья Ильич Мечников умер в 1916 году, до конца жизни занимаясь вопросами иммунологии и клеточного иммунитета. А наука об иммунитете быстро и стремительно развивалась. В этот период было необычайно много работ и ученых, изучавших факторы внутренней защиты организма.
Период с 1910 по 1940гг. был периодом серологии. В это время было сформулировано положение о специфичности и о том, что АТ являются естественными, высоковариабельными глобулинами. Большую роль здесь сыграли работы Ландштейнера, который пришел в выводу, что специфичность антител не является абсолютной.
С 1905 появились работы (Сarrel, Guthrie) по трансплантации органов. В 1930г. К.Ландштейнер открывает группы крови. Работами по фагоцитозу, бактериофагии, вирусам, патогенезу чумы занимается Амадей Боррель. Премия присуждена Ф. Макфарлейну Бернету (1899 - 1985) и Питеру Медавару (1915 - Англия) «за открытие приобретенной иммунолотической толерантности». Медавар показал, что отторжение чужеродного кожного трансплантата подчиняется всем правилам иммунологической специфичности, и в основе его лежат такие же механизмы, как и при защите от бактериальных и вирусных инфекций. Последующая работа, которую он провел вместе с рядом учеников, заложила прочную основу для развития трансплантационной иммунобиологии, которая стала важной научной дисциплиной и в дальнейшем обеспечила многие достижения в области клинической трансплантации органов. Бернет опубликовал книгу «Образование антител» (1941 г.). Со своим коллегой, Франком Феннером, Бернет утверждал, что способность к иммунологическим реакциям возникает на сравнительно поздних стадиям эмбрионального развития и при этом происходит запоминание существующих маркеров «своего» у антигенов, присутствующих в данный момент. Организм в последующем приобретает к ним толерантность и не способен отвечать на них иммунологической реакцией. Все антигены, которые не запомнились, будут восприниматься как «не свои» и смогут в дальнейшем вызывать иммунологический ответ. Было высказано предположение, что любой антиген, введенный в течение этого критического периода развития, будет затем восприниматься как свой и вызывать толерантность, в результате чего не сможет в дальнейшем активировать иммунную систему. Эти идеи были далее развиты Бернетом в его клонально-селекционной теории образования антител. Предположения Бернета и Феннера были подвергнуты экспериментальной проверке в исследованиях Медавара, который в 1953 г. на мышах чистых линий получили четкое подтверждение гипотезы Бернета - Феннера, описав феномен, которому Медавар дал название приобретенной иммунологической толерантности.
В 1969г. одновременно несколькими авторами (Р.Петров, М.Беренбаум, И.Ройт) была предложена трехклеточная схема кооперации иммуноцитов в иммунном ответе (Т-, В-лимфоцитов и макрофагов), определившая на многие годы изучение механизмов иммунного ответа, субпопуляционной организации клеток системы иммунитета.
Существенную роль в этих исследованиях сыграли кинематографические методы. Возможность непрерывного динамического изучения микробиологических объектов in vivo и in vitro в условиях, совместимых с их жизнедеятельностью, визуализация невидимых для человеческого глаза электромагнитных излучений, регистрация как быстрых, так и медленных процессов, управление масштабом времени и некоторые другие характерные особенности исследовательской кинематографии открыли большие и во многих отношениях уникальные возможности для изучения взаимодействия клеток.
Представление о фагоцитах за истекшее время подверглось существенной эволюции. В 1970 г. Van Furth и соавт. предложили новую классификацию, выделяющую МФ из РЭС в отдельную систему мононуклеарных фагоцитов. Исследователи отдали дань уважения И. И. Мечникову, пользовавшемуся термином «мононуклеарный фагоцит» еще в начале XX века. Фагоцитарная теория не стала, однако, неизменяемой догмой. Непрерывно накопляемые наукой факты изменили и усложнили понимание тех явлений, в которых фагоцитоз казался решающим или единственным фактором.
Можно утверждать, что в наши дни созданное И. И. Мечниковым учение о фагоцитах переживает свое второе рождение, новые факты значительно обогатили его, показав, как это и предсказывал Илья Ильич, огромное общебиологическое значение. Теория И. И. Мечникова явилась мощным индуктором прогресса иммунологии во всем мире, большой вклад в него внесли советские ученые. Однако и сегодня основные положения теории остаются незыблемыми.
Первостепенное значение фагоцитарной системы подтверждается созданием в США общества ученых, занимающихся изучением ретикулоэндотелиальной системы (РЭС), издается специальный «Journal of Reticulo-Endothelial Society».
В последующие годы развитие фагоцитарной теории связано с открытием цитокиновой регуляции иммунного ответа и, конечно, изучения влияния цитокинов на клеточный ответ в том числе и макрофагов. На заре этих открытий стоя ли работы таких ученых, как Н.Ерне,
Г. Келер, Ц. Милштейн.
В СССР бурный интерес к фагоцитам и связанным с ними процессами наблюдался в 80-е годы. Здесь необходимо отметить работы А.Н.Маянского, изучавшего влияния макрофагов не только в свете их иммунной функции. Он показал значение клеток РЭС на функционирование таких органов как печень, легкие, желудочно-кишечный тракт. Работы также проводили А.Д. Адо, В.М.Земсков, В.Г.Галактионов, эксперименты по изучению работы МФ в очаге хронического воспаления ставил Серов.
Следует сказать, что в 90-е годы интерес к неспецифическому звену иммунитета упал. Отчасти это можно объяснить тем, что все усилия ученых были в основном устремлены к лимфоцитам, но особенно – к цитокинам. Можно сказать, что сейчас продолжается «цитокиновый бум».
Однако это ни в коем случае не означает, что актуальность проблемы упала. Фагоцитоз составляет пример того процесса, интерес к которому не может пропасть. Будет открытие новых факторов стимулирующих его активность, будут обнаружены вещества угнетающие РЭС. Будут открытия, уточняющие тонкие механизмы взаимодействия МФ с лимфоцитами, с клетками интерстиция, с антигенными структурами. Особенно это может быть актуально сейчас в связи с проблемой опухолевого роста и СПИД’а. Остается надеяться, что в ряду открытий, начатых великим Мечниковым, будут стоять имена русских ученых.
СОВРЕМЕННОЕ СОСТОЯНИЕ УЧЕНИЯ О ФАГОЦИТОЗЕ
Основные положения о фагоцитах и системе фагоцитоза, блестяще сформулированные И. И. Мечниковым и разработанные его учениками и последователями, надолго определили развитие этого важнейшего направления биологии и медицины. Идея о противоинфекционном иммунитете, которая так увлекла современников И. И. Мечникова, сыграла решающую роль в становлении клеточной иммунологии, эволюции взглядов на воспаление, физиологию и патологию реактивности и резистентности организма. Парадоксально и вместе с тем закономерно, что учение о фагоцитозе началось с крупных обобщений и концепций, которые на протяжении многих лет дополнялись фактами частного характера, мало повлиявшими на развитие проблемы в целом. Волна современной иммунологической информации, изобилие изящных методов и гипотез направили интересы многих исследователей в сторону изучения лимфоцитарных механизмов клеточного и гуморального иммунитета. И если иммунологи быстро поняли, что без макрофага им не обойтись, то судьба другого класса фагоцитирующих клеток — полинуклеарных (сегментоядерных) лейкоцитов — до недавнего времени оставалась неясной. Только теперь можно с уверенностью сказать, что и эта проблема, сделав за последние 5—10 лет качественный скачок, прочно утвердилась и успешно развивается не только иммунологами, но и представителями смежных профессий — физиологами, патологами, биохимиками, клиницистами. Изучение полинуклеарных фагоцитов (нейтрофилов) — один из немногих примеров в цитофизиологии, а тем более в иммунологии, когда число исследований на объекте «человеческого происхождения» превосходит количество работ, выполненных в эксперименте на животных.
Сегодня учение о фагоцитозе — это совокупность представлений о свободных и фиксированных клетках костномозгового происхождения, которые, обладая мощным цитотоксическим потенциалом, исключительной реактивностью и высокой мобилизационной готовностью, выступают в первой линии эффекторных механизмов иммунологического гомеостаза. Противомикробная функция воспринимается как частный, хотя и важный, эпизод этой общей стратегии. Доказаны мощные цитотоксические потенции моно- и полинуклеарных фагоцитов, которые, кроме бактерицидности, находят выражение в уничтожении малигнизированных и иных форм патологически измененных клеток, альтерации тканей при неспецифическом воспалении в иммунопатологических процессах. Если нейтрофилы (доминирующий тип полинуклеаров) почти всегда нацелены на деструкцию, то функции мононуклеарных фагоцитов сложнее и глубже. Они участвуют не только в разрушении, но и в созидании, запуская фибробластические процессы и репаративные реакции, синтезируя комплекс биологически активных субстанций (факторы комплемента, индукторы миелопоэза, иммунорегуляторные белки, фибронектини пр.). Сбывается стратегический прогноз И. И. Мечникова, который всегда смотрел на фагоцитарные реакции с общефизиологических позиций, утверждая значение фагоцитов не только в защите от «вредных деятелей», но и в общей борьбе за гомеостаз, которая сводится к поддержанию относительного постоянства внутренней среды организма. «При иммунитете, атрофии, воспалении и излечении, во всех явлениях, имеющих величайшее значение в патологии, участвуют фагоциты».
Мононуклеарные фагоциты, которые ранее относили к ретикулоэндотелиальной системе, выделены в самостоятельное семейство клеток — систему мононуклеарных фагоцитов, которая объединяет моноциты костного мозга и крови, свободные, и фиксированные тканевые макрофаги. Доказано, что, выходя из крови, моноцит меняется, адаптируясь к условиям той среды, в которую попадает. Это обеспечивает специализацию клетки, т. е. максимальное соответствие тем условиям, в которых ей предстоит «работать». Не исключена и другая альтернатива. Подобие моноцитов может быть чисто внешним (как это случилось с лимфоцитами), и часть из них предетерминирована к трансформации в различнее варианты макрофагов. Гетерогенность зрелых нейтрофилов хотя и существует, но выражена гораздо слабее. Они почти не меняются морфологически, попадая в ткани, в отличие от макрофагов живут там недолго (не более 2—5 сут) и явно не обладают пластичностью, присущей моноцитам. Это высокодифференцированные клетки, которые практически заканчивают свое развитие в костном мозге. Не случайно, известные в прошлом попытки отыскать корреляцию между сегментацией ядра и способностью лейкоцитов к фагоцитозу не увенчались успехом. Тем не менее идея о функциональной неоднородности морфологически зрелых нейтрофилов продолжает получать подтверждения. Известны различия между нейтрофилами костного мозга и периферической крови, нейтрофилами крови, тканей и экссудатов. Причины и физиологический смысл этих особенностей неизвестны. По-видимому, изменчивость полинуклеаров в отличие от моноцитов-макрофагов носит тактический характер.
Изучение фагоцитоза ведется согласно классическим постулатам И. И. Мечникова о фазах фагоцитарной реакции — хемотаксису, аттракции (связывании)> >и поглощении>,> уничтожении (переваривании). К характеристике каждого из этих процессов в настоящее время приковано внимание, им посвящают монографии, обзоры. Результаты многочисленных исследований позволили углубиться в суть этих реакций, конкретизировать молекулярные факторы, лежащие в их основе, нащупать общие узлы и вскрыть частные механизмы клеточной реактивности. Фагоцитоз служит прекрасной моделью для изучения миграционной функции, пространственной ориентации клеток и их органелл, слияния и новообразования мембран, регуляции клеточного гомеостаза и других процессов. Иногда фагоцитоз нередко отождествляют с поглощением. Это явно неудачно, ибо нарушает исторически сложившееся представление о фагоцитозе как об интегральном процессе, который объединяет сумму клеточных реакций, начиная с распознавания объекта и кончая его разрушением или стремлением к разрушению. С функциональной точки зрения фагоциты могут пребывать в двух состояниях — покоящемся и активированном. В наиболее общем виде активация — есть результат преобразования внешнего стимула в реакцию эффекторных органелл. Больше пишут об активированном макрофаге, хотя в принципе то же самое можно сделать и для полинуклеаров. Надо выбрать лишь точку отсчета — к примеру, функциональный статус в сосудистом русле нормального организма. Активация различается не только степенью возбуждения индивидуальных клеток, но и масштабом охвата клеточной популяции в целом. В норме активировано небольшое количество фагоцитов. Появление раздражителя резко меняет этот показатель, отражая подключение фагоцитов к реакциям, корригирующим внутреннюю среду организма. Стремление проактивировать фагоцитарную систему, усилив тем самым ее эффекторные возможности, неоднократно звучало в работах И. И. Мечникова. Современные исследования по адъювантам, биологическим и фармакологическим модуляторам мононуклеарных и полинуклеарных фагоцитов по существу развивают эту мысль с позиций межклеточной кооперации, общей и частной патологии. В этом видится перспектива рационального воздействия на воспаление, репаративные и регенеративные процессы, иммунопатологию, резистентность к острому и хроническому стрессу, устойчивость к инфекциям, опухолям и пр.
Многие признаки активации стереотипны, повторяясь у всех фагоцитирующих клеток. К ним относятся изменение активности лизосомальных и мембранных ферментов, усиление энергетического и окислительного метаболизма, синтетических и секреторных процессов, изменение адгезивных свойств и рецепторной функции плазматической мембраны, способности к случайной миграции и хемотаксису, поглощению и цитотоксичности. Если учесть, что каждая из этих реакций по своей природе интегративна, то количество частных признаков, по которым можно судить о возбуждении клеток, будет огромным.
Один и тот же раздражитель способен индуцировать все или большинство признаков активации. Однако это, скорее, исключение, чем правило. Сегодня многое известно о конкретных механизмах, реализующих эффекторные свойства моно и полинуклеарных фагоцитов. Расшифрована структурная основа двигательных реакций, открыты органеллы, обеспечивающие векторную ориентацию в пространстве, изучены закономерности и кинетика образования фаголизосом, установлена природа цитотоксичности и бактерицидности, определены синтетические и секреторные потенции, обнаружены рецепторные и каталитические процессы в плазматической мембране и пр. Становится все более очевидным, что дискретные проявления клеточной реактивности обеспечиваются или по крайней мере инициируются обособленными механизмами и могут возникать независимо друг от друга. Удается подавить или усилить хемотаксис, не изменив способности к поглощению и цитотоксичности, секреция не связана с поглощением, повышение адгезивности не зависит от потребления кислорода и т. д. Известны генетические дефекты, когда выпадает одна или несколько из перечисленных функций, причем многие из них стереотипны по клинической симптоматике. Если к этому присовокупить патологию медиаторных систем, генерирующих хемоаттрактанты и опсонины, станет понятно, насколько сложным и одновременно конкретным должен быть сегодня диагноз, констатирующий нарушение фагоцитоза.
Крупным событием явилось утверждение молекулярных основ цитотоксичности (в том числе бактерицидности) и ее отношения к реактивности клеток. Стремление понять сущность реакций, приводящих к гибели поглощенных бактерий, проглядывает в большинстве работ И. И. Мечникова. Многие годы эта проблема традиционно сводилась к «перевариванию», в котором участвуют гидролитические ферменты (цитазы, по И. И. Мечникову), определяющие, как полагали, антимикробные свойства фагоцитов. Эта позиция была сильно поколеблена, после того как оказалось, что лизосомальные гидролазы обладают слабой бактерицидностью in vitro или лишены ее. В основу современных взглядов положены наблюдения, свидетельствующие об усилении окислительного метаболизма активированных фагоцитов и разобщении двух главных событий — киллинг-эффекта и деградации убитых, нежизнеспособных объектов. Прежняя терминология, в которой закреплена причинная связь между уничтожением живой мишени и переваривающей функцией клетки, оставлена. Переваривание, которое обусловлено кислыми и нейтральными гидролазами, преформированными в гранулах, является вторичным и нацелено на объекты, убитые зависимыми и независимыми от кислорода механизмами — биооксидантами, системой миелопероксидазы, катионными белками, лактоферрином, лизоцимом. Реализация цитотоксичности отражает реактивное возбуждение фагоцитирующих клеток, которые секретируют эффекторные молекулы внутрь фагосом (с образованием фаголизосом) либо наружу, атакуя внеклеточные (непоглощенные) объекты. То, что количество кислорода, поглощаемого лейкоцитами, значительно увеличивается при фагоцитозе, известно давно. Однако истинное значение этого феномена, который в современной литературе часто называют респираторным, или метаболическим, взрывом, осмыслено лишь в последние годы. В отличие от многих клеток кислородное дыхание не является системой жизнеобеспечения фагоцитов — они хорошо переносят гипоксию и выполняют ряд функций в условиях анаэробиоза. При помощи респираторного взрыва моноциты-макрофаги и нейтрофилы решают чисто эффекторные задачи, «вооружаясь» против микробов и других объектов, которые воспринимаются ими как антигомеостатические факторы. В анаэробной среде фагоциты сохраняют способность к поглощению, но резко снижают токсичность в отношении многих патогенных и условно-патогенных бактерий. Ключевой механизм сводится к активации мембранных оксидаз, катализирующих перенос электронов с НАДФН на молекулярный кислород. Это стимулирует окисление глюкозы в гексозомонофосфатном шунте, приводя к гиперпродукции перекиси водорода и свободных радикалов кислорода - биологических оксидантов с мощными цитотоксическими потенциями. Клиническое значение подобных реакций стало очевидным после того, как было обнаружено фатальное снижение противоинфекционного иммунитета у детей с врожденной патологией системы респираторного взрыва нейтрофилов. Впрочем, было бы неверно противопоставлять различные антимикробные факторы. Их эффективность во многом зависит от взаимной сбалансированности условий, в которых происходит фагоцитоз, вида микроба. Нейтрофилы, лишенные возможности использовать бактерицидные свойства активированного кислорода, тем не менее нормально убивают эпидермальный стафилококк, синегнойную палочку, зеленящий стрептококк, облигатные анаэробы. Относительная устойчивость к фагоцитозу определяется суммой признаков — поверхностными свойствами микробной клетки, наличием факторов типа лейкотоксинов и антифагинов, инактивацией биооксидантов и пр. Давно обнаружена способность некоторых бактерий тормозить образование фаголизосом. Этот механизм, который исключает контакт с цитотоксическими компонентами фагоцитов, обеспечивает длительное персистирование в макрофагах туберкулезной палочки, a в нейтрофилах — бруцелл, а также других микроорганизмов. Одну из причин видят в повышении внутриклеточного уровня циклического аденозинмонофосфата, блокирующего мобилизацию гранул и их слияние с фагосомами. Этот пример показывает, насколько глубокими могут быть взаимоотношения, которые складываются в процессе фагоцитоза.
Становление взглядов на механизмы цитотоксичности фагоцитов не подорвало мечниковской концепции о цитазах как о медиаторах, опосредующих деструктивные функции клеток. И. И. Мечников не раз подчеркивал, что, кроме разрушения микробов, фагоциты способны повреждать собственные ткани. Эти идеи получили блестящее развитие в современных работах по ферментологии лизосомальных гранул и способам их подключения к фагоцитарным реакциям. В гранулах моно- и полинуклеарных фагоцитов идентифицирован большой арсенал гидролитических ферментов (нейтральных и кислых гидролаз), для каждого из которых можно подыскать мишень во внеклеточном пространстве. Под их прицелом находятся коллагеновые и эластиновые волокна, пептидогликановый матрикс хряща, фибронектин, факторы комплемента, системы калликреин-кининов, свертывания, фибрино-лиза, иммуноглобулины, клеточные мембраны. В противовес старым представлениям сегодня сделан акцент на активном, секреторном высвобождении эффекторных молекул, которое значительно повышает эффекторную пластичность клетки, позволяя в кратчайший срок мобилизовать и рационально использовать свои возможности в физиологических ситуациях и в ходе разнообразных патологических процессов. Секретируя, фагоциты воздействуют на другие медиаторные системы и разрушают внеклеточные объекты, размер которых исключает возможность поглощения. Так, по-видимому, обстоит дело при эмфиземе легких, в реакциях на иммунные комплексы, при остром и хроническом воспалении. Кроме гидролаз и других компонентов лизосомальных гранул, активированные фагоциты выделяют пирогены, интерфероны и интерфероноподобные вещества, простагландины, тромбоксаны, биооксиданты, монокины, факторы, стимулирующие предшественники миелопоэза и пр. Лейкотриены вызывают сокращение гладких мышц и повышение сосудистой проницаемости, действуя в 100— 10000 раз сильнее, чем гистамин.
Прав был И. И. Мечников, когда говорил о широком спектре задач, решаемых фагоцитами, и многообразии их функциональных контактов («живой цепи», по И. И. Мечникову) с другими клетками и тканями. Активированные макрофаги и нейтрофилы служат одним из самых ярких примеров экстренной мобилизации эффекторных механизмов с обширной сферой приложения в масштабе не только соединительной ткани, но и всего организма.
Активаторы моноцитов-макрофагов и полинуклеаров образуются в системах комплемента, свертывания, фибринолиза, калликреин-кининов, иммуноглобулинов, выделяются лимфоцитами, фибробластами, тромбоцитами, эндотелием. Сложные взаимоотношения складываются и внутри самой фагоцитарной системы. Моноцитарный инфильтрат при воспалении формируется под влиянием хемоаттрактантов, продуцируемых нейтрофилами, которые первыми мигрируют в зону альтерации. В свою очередь моноциты и макрофаги выделяют факторы, избирательно активирующие нейтрофилы. Существенное значение имеет функциональная кооперация между однотипными фагоцитами, которая предполагает участие «аутокаталитических» механизмов, контролирующих миграционную, секреторную и другие функции активированных клеток. Липоксигеназные метаболиты арахидоновой кислоты — различные варианты гидроксиэйкозантетраеновые кислоты хемотаксичны в ничтожных дозах (особенно для нейтрофилов) и, являясь потенциальными «осколками» клеточного метаболизма унифицируют сигналы, которые обеспечивают направленную миграцию фагоцитов в очаги тканевого повреждения. Любая травма любой ткани может стать инициатором подобных реакций. В этом прослеживается один из универсальных механизмов гомеостаза — внутри популяции фагоцитов, в масштабе соединительной ткани, за ее пределами.
Из сказанного следует важный вывод. Трудно подыскать такое изменение внутренней среды организма, которое бы не фиксировалось системой фагоцитоза. Являясь мощными эффекторами, фагоциты превращаются в узел связи, своего рода стратегическую мишень, через которую трансформируются все реакции крови и соединительной ткани. Особенно показательны нейтрофилы. Обмениваясь в циркуляции каждые 5 ч, они как бы фотографируют сдвиги, которые происходят в течение этого периода, являясь своеобразным зеркалом гомеостаза. Не случайно, что индикаторные тесты, основанные на высокой реактивности полинуклеаров, давно используются в клинике и по информативности нередко превосходят другие гематологические показатели.
Много внимания уделяется молекулярным механизмам активации фагоцитов. В соответствии с общими принципами современной цитофизиологии схема фагоцитарной реакции предусматривает распознавание и рецепцию (связывание) внешнего сигнала, реактивные изменения в плазматической мембране, активацию внутриклеточных сигналов-посредников, функциональную трансформацию эффекторных органелл. Открытие стимуляторов с известной структурой привело к заключение, что их воздействие на фагоциты опосредуется через дискретные участки плазматической мембраны — специфические рецепторы, которые по молекулярной конфигурации комплементарны стимулирующему агенту. Это определяет важнейшее свойство плазматической мембраны — дифференцировать молекулярный профиль внешних раздражителей и трансформировать его в определенную форму клеточного ответа. Макрофаги и нейтрофилы рецептируют Fc-фрагмент IgG- и IgA-иммуноглобулинов, производные комплемента (СЗЬ, C3d, СЗе, С5а, С567), различные хемоаттрактанты, пектины, бактериальные гликопротеиды, фибронектин, адрен- и холинергические агенты, гистамин, кортикостероиды, эстрогены и пр. Вместе они определяют фармакологический профиль,
т. е реактивность клетки к соответствующему набору функциональных модуляторов. Выясняется, что рецепторный аппарат — весьма динамичная система. Имея определенный стартовый уровень, она меняется в зависимости от конкретной обстановки и состояния клеток. Интенсивность специфической рецепции является одной из самых интересных форм реактивности, управление которой позволит воздействовать на наиболее ранние этапы фагоцитарного процесса. Принципиально, что экспрессия разных рецепторов меняется несинхронно, дифференцируется фармакологическими агентами и приводит к неодинаковым функциональным последствиям. Пассивная по чисто внешним признакам (к примеру, хемоаттрактанты связываются разрушенными клетками) рецепция сопровождается молекулярными сдвигами в плазматической мембране, многие из которых играют ключевую роль в активации клетки. Один из постулатов современной цитофизиологии, утверждающий функциональное или даже структурное единство рецепторов и ферментов плазматической мембраны (эктоферментов), нашел отражение и в исследованиях по фагоцитозу. В составе плазматической мембраны фагоцитов обнаружены серинэстеразы, протеиназы, аденилатциклазы, оксидазы, АТФ-азы. Полагают, что они активируются при стимуляции, воспринимая изменения молекулярной топографии клеточной поверхности. Ферментология плазматической мембраны отражает стремление понять механизмы, инициирующие переключение энергии раздражения в энергию клеточного возбуждения Поиски универсального фермента-инициатора не оправдались, что, скорее, говорит о специфике различных форм клеточной реактивности, нежели отрицает саму идею эктоферментного опосредования. Не увенчались успехом и поиски универсального медиаторного звена между реакциями плазматической мембраны и эффекторными органеллами. На эту роль претендовали циклические нуклеотиды, Са2+, производные активированного кислорода. Каждый из них контролирует более или менее сложный набор клеточных функций, но в целом их эффект неуниверсален. Напротив, есть факты, которые убеждают, что внутриклеточное опосредование может быть полидетерминантным, т. е. зависит от совместного действия нескольких медиаторов. Именно такое сочетание определяет конечную форму ответа и свойственно большинству фагоцитарных реакций. По последним данным, механизмы, обеспечивающие выход на одни и те же органеллы, могут быть неодинаковы для разных стимуляторов. Глубокое развитие получили идеи И. И. Мечникова и его современников об опсонинах, сыгравшие некогда решающую роль в объединении клеточных и гуморальных концепций иммунитета. Понятие об опсонинах сформулировано в 1903 г., а усиление фагоцитоза в сывороточной среде замечено еще раньше. Однако лишь последние десятилетия ознаменовалось радикальными успехами в изучении молекулярных основ опсонической функции и ее реализации на уровне клеток-эффекторов. К семейству опсонинов обычно относят четыре группы хорошо охарактеризованных сывороточных белков — IgG, СЗ (СЗЬ), фибронектин, С-реактивный белок, однако в действительности их, очевидно, больше. Наиболее универсальными свойствами обладают СЗЬ- и IgG-антитела. Кооперируясь друг с другом, они создают мощный опсонический заслон, который традиционно считается одним из главных факторов противоинфекционного иммунитета. Функции других опсонинов, по-видимому, более ограниченны, их активность сильнее зависит от свойств фагоцитируемого объекта и типа фагоцитов. Именно неоднородность субстратов, с которыми приходится сталкиваться фагоцитирующим клеткам, следует считать первопричиной эволюционно закрепленной гетерогенности опсонических факторов.
Проблема опсонинов гораздо шире, чем противомикробная защита. В наиболее общем виде она сводится к фокусированию клеточных реакций на любых объектах экзогенного и эндогенного происхождения, которые, попадая во внутреннюю среду, вступают в противоречие с гомеостазом. Кроме уничтожения микробов, опсонизация способствует удалению продуктов тканевого распада, разрушению малигнизированных и инфицированных вирусами клеток, гибели одноклеточных и многоклеточных паразитов, элиминации антигенов, прочно связанных с базальными мембранами и другими тканевыми субстратами. Опсонические реакции, как и фагоцитоз в целом,— явление, безусловно, физиологическое. Но это тот случай, когда о значении эффекторных механизмов удается судить по их поведению в патологических ситуациях. И. И. Мечников утверждал, что действие опсонинов не ограничивается улучшением физических контактов между объектом и фагоцитирующей клеткой. По его мнению, опсонины не только меняют поверхностные свойства фагоцитируемых частиц, но и стимулируют фагоциты. Есть основания считать опсонины своеобразными фармакологическими агентами, которые стимулируют клетки независимо от поглощения по сигналу с рецепторов плазматической мембраны, вошедших в контакт с опсонизированной поверхностью. В составе Fаb-фрагмента IgG обнаружен тетрапептид — тафтсин (в честь Тафтского университета в США), который в незначительных дозах усиливает многие функции поли- и мононуклеарных фагоцитов. Справедливость идеи о стимулинах становится еще очевиднее, если изучать не изолированные системы фагоцит — опсонизированный объект, а реальные ситуации, возникающие в организме. Многочисленные наблюдения показывают, что на объекты фагоцитоза нельзя смотреть как на пассивные сорбенты опсонических факторов. Процесс опсонизации сопровождается реакциями в различных системах гуморального гомеостаза, что приводит к гиперпродукции биологически активных веществ, прямо или косвенно влияющих
на клеточную реактивность. Это отчетливо прослеживается на примере комплемента. Наработка СЗ-опсонинов связана с его каскадной активацией и сочетается с образованием С5а — одного из самых мощных эндогенных стимуляторов фагоцитоза.
Важен вопрос об отношении
фагоцитоза к приобретенному
(специфическому)
иммунитету. Классический
постулат И. И. Мечникова о переваривании
антигенного материала фагоцитами как
необходимом этапе образования антител
трансформировался в современную
концепцию о предъявлении антигенных
детерминант Т-лимфоцитам в виде комплекса
с продуктами иммунорегуляторных генов
(Ia-белками),
премированных на плазматической
мембране макрофагов. Вкупе с медиаторами
типа интерлейкинов это определяет
центральную позицию мононуклеарных
фагоцитов в механизмах формирования
приобретенного иммунитета. Для нейтрофилов
не удалось добиться большего, чем факта
слабого усиления пролиферации
В-лимфоцитов нейтральными протеиназами
нейтрофилов человека и негативного
влияния избытка нейтрофилов на
аналогичный феномен. Скорее всего,
это «пробирочные» реакции, не имеющие
серьезного приложения к регуляции
лимфоцитарных функций in
vivo.
Иначе обстоит дело
на уровне эффекторного
звена иммунных процессов. По существу,
ни одно из проявлений приобретенного
иммунитета не воспроизводится в полном
объеме без участия моноцитов-макрофагов
и (или) полинуклеаров. Об этом говорят
реакции, наведенные антителами-опсонинами,
гиперчувствительность замедленного
типа, повреждения, вызываемые
иммунными комплексами, и проч.
Зависит от МФ ответ лимфоцитов на неспецифические митогены — фитогемагглютинин, конканавалин А, перийодат, как и продукция ими лимфокинов — МИФ, МАФ, лимфотоксина и др. Предполагают, что активация Т-лимфоцитов осуществляется непосредственно свободным или связанным с МФ митогеном, а МФ выделяет фактор, трансформирующий Т-клетки. МФ, выделяя различные факторы, регулируют пролиферацию и дифференцировку незрелых костномозговых МФ, програнулоцитов, возможно, эритроидных клеток. Удалось выявить генетически обусловленные различия регулирующего влияния МФ на пролиферативную активность стволовых кроветворных клеток. Макрофаги также с помощью различных растворимых факторов усиливают пролиферацию фибробластов, эпидермальных клеток кожи, эндотелия сосудов, участвуют в созревании клеток тимуса.
Сегодня не может быть полностью принята гипотеза о центральной роли тимуса и Т-клеток в противоопухолевом надзоре, поскольку есть сведения об одинаковой частоте возникновения опухолей у бестимусных и нормальных животных, одинаковом их отторжении у иммунодефи-цитных или супрессированных животных, ограниченном числе опухолей у людей с тяжелой иммуносупрессией. По мнению исследователей, роль Т-лимфоцитов достаточно однозначна в отторжении вирусиндуцированных опухолей, но она невелика при спонтанных и индуцированных канцерогенами неоплазмах. Целый ряд доказательств свидетельствует о сложной системе естественной и приобретенной антиопухолевой защиты организма и ограниченной роли в ней Т-лимфоцитов. Об этом говорит раннее развитие естественной резистентности к опухолям на протяжении нескольких дней после рождения, подавление ее при введении веществ, угнетающих МФ, непосредственно перед трансплантацией опухоли или одновременно с ней, совпадение индуцированной стимуляции резистентности к опухолям с активированием МФ. Поэтому все большее значение в этой системе придают МФ. Оказалось, что неактивированные МФ не оказывают антиопухолевое действие, но положение меняется при активации клеток, которая может быть специфической и неспецифической. Специфическая активация возникает у клеток, взятых из иммунного организма или у интактных МФ, инкубированных с иммунными Т-лимфоцитами, с этими лимфоцитами и АГ или с продуктами реакции. МФ в этом случае активируются специфическим армирующим фактором, специфически узнают и убивают опухоль в результате цитолизиса. Неспецифическая активация обусловлена инфекционным процессом, эндотоксинами, липидом А, полинуклеотидами, иммунными комплексами иной специфичности. Активированные таким путем МФ приобретают цитостатические свойства. МФ могут армироваться специфическим к данным лимфоцитам фактором иной специфичности в отличие от фактора, индуцированного опухолевыми АГ, в таком случае они становятся неспецифически цитотоксичными по отношению к опухоли. Поэтому, если к иммунным МФ добавить специфические опухолевые клетки, а затем через некоторое время неспецифические, МФ будут специфически лизировать гомологичную мишень и неспецифически подавлять неродственную.
Таким образом, МФ в организме скорее всего проявляют одновременно специфическую и неспецифическую клеточную цитотоксичность, обнаруживая в первом случае цитолитические, а во втором — цитостатические потенции.
Активированные МФ подавляют пролиферацию нормальных и опухолевых сингенных, алло-генных и ксеногенных клеток — быстро пролиферирующих сильнее, чем медленно пролиферирующих. Однако быстро пролиферирующие опухолевые клетки полностью подавляются, тогда как нормальные клетки — лишь частично.
Механизм цитотоксичности МФ сложен. Специфический армирующий фактор с молекулярной массой 25000—50000 дальтон имеет аффинность к АГ опухоли, связывается с поверхностью МФ, секретируется коммитированными Т-лимфоцитами. Важен межклеточный контакт мишени и МФ, который постоянно возникает, причем зона контакта имеет 100—200А. Предполагают, что он может способствовать локальному слиянию и инъецированию в мишень лизосом МФ, лизирующих ее. По разным данным, киллинг может осуществляться сериновыми протеазами, возникающим под влиянием лизосомальных гидролаз СЗа-субкомпонентом комплемента, катионным белком или индукцией макрофагами в мишенях аберрантного деления, приводящего к их лизису. Вместе с тем считают, что роль простагландинов, аргиназы, перекиси водорода и интерферона не столь существенна в непосредственном цитолитическом эффекте.
Определенное значение придают изменению структуры мембран эффекторов и мишеней, так как обработка МФ фосфолипазой или лизолецитином индуцирует в них противоопухолевую цитотоксичность, что объяснили возможным образованием на мембране цитолитической структуры в результате изменения ее конформации. Подобные процессы, по-видимому, происходят при активации МФ липополисахаридами (ЛПС), в результате которой ЛПС через липид А связывается с плазматической мембраной М.Ф, изменяя ее организацию в результате формирования комплекса липид А — мембранный липид, по этой же причине, возможно, тумороцидная способность МФ резко подавлялась после их экспозиции с липопротеидами низкой плотности или холестериновыми липосомами.
В организме в силу постоянного выделения бактериальных эндотоксинов, иммунологических реакций различной специфичности, сопровождающихся освобождением лимфокинов, образованием иммунных комплексов, постоянно поддерживается популяция неспецифически активированных МФ, выполняющих надзор за спонтанно появляющимися трансформированными клетками и элиминирующими их.
МФ имеют огромное значении системы мононуклеарных фагоцитов в естественной устойчивости организма к опухолям. Поскольку подавление пролиферации макрофагами не зависит от вида и типа клеток, характеристики роста, трансформации и реактивности, это свидетельствует о наличии у МФ структур узнавания, не имеющих иммунологической специфичности. В мишенях появляются общие изменения, узнаваемые вездесущими МФ, которые поэтому являются широкими регуляторами общего клеточного гомеостаза.
За 120 лет, прошедшие со дня создания И. И. Мечниковым учения о фагоцитах, оно ушло далеко вперед. Роль МФ оказалась неизмеримо более широкой и вышла за рамки иммунологии.
Эта теория оказала глубочайшее конструктивное влияние на все развитие современной иммунологии. Именно она послужила началом изучения клеточных аспектов иммунитета. Некоторые аспекты теории, предсказанные И. И. Мечниковым, до сих пор остаются недостаточно разработанными. Очевидно, что научное наследие, оставляемое нам И. И. Мечниковым, и в будущем будет определять основные направления учения о фагоцитах.
Макрофаги перитонеального экссудата как модель фагоцитоза и нарушений фагоцитарной активности.
В организме человека фагоцитирующую функцию выполняют несколько типов клеток. Прежде всего, это те клетки, которые осуществляют защиту при каких-либо инфекциях и инвазиях – макрофаги, моноциты и нейтрофилы. В меньшей степени она представлена у эозинофилов и базофилов. Кроме того, общеизвестным является тот факт, что в различных тканях фагоцитирующую функцию берут на себя (помимо вездесущих макрофагов) специфические клеточные элементы данной ткани, например: фиброкласты, остеокласты, клетки микроглии. Также нельзя забывать о том, что к тем немаловажным клеткам, благодаря которым идет специфический иммунный ответ, относятся дендритные клетки, широко представленные в местах, являющихся барьерными в организме. И хотя их фагоцитирующая функция до конца не доказана, данные о том, что это так, есть.
Существуют различные методы изучения фагоцитирующей активности у клеток, перечисленных выше. В данном реферате будут рассмотрены методы изучения тех фагоцитов, у которых фагоцитирующая функция наиболее выражена и которые представляют исключительную важность в иммунной системе человека, а их патология носит тяжелый характер. Речь идет о макрофагах (МФ). Они хорошо поддаются изучению in vivo и in vitro, культивированию; моделирование различных процессов на этих клетках получило широкое распространение и дало хорошие результаты. Это обусловлено их крупными размерами, широчайшей распространенностью в организме, активностью метаболических процессов, протекающих в них, разнообразием функций, на них возложенных.
В качестве модели, хорошо себя зарекомендовавшей и наиболее часто используемой в опытах по изучению фагоцитов, можно рассмотреть модель перитонеальных макрофагов in vitro. Широкое распространение данная модель получила по нескольким причинам. Во-первых, она легко воспроизводится. Во-вторых, она позволяет легко регистрировать результаты исследований. В-третьих, данную модель можно получить как у лабораторных животных (крысы, мыши, морские свинки), так и у человека. В-четвертых, при проведении опытов на животных можно использовать различные линии животных (в т.ч. нокаут-животных) и животных с приобретенными (искусственно вызываемыми) дефектами иммунитета. В-пятых, при постановке опытов можно индуцировать септическое и асептическое воспаление, можно перед взятием клеток провести различные воздействия, а на модели лишь зарегистрировать результаты (т.е. сама реакция проходит in vivo).
Можно приводить также и другие причины, но ограничимся этими. Естественно, эта модель не является единственной, предложены и другие, о которых будет упомянуто ниже.
Итак, рассмотрение выбранной модели будет происходить следующим образом:
Варианты получения модели.
Возможные методы регистрации результатов исследования.
Различные варианты моделируемых процессов.
Вопросы, касающиеся практического аспекта использования результатов исследования, будут рассматриваться в каждом случае, а не будут выносится в отдельный раздел.
I.Получение модели.
Опыты проводят на белых мышах, крысах, морских свинках (в редких случаях на кроликах) различных линий (CC57/W, CBA, «WISTAR», C57BL/6), а также на нелинейных животных. Выделяют индуцированные и неиндуцированные МФ. В случае, если необходимы интактные МФ, то животным вводят интраперитонеально стерильный 10% раствор пептона или несколько мл стерильного парафинового масла, можно также использовать 2,5% раствор крахмала в физ. растворе. Обычно, через 48 часов наркотизированное животное забивают, перитонеальную полость промывают и перитонеальную жидкость отсасывают. В полученную жидкость добавляют стабилизатор (гепарин, глютатион) и антибиотики (но только не макролидового ряда!), чаще используют пенициллин, стрептомицин. Далее жидкость можно центрифугировать и последующей выдержкой, а можно сразу выдерживать в стеклянных кюветах (2 ч). Основной принцип состоит в том, что макрофаги обладают способностью прикрепляться к стеклу или пластику, тогда как другие клетки этой способностью не обладают. После экспозиции саму среду сливают (или промывают) и готовят новую, не содержащую гепарин. Полученная таким образом популяция клеток считается, что состоит на 95% из МФ. Далее клетки помещают на специальные среды (N199) с питательными веществами и антибиотиками. Сохраняться такие МФ могут не более 48-72 часов при поддержании оптимальной температуры (37 С) и ионно-осмотического баланса.
В случае, ежели необходимы активированные МФ, то их активацию проводят путем
Иммунизации животного введением различных сывороток или мощных антигенов,
Индуцированием очага септического воспаления брюшины (введение токсина в р-ре пептона, введение взвеси убитых или живых микроорганизмов).
Дальнейшие действия совпадают с уже названными.
Представляет интерес также выделение человеческих МФ. Обычно, их получают из асцитической жидкости больных с недостаточностью кровообращения III степени. Затем их осаждают центрифугированием (400g, 10 мин), замораживают при температуре жидкого азота. После размораживания их помещают в специальные чашки со средой и культивируют.
Подчас непосредственно МФ полученные из перитонеального экссудата служат лишь для регистрации опыта поставленного над животным in vivo и их культивирование носит только диагностических характер.
II.Регистрация результатов
После постановки опытов возникает резонный вопрос, а как обнаружить изменение активности МФ, как определить те изменения, повлиявшие на работу фагоцитирующих клеток. В нашей стране наиболее широко используется несколько методов.
Для исследования поглотительной фазы фагоцитоза используют различные тест-объекты. Ими могут служить кроме микробов эритроциты и различные индифферентные частицы: латекса, туши, кармина, коллоидного угля, кадмия. Поглотительную активность фагоцитов оценивают прямым визуальным подсчетом поглощенных микробов или других частиц внутри МФ, а также по числу частиц, оставшихся непоглощенными, например частиц латекса, с помощью электронного счетчика частиц, эритроцитов по концентрации гемоглобина спектрофотометрически, эмульгированных частиц масляного красного со спектрометрической регистрацией или меченных флюоресцеинизотиоцианатом микрококков с помощью флюориметра. Высокая точность и производительность характеризуют метод изучения фагоцитоза флюоресцирующих частиц латекса с помощью автоматического проточного цитофлюо-риметра. При использовании прямого визуального метода рассчитывают фагоцитарный индекс (ФИ) — процент фагоцитирующих клеток от общего числа, фагоцитарное число (ФЧ) — среднее количество частиц, захваченных одной клеткой. Отдельно учитывают результаты через 1 и 3 ч: соответственно ФИ1, ФИ3, ФЧ1 и ФЧ3 , а также коэффициент фагоцитарного числа (КФЧ): отношение ФЧ1 к ФЧ3 — показатель, характеризующий скорость фагоцитоза.
Необходимо помнить, что эффективность всех этих показателей зависит от ряда условий, таких как длительность инкубации, формы дна сосуда — круглой и конической (в конических пробирках наблюдались более высокие показатели фагоцитоза, что, видимо, обусловлено стимулирующим влиянием короткодистанционного взаимодействия), pH среды, содержания кислорода и углекислоты.
Оценка хемотаксиса лейкоцитов осуществляется двумя распространенными методами. Метод Бойдена основан на принципе прохождения лейкоцитов из одной половины камеры со взвесью клеток в другую половину с хемоатрактантом, разделенных между собой мембранным фильтром. Для изучения хемотаксиса макрофагов применяют фильтры с размером пор соответственно 5— 8 мкм. Имеющиеся разновидности метода Бойдена включают двухфильтровый и радиоизотопный варианты. Другой метод основан на хемотаксисе под слоем агарозы. В качестве хемоатрактанта чаще используют обработанную зимозаном или липополисахаридом сыворотку, казеин, фильтрат культуры Е. coli или других микроорганизмов, синтетические формилпептиды.
Движение клеток при отсутствии хемотаксического стимула дает характеристику
случайной двигательной активности (спонтанная миграция) фагоцитов.
Измерение эластичности клеток также можно осуществить в камерах Бойдена.
Адгезивные свойства фагоцитов оценивают по прилипаемости на поверхности стекла
или в колонках с бусами. Между способностью к распластыванию макрофагов, оп-
ределяемой под фазово-контрастным микроскопом, и фагоцитозом имеется
определенная корреляция
Для оценки уровня активности МФ используется полярографический метод (потребление кислорода), НСТ-тест (восстановление нитросинего тетразолия), йодирование (переход радиоактивного меченого йода в кислотонерастворимый осадок), окисление глюкозы (образование молекул 14СО>2> при окислении глюкозы-1-14С). Данные тесты основаны на том, что активация МФ сопровождается кислородзависимым метаболическим «взрывом». Классическим из данных методов стал НТС-тест. Дело в том, что активированные фагоциты способны поглощать нитросиний тетразолий (НСТ) и восстанавливать его в формазан. НСТ-тест позволяет дифференцировать активированные и интактные фагоциты, но его нельзя считать количественным, так как визуальная оценка результатов субъективна
Также для определения бактерицидной способности МФ используется хемолюминесцентный метод, предложенный сравнительно недавно. Как известно, фагоцитоз нейтрофилами и макрофагами сопровождается генерацией активных форм кислорода (О>2>-, Н>2>О>2>, ОН-), индуцирующих явление хемилюминесценции. Последняя пропорциональна интенсивности генерации фагоцитами активных форм кислорода и может служить косвенным критерием их фагоцитарной способности, тем более что образуемые продукты обладают выраженными бактерицидными свойствами. Метод анализа хемилюминесценции используется в клинике и эксперименте.
Среди методов регистрация хемилюминесценции (ХЛ) является наиболее чувствительным и информативным методом функциональной оценки фагоцитирующих клеток, но вместе с тем и одним из наиболее сложных, не столько в методическом плане, сколько в понимании природы биохимических и физических процессов, которые приводят к излучению света. Механизмы, лежащие в основе ХЛ фагоцитов, сложны и недостаточно изучены. Свечение может возникать в реакции O>2>+O>1>=2O>2>+hV, важную роль могут играть радикалы ОН-. Анализ различных ингибиторов свечения приводит к мысли, что синглетный кислород, гидроксильный радикал и перекись водорода вовлечены в процесс ХЛ.
ХЛ фагоцитирующих клеток значительно усиливается в присутствии люминола или
люцигенина.
Предложено много методов регистрации ХЛ фагоцитарных клеток, эти методы можно разделить на 2 основных класса.
/. Регистрация собственной ХЛ. Усиление собственной ХЛ фагоцитирующих клеток наблюдается при стимуляции опсонизированным зимозаном, бактериями, частицами латекса. Собственная ХЛ клеток имеет низкую интенсивность и лежит в широком спектральном диапазоне с максимумом в области 400—500 нм. Регистрация ХЛ требует высокий чувствительности прибора и достаточного количества выделения клеток (обычно не менее 106 клеток). Эритроциты, гемоглобин, сыворотка крови ингибируют ХЛ.
2. ХЛ в присутствии люминола. Свечение имеет на 2— 3 порядка большую интенсивность, чем собственная ХЛ. Усиление ХЛ наблюдается при действии зимозана, бактерий, частиц латекса, комплексов антиген — антитело, ионофора кальция, хемотаксических пептидов. ХЛ может наблюдаться в суспензии как выделенных, клеток, так и клеток в сыворотке крови.
Таким образом, хемилюминесцентный метод позволяет проводить быструю количественную оценку фагоцитарной и бактерицидной активности клеток. Он может использоваться при исследовании малых количеств биологического материала крови, или может служить как для оценки состояния клеток, так и для оценки опсонической активности сыворотки и влияния лекарственных препаратов.
Наиболее точными и быстрыми методами определения фагоцитарной активности лейкоцитов являются радиометрические. Так, поглотительную способность оценивают по уровню включения изотопа в фагоцитирующие клетки. Для этого используют меченные Сr эритроциты, радиоактивную масляную эмульсию или микробы, меченные 14С-глицином, 3Н-лейцином, 3Н-уридином, или частицы 192Ir. Иногда фагоцитоз оценивают по уменьшению метки (32Р) во внеклеточной среде.
Радиометрические методы отличаются быстротой постановки и объективностью оценки результатов. Как правило, в конце инкубации микробов с фагоцитами последние разрушают осмотическим лизисом, замораживанием — оттаиванием или дезоксихолатом натрия, затем добавляют на 30 мин при 37°С 3Н-тимидин и подсчитывают радиоактивность осажденных на фильтре бактерий. С помощью двойной метки определяют одновременно поглотительную и бактерицидную функцию лейкоцитов. Для этого предварительно метят микробы одним из изотопов (14С-фенилаланин, 14С-ацетат натрия), а затем в конце инкубации разрушают фагоциты и вносят 3Н-тимидин. Радиоактивность первоначально меченых микробов, включенных в фагоциты, будет отражать их поглотительную функцию, а радиоактивность, включенная в микробы, после разрушения фагоцитов будет характеризовать их бактерицидность. Существуют авторадиографические методы оценки завершенности фагоцитоза по включению изотопа в процессе инкубации на стеклах монослоя фагоцитов с микробами.
Одним из показателей функциональной активности макрофагов является уровень активности 5’-нуклеотидазы. Активность данного фермента определяют в суспензии не разрушенных МФ по методу Туманян и Кириличевой. Метод отличается простотой и точностью, достоверностью, достаточно часто используется.
III.Некоторые моделируемые процессы.
СНИЖЕНИЕ БАКТЕРИАЛЬНОЙ АКТИВНОСТИ ПЕРИТОНЕАЛЬНЫХ МАКРОФАГОВ МЫШЕИ В УСЛОВИЯХ СОЧЕТАННОГО ПРИМЕНЕНИЯ СТАФИЛОКОККОВОГО
ЭНТЕРОТОКСИНА ТИПА А И ЭНДОТОКСИНА
Механизмы патогенного действия стафилококковых энтеротоксинов (СЭ) изучены недостаточно. Известно, что блокада ретикулоэндотелиальной системы (РЭС) торотрастом повышает чувствительность животных к рвоте, индуцированной СЭ. Это предполагает, что функциональный статус РЭС играет важную роль в ответе организма на энтеротоксин. Данные литературы свидетельствуют и о возможности влияния СЭ на функционирование фагоцитирующих клеток. Во-первых, введение СЭ обезьянам через желудочный зонд приводит к развитию острого гастроэнтерита с экссудацией нейтрофилов, макрофагов и другими признаками воспаления. Во-вторых, важнейшим свойством СЭ является способность сенсибилизировать животных к летальному действию эндотоксинов грамотрицательных бактерий (липополисахарид — ЛПС), что ставит их в один ряд с веществами, вызывающими гиперактивацию РЭС, и также оказывающими сенсибилизирующее действие. Принимая во внимание постоянный контакт организма с условно-патогенными бактериями, а соответственно и с эндотоксинами кишечной микрофлоры, исследование макрофагальных функций, ответственных за элиминацию микроорганизмов в условиях воздействия СЭ и при сочетанием применении их с ЛПС, приобретает особую актуальность. В связи с этим, задачей опыта явилось изучение основных закономерностей изменения фагоцитарной и бактерицидной функций макрофагов под действием СЭ типа А (СЭА) и ЛПС.
I серию опытов по изучению фагоцитарной и бактерицидной активности проводили с макрофагами, полученными от мышей следующих групп: 1-я — через 2 ч после инъекции энтеротоксина, 2-я и 3-я — через 24 ч после введения СЭА и ЛПС в отдельности, 4-я — через 24 ч после введения эндотоксина на фоне СЭА. СЭА вызывал двукратное снижение общего числа клеток уже через 2 ч после инъекции; через 24 ч общее количество клеток по-прежнему оставалось пониженным. Если ЛПС у интактных мышей способствовал увеличению выхода клеток в брюшную полость, то на фоне СЭА их количество не только не возрастало под действием ЛПС, а даже достоверно уменьшалось по сравнению с контролем.
Изучение фагоцитарной и бактерицидной активности макрофагов через 2 ч после введения мышам СЭА выявило их заметное снижение по сравнению с показателями для контрольных животных. Через 24 ч после инъекции СЭА и ЛПС в отдельности наблюдалось усиление фагоцитоза. Выявленные закономерности изменения фагоцитарной функции макрофагов вполне согласуются с данными литературы, посвященными изучению клиренса угля у кроликов после введения стафилококковых знтеротоксинов. Н. Sugiyama также наблюдал бифазовые изменения фагоцитарной функции РЭС; подавление степени клиренса угля через 2ч после инъекции, сменяющееся его увеличением через сутки.
Бактерицидная активность макрофагов, полученных через 24 ч после обработки животных отдельно СЭА и ЛПС, также возрастала. В случае же совместного введения указанных токсинов функция поглощения изменялась незначительно, зато степень завершенности фагоцитоза резко снижалась. Необходимо отметить, что исследование морфологического состава клеток перитонеального экссудата мышей через 24 ч после инъекции СЭА и ЛПС не выявило существенных различий в процентном соотношении макрофагов в опытных группах животных. Поэтому можно утверждать, что выявленные изменения фагоцитарной и бактерицидной функций обусловлены влиянием исследуемых токсинов.
Для уточнения характера влияния СЭА и ЛПС на функциональную активность макрофагов следующую серию опытов провели в системе in vitro. С этой целью резидентные перитоне-альные макрофаги, полученные от интактных животных, инкубировали с токсинами в течение 24 ч. В данном случае функция поглощения изменялась в меньшей степени. Бактерицидная активность, также как и в опытах in vivo, повышалась под действием СЭА и ЛПС в отдельности. При одновременном добавлении токсинов к макрофагам бактерицидная функция увеличивалась, а в условиях, приближающихся к таковым in vivo, т. е. при добавлении ЛПС через 4 ч после СЭА, способность макрофагов убивать Staph. aureus также резко снижалась.
Таким образом, в условиях сочетанного применения СЭА и ЛПС происходит резкое снижение функции завершенного фагоцитоза макрофагов. Тот факт, что в условиях in vitro прослеживаются те же закономерности, что и в системе in vivo, предполагает, что СЭ оказывает непосредственное действие на макрофагальные функции. Учитывая, что фагоцитирующие клетки представляют собой первую линию защиты от чрезвычайно распространенных условно-патогенных микробов, снижение бактерицидных свойств в условиях синергического действия СЭ с эндотоксинами грамотрицательных бактерий в организме может привести к развитию тяжелых септических осложнений.
ОТМЕНА УСИЛИВАЮЩЕГО ФАГОЦИТОЗ ДЕЙСТВИЯ ОПСОНИНОВ С ПОМОЩЬЮ
ФРАГМЕНТОВ АНТИТЕЛ ПРОТИВ Fc-РЕЦЕПТОРОВ МАКРОФАГОВ
Один из наиболее важных и давно установленных иммунологических феноменов — усиление захвата фагоцитирующими клетками корпускулярных антигенов после их сенсибилизации IgG-антителами — оставался продолжительное время малоизученным. После выяснения принципов структурной организации молекулы IgG и обнаружения на поверхности фагоцитов и в том числе макрофагов рецепторов для Fc-участка IgG было постулировано, что опсонизирующие антитела обеспечивают усиление захвата корпускулярного антигена благодаря взаимодействию Fc-участка молекулы антитела с Fc-рецептором (FcR) макрофага. Единственным экспериментальным доказательством в пользу этого был факт отсутствия у Fab-фрагментов опсонизирующих антител способности усиливать захват корпускулярного антигена доказательства вовлечения FcR в процесс захвата опсонизированного корпускулярного антигена. Подобный экспериментальный подход нельзя рассматривать как адекватный для прямого доказательства вовлечения FcR в процесс захвата опсонизированного корпускулярного антигена. Исходя из сказанного, было решено получить прямые доказательства роли FcR в реализации механизма усиления захвата макрофагами корпускулярного антигена, сенсибилизированного IgG-антителами. Это было достигнуто при оценке влияния на указанный процесс Fab-фрагментов антител против FcR макрофагов, которые, как было установлено, препятствуют взаимодействию с макрофагами агрегированного IgG. Пре-инкубация перитонеальных макрофагов с Fab-фрагментами анти-FcR-антител полностью отменяла эффект усиления захвата макрофагами опсонизированного антигена.
В результате опыта было установлено, что Fab-фрагмент IgG из анти-FcR-сыворотки эффективно блокирует FcR мышиных макрофагов, препятствуя связыванию этими клетками гетерологичного агрегированного IgG. Эти данные хорошо согласуются с фактом блокирования FcR перитонеальных макрофагов бивалентными FаЬ-фрагментами из антиFcR. Эти данные в сочетании с результатами контрольных экспериментов, свидетельствующими об отсутствии способности блокировать FcR Fab-фрагментами IgG из антисыворотки против иммунного преципитата, указывают на возможность использования моновалентных Fab-фраг-ментов aнти-FcR-aнтитeл для изучения функции FcR макрофагов в реализации действия опсонинов. Следует подчеркнуть, что использование моновалентных фрагментов анти-FcR-антител имеет принципиальное значение при изучении функциональной роли FcR, поскольку в отличие от нерасщепленных антител или бивалентных FаЬ-фрагментов моновалентные Fab-фрагменты неспособны, связавшись с FcR, вызвать при 37 °С их латеральное перемещение по цитоплазматической мембране и последующий кэппинг.
Полученные данные свидетельствуют, что преинкубация макрофагов с Fab-фрагментами aнти-FcR-антител полностью отменяет эффект усиления фагоцитоза S.typhimurium(взятую как объект проверки эффективности фагоцитоза), обусловленный IgG-антителами кролика против этого микроорганизма. Собственно Fab-фрагменты aнти-FcR-антител не оказывают какого-либо влияния на фагоцитоз несенсибилизированных антителами бактерий. Если к макрофагам предварительно добавляли Fab-фрагменты антител кролика против иммунного преципитата, образованного яичным альбумином и IgG-антителами кролика против этого антигена, это не оказывало никакого влияния на процесс фагоцитоза сенсибилизированных антителами бактериальных клеток.
Таким образом, Fab-фрагменты анти-FcR-aнтител избирательно подавляют фагоцитоз только сенсибилизированных антителами бактерий. С учетом результатов контрольных экспериментов это служит несомненным доказательством в пользу того, что усиление захвата корпускулярного антигена после его опсонизации IgG-антителами обусловленно взаимодействием Fc-уча-стка опсонизирующих антител с FcR макрофагов.
Обращает на себя внимание тот факт, что макрофаги, предварительно обработанные Fab-фрагментами aнти-FcR-антител, менее эффективно поглощают сенсибилизированные антителами бактериальные клетки, чем несенсибилизированные. Этот результат можно объяснить, исходя из представления, что после сенсибилизации бактерий у части из них происходит стерическое экранирование участка клеточной поверхности, посредством которого микроорганизмы прикрепляются к макрофагам. Фагоцитоз таких бактериальных клеток осуществляется, по-видимому, после взаимодействия двух молекул антител или более через их Fс-участки с несколькими FcR на цитоплазматической мембране макрофага. Если указанное предположение справедливо, захват этой части сенсибилизированных бактериальных клеток будет невозможен после блокирования FcR с помощью Fab-фрагментов aнти-FcR-антител.
Полученные в настоящей работе данные открывают новые подходы для регуляции процесса иммунного фагоцитоза, что может иметь существенное значение для детального анализа роли этого процесса при различных патологических состояниях.
УСИЛЕНИЕ С ПОМОЩЬЮ ХИТОЗАНА РЕАКЦИИ КОНТАКТНОГО
ВЗАИМОДЕЙСТВИЯ МАКРОФАГА С ТИМОЦИТАМИ in vitro
Старая и многогранная проблема иммуностимуляции приобрела новое выражение в связи с поиском путей к созданию искусственных вакцин. Основные усилия в данном направлении связаны с получением конъюгатов вакцинирующих антигенных детерминант с природными или искусственно синтезированными полимерными носителями. Роль носителя в таком молекулярном комплексе должна состоять в усилении специфического ответа на избранную антигенную детерминанту.
При поиске эффективного носителя, который мог бы быть использован при конъюгации с антигеном, необходимо иметь сведения о его адъювантном эффекте и отсутствии побочных патогенетических свойств. В этой связи было обращено внимание на хитозан — гомополисахарид, выделяемый из хитина наружного скелета беспозвоночных. Молекулярная масса изучаемого вещества ~ 120000 дальтон.
Цель исследования — выяснить влияние хитозана на процесс контактного взаимодействия макрофага с тимоцитами в опытах in vitro. Обращение к данным клеточным типам не случайно. Известно, что взаимодействие макрофага с тимоцитами является дополнительным фактором трансформации недифференцированных тимусзависимых клеток в зрелые Т-лимфоциты. В связи с этим интересно определить, оказывает ли какое-либо влияние избранный гомополимер на один из самых ранних этапов становления иммунной системы — формирование функционально активной популяции зрелых Т-клеток.
Было проведено 3 серии опытов. В 1-й серии изучали характер взаимодействия сингенных тимоцитов с прилипающими клетками перитонеального экссудата, которые инкубировали непосредственно перед постановкой реакции с одной из выбранных доз хитозана в течение различного времени. Установлено, что наиболее эффективно реакция гроздеобразования проходит при 30-минутной инкубации макрофагов с хитозаном. Количество тимоцитов, группирующихся вокруг макрофага, в 2,5 раза больше по сравнению с таковым в контроле. В этой же серии опытов решался вопрос об интенсивности взаимодействия анализируемых типов клеток в зависимости от дозы, использованного для инкубации хитозана, при оптимальном времени инкубации 30 мин. Выяснено, что наибольшее усиление реакции гроздеобразования наблюдается при добавлении в культуру 50 мкг полисахарида.
Во 2-й серии опытов анализ реакции гроздеобразования проведен в условиях предварительной 30- и 60-минутной инкубации тимоцитов с разными дозами хитозана. Инкубация в течение как 30, так и 60 мин приводила к усилению контактного взаимодействия тимоцитов с макрофагами. Как и в предыдущей серии опытов, оптимальная доза, усиливавшая эффект гроздеобразования, равнялась 50 мкг.
В заключительной 3-й серии опытов проведено изучение интенсивности гроздеобразования при внесении хитозана непосредственно в реагирующую систему макрофаг—лимфоцит. Как и в 2 предыдущих сериях опытов, констатировано усиление контактного взаимодействия тимоцитов с макрофагами.
Для объяснения выявленных фактов необходимо иметь в виду, что хитозан является поликатионом. В то же время интегральный заряд как тимоцитов, так и макрофагов отрицательный. Возможно, эффект усиления контактного взаимодействия связан с электростатическими межклеточными притяжениями под влиянием хитозана, включающими 2 этапа: 1-й этап — ад-гезия положительно заряженного хитозана на макрофаге или тимоците, 2-й — непосредственное взаимодействие отрицательно заряженной клетки с клеткой-партнером, проинкубированной с поликатионом и несущей в результате этого больший положительный заряд. Подобное представление подкрепляется тем, что эффект усиления реакции гроздеобразования не связан со схемой инкубации и будет одинаков независимо от того, какой тип клеток подвергался инкубации с хитозаном.
Второй момент, который требует объяснения, — это незначительное время инкубации клеток с хитозаном, при котором обнаруживается эффект усиления контактного взаимодействия. Возможно, что отсутствие эффекта при более длительной инкубации связано с фагоцитозом высокомолекулярного полисахарида, вследствие чего происходит восстановление исходного заряда взаимодействующих клеток. Однако представленное объяснение должно быть подтверждено дополнительными экспериментальным фактами.
АКТИВАЦИЯ ФАГОЦИТАРНЫХ КЛЕТОК И КЛЕТОЧНОГО ИММУНИТЕТА
СИНТЕТИЧЕСКИМИ ПОЛИЭЛЕКТРОЛИТАМИ
Ряд перспективных неприродных полиэлектролитов, использующихся для создания синтетических вакцин, обладает многими иммуномоделирующими потенциями: они усиливают миграцию стволовых клеток, Т- и В-лимфоцитов, являются стимуляторами Т- и В-клеток, замещают хелперную функцию Т-лимфоцитов и макрофагов. Эти свойства обеспечивают сильное стимулирующее действие на реакции гуморального и клеточного иммунитета.
Вместе с тем недостаточно ясным остается вопрос об их действии на систему фагоцитарных клеток, формирование клеточного, в частности трансплантационного, иммунитета. Целью настоящей работы было изучение этих вопросов.
Методика исследований была следующей: мышам гибридам(CBAXC57BL/6) внутрибрюшинно вводили различные дозы полиэлектролитов однократно, выделяли МФ через 48 ч и анализировали их.
Введение полианиона NA-5 вызывает в макрофагах мышей активацию гликолиза. Например, в 3 экспериментах усиление гликолиза по сравнению с контрольными клетками было в 1,45, 2,35, 4,3 раза. Это очень сильная активация гликолиза в клетках, свидетельствующая о переходе их на более высокий физиологический уровень метаболизма. Значительно возрастала в клетках и интенсивность гексозомонофосфатного шунта: в 2 из 3 опытов введение полианиона сопровождалось появлением макрофагов со средней активностью окисления глюкозы, соответствующей 8,67+1,47 и 7,24+1,95 МФЕ на 10б клеток (в контроле 5,17+0,95 и 4,1 + 1,29 МФЕ на 106 клеток). Еще более сильной оказалась интенсификация цикла мочевины, различия которого по сравнению с контрольными клетками были уже порядковыми. Например, в опытах 1—3 они составляли соответственно 8,43, 11,54 и 2,06 раза.
Существенное усиление гликолиза обусловливалось также введением животным карбоцепного полиамина Н-З: в 2 из 3 опытов активация была значительной, для ЛДГ она составляла в опытных макрофагах соответственно 89,27+7,41 и 39,54±4,56 МФЕ на 106 клеток, в контрольных — 26,36+8,36 и 20,59+3,86 МФЕ на 106 клеток. Столь же выраженным было усиление активности окисления глюкозы, которое превышало его в опытных макрофагах в сравнении с контрольными в 3 экспериментах соответственно в 2,11, 1,28 и 1,41 раза.
Крайне значительной была интенсификация цикла мочевины, так как активация ключевого фермента АРГ возрастала в различных опытах в 3,65—54,6 раза..
В то же время активность поликатиона D>11-100э >была значительно менее выражена, он не влиял существенно на состояние гликолиза и гексозомонофосфатного шунта макрофагов. Однако все же активность цикла мочевины в клетках достоверно увеличивалась, хотя и менее существенно, чем под влиянием Н-3 и NA-5.
В макрофагах мышей, стимулированных полианионом NA-5, почти двукратно повышалась активность лизосомальных гидролаз, составляя в опыте 27,42+4,09 нМ Р/ч на 10б клеток и в контроле 15,04+3,66 нМ P/ч на 10б клеток. Активность КФ после введения мышам Н-3 была еще большей — 35,51+4,82 нМ P/ч на 10б клеток. Подобное усиление свидетельствует о существенном возрастании в макрофагах переваривающей способности.
У макрофагов мышей полианион NA-5 вызывал не только интенсификацию некоторых путей метаболизма, но и повышение экспрессии рецепторов к IgG, которое было нерезко выраженным, но статистически достоверным.
Дозозависимым оказался ответ клетки, выявляемый по генерации кислородных радикалов у мышей, которым вводили различные дозы полиамина Н-3. Так, если доза 0,5 мг/мышь подавляла хемилюминесценцию макрофагов при фагоцитозе, не изменяя ее при адгезии клеток на стекло, то дозы 1, 5 и 10 мг/мышь уже обусловливали существенное возрастание хемилю-минесценции при фагоцитозе частиц. При адгезии эти дозы также оказались активирующими, за исключением дозы 5 мг/мышь. Оптимальное усиление генерации активных кислородных радикалов вызывало введение животным 1 мг/мышь препарата Н-3 — в этом случае повышалась хемилюминесценция максимально и при фагоцитозе, и при адгезии. Дальнейшее увеличение дозы не сопровождалось повышением действия на клетки. Подобное наблюдение полностью подтверждается данными литературы о влиянии этого препарата на различные иммунологические параметры.
Таким образом, синтетические полиэлектролиты—полианион NA-5 и карбоцепный полиамин Н-3 вызывают активацию макрофагов, усиливая гликолиз, гексозомонофосфатный шунт, цикл мочевины, активность лизосомальных гидролаз. Препараты повышают также экспрессию на плазматической мембране макрофагов Fc>v>-peцепторов. Имеются, однако, особенности, состоящие в том, что если Н-3 вызывает усиление генерации макрофагами активных кислородных радикалов, полианион NA-5 оказывается в этом отношении неактивным. Поликатион D>11-100э >оказывает менее выраженное влияние на макрофаги, однако существенно повышает на них экс- прессию Рс>7>-рецепторов, интенсифицирует цикл мочевины.
Формирование трансплантационного иммунитета изучали у животных, которые получали однократно внутрибрюшинно по 1 мг/мышь препаратов в день трансплантации кожи.
Результаты свидетельствовали, что все 3 препарата вызывали усиление трансплантационного иммунитета, выражающееся в достоверном ускорении отторжения трансплантата у мышей, которым их вводили. Причем, как и на других моделях, в частности при анализе состояния макрофагов в условиях воздействия препаратов, активность полииона D>11-100э> уступала активности NA-5 и Н-3. Можно сделать вывод, что полиион NA-5 и карбоцепный полиамин Н3 обладают способностью усиливать клеточный Т-опосредованный иммунитет, менее активным был D>11-100э.>
АКТИВАЦИЯ МАКРОФАГОВ ПОД ВЛИЯНИЕМ СИНТЕТИЧЕСКОГО АНТИОКСИДАНТА
Сейчас общепринятой считается закономерность: активация макрофагов (МФ) сопряжена с метаболическим (окислительным) взрывом, с активацией глюкозомонофосфатного шунта (ГМФШ), с продукцией и секрецией высокоактивных нестабильных продуктов восстановления кислорода — супероксиданионов О>2>~, перекиси водорода (Н>2>О>2>), радикалов ОН~ и синглетного кислорода (О>2>) .
Образующийся при этом избыток токсичных супероксидных радикалов, а также липопереки-си, накапливающиеся в фагосомах МФ в процессе фагоцитоза, могут обусловливать окислительное повреждение клеточных мембран и связанное с этим подавление функций МФ. У МФ описана собственная система антиоксидантной защиты, включающая супероксиддисмутазу, удаляющую избыток супероксидных радикалов, а также глу-татионпероксидазу и НАДФ-зависимую глутатионредуктазу, нейтрализующие липоперекиси.
Однако при недостаточности эндогенных антиоксидантов могут возникать различные нарушения функций МФ. Было показано, что алкилзамещенные производные 3-оксипиридина (8 ОП), оказывающие умеренное антиокислительное действие, являются эффективными ингибиторами свободнорадикальных реакций и могут быть использованы для защиты от деструктивного влияния свободных радикалов.
Целью работы было изучение влияния синтетических антиоксидантов на функции МФ. Из ряда синтетических производных ОП были выбраны 2-третбутил-З-оксипиридин (ТБОП), у которого была описана способность стабилизировать мембраны эритроцитов. Исследование проводились в сравнении со стандартным активатором МФ — бактериальным липополисахарида (ЛПС из Е. coli О55).
Данные, полученные при изучении непосредственного влияния ТБОП в сопоставлении с ЛПС на перитонеальные МФ таковы: для активации ГФДГ достаточно получасовой инкубации клеток с ТБОП. Судя по показателям прироста активности фермента, эффект ТБОП аналогичен действию стандартного активатора — бактериального ЛПС. Доля распластанных МФ возрастает по сравнению с контролем уже через 2 ч инкубации с испытуемыми препаратами. На ранних сроках (2 ч) эффект ТБОП более выражен по сравнению с эффектом стандартного активатора — ЛПС. На более поздних сроках культивирования (24 ч), активирующий эффект ЛПС продолжает нарастать, в то же время доля распластанных МФ под влиянием ТБОП имеет тенденцию к понижению по сравнению с ранними сроками, но остается достоверно повышенной в сопоставлении с постепенно возрастающим контрольным уровнем.
После внутрибрюшинного введения ТБОП уже через 1 ч он вызывает отчетливое повышение количества клеток в брюшной полости за счет МФ с преимущественным накоплением крупных МФ, причем и этот эффект аналогичен действию ЛПС.
МФ, извлеченные из брюшной полости мышей через 1 ч после внутрибрюшинного введения ТБОП, отличались усиленным распластыванием по сравнению с МФ контрольных животных. Фагоцитарная активность этих же МФ была повышенной, судя по интенсивности захвата ими клеток Candida albicans. В этих условиях эксперимента ТБОП по сравнению со стандартным активатором — бактериальным ЛПС — в большей степени активирует распластывание, а фагоцитарную активность повышает в меньшей степени. В более поздние сроки после введения исследуемого препарата (1.5— 24 ч) дальнейшего нарастания количества МФ в брюшной полости и их функциональной активности не наблюдали. В отличие от этого после введения ЛПС количество МФ в брюшной полости и их функциональная активность достигали максимального уровня лишь через 24 ч.
В связи с выявленными временными различиями стимулирующих эффектов при изучении влияния препаратов на интенсивность очищения брюшной полости мышей от введенных бактерий S. typhimurium (клиренс) ЛПС вводили за 24 ч, а ТБОП — за 1 ч до заражения. Для оценки клиренса вычисляли средние разности логарифмов концентрации бактерий через 1 ч после заражения у мышей контрольных и опытных групп. Было обнаружено значительное отставание интенсивности клиренса у мышей, получивших за 4 сут до заражения по 1 мл среды с тиогликолятом, по сравнению с контролем.
На рисунке видно, что ни ТБОП, ни стандартный активатор МФ бактериальный ЛПС не влияют на интенсивность очищения брюшной полости от введенных бактерий. Однако на фоне дефекта бактерицидности МФ, индуцированного предварительным введением среды с тиогликолятом, оба препарата в равной степени достоверно повышают исходно сниженную интенсивность очищения брюшной полости мышей. Под влиянием ТБОП, как и под влиянием бактериального ЛПС, наблюдали нормализацию уровня очищения брюшной полости, т. е. коррекцию моделированного в эксперименте дефекта бактерицидности МФ.
Таким образом, у изученного синтетического антиоксиданта ТБОП выявлена способность активировать мышиные перитонеальные МФ при непосредственном воздействии in vitro. После внутрибрюшинного введения того же препарата наблюдали повышение количества МФ в брюшной полости и их функциональной активности. У мышей с предварительно индуцированным дефектом функции клиренса брюшной полости препарат способствовал восстановлению нормального уровня антибактериальной защиты. По всем изученным тестам активирующего действия на МФ синтетический антиоксидант не уступал стандартному активатору МФ — бактериальному ЛПС. При введении мышам ТБОП наблюдали более ранние проявления активации МФ по сравнению с эффектами ЛПС.
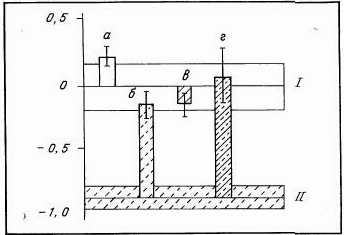
Показатели очищения брюшной полости мышей от введенных бактерий после инъекции испытуемых препаратов.
По оси ординат — средние величины разностей логарифмов концентрации бактерий в брюшной полости (М±т). I — доверительный интервал для контрольных мышей; // — доверительный интервал для мышей через 4 сут после введения среды с тиоглико-латом. а — через 1 ч после введения ТБОП; б — через 1 ч после введения ТБОП на фоне введения среды с тиогликолатом; в — через 24 ч после введения ЛПС; г — через 24 ч после введения ЛПС на фоне введения среды с тиогликолатом.
ФАГОЦИТАРНАЯ АКТИВНОСТЬ МАКРОФАГОВ ПЕРИТОНЕАЛЬНОГО ЭКССУДАТА
МЫШЕЙ ПРИ ДЕЙСТВИИ ПРЕПАРАТОВ ПЛАТИНЫ
Макрофаги способны вызывать лизис различных типов опухолевых клеток, не повреждая нормальные клетки того же гистогенеза. Нормальные «неармированные», неактивированные макрофаги осуществляют взаимодействие с опухолевыми клетками на стадии их возникновения и в период начальной стадии их развития. Вещества-цитостатики, применяемые в химиотерапии новообразований, оказывают влияние на иммунную систему макроорганизма, в частности, поражая и систему мононуклеаров. Влияние различных классов цитостатиков на функционирование макрофагального звена иммунитета достаточно глубоко изучено. Однако данных о характере влияния нового класса противоопухолевых соединений — координационных соединений платины на макрофаги в доступной литературе не встречается. Было проведено исследование — определение действия препаратов платины на фагоцитарную активность макрофагов перитонеального экссудата. В качестве препаратов были взяты Оксоплатина (цисдихлородиаминтрансдигидроксоплатина IV производства фирмы «Lachema») и циклоплатам (аминциклопептиламин-5-малатоплатина (II) отечественного производства).
В ходе проведенных исследований было установлено, что привнесении препаратов платины непосредственно в пробирки для счета при регистрации хемилюминесценции в опытах in vitro происходит незначительное увеличение высвобождения гидроксильного радикала (ОН~), супероксиданиона (О2-), синглетного кислорода ('0>2>), перекиси водорода (H>2>0>2>), что косвенно позволяет судить о стимуляции фагоцитарной активности перитонеальных макрофагов препаратами платины in vitro. Так, для циклоплатама максимальное увеличение образования активных метаболитов кислорода наблюдалось в дозе, равной 0.5 МПД (LD=23 мг/кг), и индекс хемилюминесценции составлял 3,2 по сравнению с 2,28 в контроле, тогда как добавление оксоплатины в дозе, равной 1/4 МПД, к суспензии перитонеальных макрофагов вызывало увеличение индекса хемилюменисценции с 1,69 в контрольных пробах до 2,62 в опыте.
Неоднозначные и достаточно противоречивые результаты были получены при дальнейшем исследовании влияния оксоплатины и циклоплатама на фагоцитарную функцию перитонеальных макрофагов in vivo (при введении препаратов внутрибрюшинно мышам). Введение оксоплатины и циклоплатама неиммунным мышам вызывало подавление фагоцитоза (на 1-й день после введения циклоплатама во всех дозах, на 1-й и 2-й дни после введения оксоплатины во всех дозах, с установлением стимулирующего влияния в последующие дни для обоих препаратов).
Однако введение оксоплатины и циклоплатама в тех же дозах в аналогичные сроки совместно с антигенной стимуляцией дало противоположный эффект. На 1-й день после введения оба препарата вызвали дозозависимое увеличение индекса хемилюминесценции на 2 и более порядка (индекс хемилюминесценции И>хл> для оксоплатины в дозе 1,0 МПД составил 106,9, для циклоплатама в дозе 1,0 МПД — 407,0, тогда как в контроле — 1,3—2,5). В последующие дни после введения препаратов иммунным мышам стимулирующее влияние на фагоцитарную активность перитонеальных макрофагов прослеживалось отчетливо для всех доз, но носило менее выраженный характер.
Предполагается, что при объяснении подобного явления нельзя оставить без внимания факт гетерогенности перитонеальных макрофагов и неизбежной реакции на внутрибрюшинное введение аллоантигена, выражающейся в перераспределении субпопуляций перитонеальных макрофагов в пользу так называемых воспалительных в отличие от резидентных. Не исключено появление незрелых резидентных макрофагов, также характеризующихся большей пероксидазной активностью.
Однако возможно, что при подобной постановке реакции регистрировался факт захвата и поглощения перитонеальными макрофагами частиц, коими могли быть (и явно были) не только гранулы зимозана, но и гетерологичные эритроциты барана. В пользу этого говорят данные работы, проделанной X. М. Исиной в лаборатории И. Я. Учителя , о том, что именно в 1-е сутки после иммунизации происходят максимальный захват, поглощение и разрушение макрофагами гетерологичных эритроцитов с последующей (к 48-му часу) стабилизацией процесса. Поэтому делается вывод, что 1-е сутки введения не могут рассматриваться как основополагающие при утверждении стимулирующего влияния оксоплатины и циклоплатама на фагоцитарную активность перитонеальных макрофагов, тогда как результаты последующих дней являются достоверным подтверждением подобного явления
ИЗУЧЕНИЕ ФАГОЦИТАРНОЙ АКТИВНОСТИ ПЕРИТОНЕАЛЬНЫХ МАКРОФАГОВ В
ОТНОШЕНИИ YERSINIA PESTIS С ДЕФЕКТНЫМИ И ПОЛНОЦЕННЫМИ
FRA-ГЕНАМИ
Известно, что возможность развития чумной инфекции во многом определяется исходом взаимодействия клеток возбудителя Y. pestis с фагоцитами, который зависит от степени бактерицидной активности макрофагов (МФ) и наличия антифагоцитарных факторов у микробов. К антифагоцитарным субстанциям Y. pestis относят термоиндуцируемый капсульный антиген «фракция I», антигены вирулентности V, W, I и др.. Не исключено существование неидентифицированных компонентов с той же функцией. Действие фракции I связывают с ингибицией бактерицидной активности МФ, ее участие в процессе захвата бактерий МФ отрицается. Специфическая иммунизация животных приводит к изменению МФ, которое способствует ускорению поглощения вирулентных и вакцинных штаммов Y. pestis и последующего их лизиса. В последние годы установлено, что детерминанты фракции I и VW-антигенов локализованы на плазмидах, которые, вполне вероятно, несут и другую генетическую информацию, пока неидентифицированную, но, возможно, связанную с антифагоцитарной активностью Y. pestis. Имеющиеся в литературе данные о фагоцитозе при чуме получены в опытах, в которых не идентифицировалось, связано ли нарушение синтеза исследуемых антигенов с дефектом отдельных конкретных генов или утратой соответствующей плазмиды целиком. Последнее событие может вызвать одновременно дефектность по другим, еще не исследованным антигенам. Это еще требует уточнения.
Цель работы — определение вклада фракции I в процесс взаимодействия возбудителя чумы с индуцированными МФ иммунизированных и интактных экспериментальных животных.
В опытах использовали природный вирулентный штамм Y. pestis 4 (Fra+) и изогенный штамм 4 (Fra-), у которого синтез фракции I был «выключен» встройкой элемента Tn10 в соответствующий ген плазмиды . Испытывали по 3 клона каждого штамма. Бактерии перед опытом выращивали в течение 48 ч при 28 и 37 °С на агаре LB («Difco») рН 7,2. Фагоцитоз изучали in vitro в культуре индуцированных перитонеальных МФ морских свинок и белых мышей, интактных и иммунизированных подкожно однократно в дозе 106 микробных клеток (МК) чумной вакциной. В опытах с фагоцитами нагрузка составляла 50 микробных клеток (мк) на МФ. Пробы инкубировали при 37 °С в течение 6 ч. Интенсивность фагоцитоза оценивали с помощью показателя активности фагоцитов (АФ) и индекса завершенности фагоцитоза (ИЗФ).
Все изученные бактерии, выращенные при 28 °С (28°-культуры), когда синтез фракции I находится на очень низком уровне, поглощались одинаково вне зависимости от способности их fra-генов нормально функционировать и от того, выделены испытываемые МФ от иммунизированных или интактных животных. В опытах с бактериями, выращенными при 37 °С (37°-культуры), эффективность захвата (АФ) во всех пробах была значительно ниже, чем при 28 °С.. Поскольку снижение наблюдали у штаммов как способных, так и неспособных продуцировать фракцию I, сделано предположение, что в 37°-культуре имеет место индукция синтеза или проявление функций не фракции I, а каких-то дополнительных компонентов клеточной стенки бактерий, мешающих установлению контакта бактерий и МФ. Необходима дальнейшая работа по идентификации этих компонентов.
МФ интактных белых мышей одинаково захватывали бактерии Fra+- и Fra-, МФ иммунизированных мышей несколько активнее поглощали Fra+-бактерии. МФ морских свинок независимо от того, получены они от интактных или иммунизированных животных, более активно захватывали Fra+-бактерии. Похоже, что в организме морских свинок в отношении испытанных МФ фракция I проявляет себя как неспецифический стимулятор фагоцитоза, тогда как в МФ белых мышей должна произойти специфическая перестройка, сопровождающая иммунизацию, прежде чем фракция I окажется способной слабо стимулировать захват бактерий возбудителя. Более высокие значения АФ у вакцинированных морских свинок в отношении как Fra+-, так и Fra--культур возбудителя чумы, выращенных при 37 °С, позволяют думать также о появлении у бактерий при этой температуре культивирования дополнительных факторов, которые обладают избирательной активностью именно в отношении МФ морских свинок. Еще более выраженный стимулирующий эффект этих дополнительных факторов проявляется при контакте МФ иммунизированных морских свинок с 37°-культурами, содержащими фракцию I, что позволяет предположить также и специфический элемент действия этого антигена, направленный на усиление захвата бактерий указанными фагоцитами.
Иными словами, данные экспериментов свидетельствуют, что помимо фракции I, возбудитель чумы, выращенный при 37 °С, содержит компоненты, снижающие фагоцитарную активность МФ интактных и иммунизированных животных, и компоненты, специфически способствующие захвату бактерий МФ морских свинок. Действие последних частично или полностью в присутствии фракции I нейтрализует эффект неидентифицированных негативных факторов. Фракция I способствует захвату бактерий чумы МФ и более значима для морских свинок.
В общем виде выводы по данному эксперименту можно сделать следующие: 1. Иммунизация чумной вакциной морских свинок индуцирует специфическую в отношении фракции I перестройку в макрофагах, обусловливающую усиление захвата и переваривания Fra+-Y. pestis выросших при 37 °С. Подобного не происходит у белых мышей.
2. Дефект Y. pestis по fra-генам и отсутствие фракции I обусловливают более выраженное снижение эффективности захвата бактерий, выращенных при 37 °С макрофагами морских сви-
нок, но не белых мышей и большую степень переваривания макрофагами морских свинок при всех условиях опыта, а макрофагами белых мышей — только в отношении 37°-культур.
3. Как в захвате, так и завершении фагоцитоза роль фракции более существенна в макрофагах морских свинок.
ВЛИЯНИЕ МОДИФИКАТОРОВ БИОЛОГИЧЕСКОГО ОТВЕТА ПРИРОДНОГО
ПРОИСХОЖДЕНИЯ НА ФУНКЦИОНАЛЬНУЮ АКТИВНОСТЬ МАКРОФАГОВ
(ОНКОЛОГИЧЕСКИЙ АСПЕКТ)
Несмотря на значительные успехи химиотерапии некоторых видов злокачественных новообразований, результаты применения противоопухолевых химиопрепаратов при наиболее распространенных локализациях рака остаются малоудовлетворительными. Становится все более очевидным, что одним из основных препятствий для успешной химиотерапии злокачественных опухолей является гетерогенность популяции неопластических клеток, которая выражается, в частности, в наличии в ней клонов клеток, резистентных к химиотерапевтиче-ским агентам. Более того, такая резистентность может относиться к целым классам препаратов, что может ограничивать эффективность и комплексной полихимиотерапии. Еще более осложняет положение генетическая нестабильность опухолевых клеток, которые, имея высокий уровень спонтанных мутаций, чрезвычайно легко подвергаются мутагенному воздействию химиопрепаратов и продуктов их метаболизма. Это в значительной мере усиливает гетерогенность опухолевой популяции, способствует генерации еще большего числа резистентных к химиотерапии клонов, усиливает их способность к метастизированию, рецидиву на фоне продолжающейся химиотерапии.
В конечном счете даже весьма радикальная (на 99,5 %) редукция опухолевой массы в процессе химиотерапии почти неизбежно приводит к возобновлению процесса за счет резистентных клонов — предшествовавших или возникших в процессе химиотерапии. Более того, такие клоны оказываются в далеко зашедшей стадии опухолевой прогрессии и, следовательно, более злокачественными.
В этих условиях вполне закономерными представляются поиски путей элиминации опухолевых клеток с так называемой множественной лекарственной устойчивостью с помощью других механизмов, в частности литического потенциала иммунокомпетентных клеток. Особый интерес в этом отношении представляют макрофаги. В отличие от других типов иммуноцитов их активность в меньшей степени подавляется в процессе интенсивной циторедуктивной терапии, они способны к эффективным противоопухолевым реакциям в соотношении эффектор/мишень, приближающемся к 1:1 и, инфильтрируя опухолевую строму, имеют достаточную возможность для контакта с опухолевой клеткой. Показана возможность активации цитолитического действия макрофагов с помощью различных модификаторов биологического ответа (МБО) после воздействия противоопухолевых химиопрепаратов, в то время как активность других эффекторных систем может быть существенно подавлена. Поэтому в настоящее время идет активная разработка методов адъювантной иммунотерапии с включением активаторов макрофагов. При этом предварительная оценка эффекта последних проводится in vitro и в основном по способности индуцировать цитолитическую и цитостатическую активность. По сохранению такой способности в процессе применения химиотерапевтических противоопухолевых препаратов оценивается и «совместимость» МБО с ними. Однако индукция цитотоксичности является только одной стороной активации макрофагов, под влиянием МБО происходят другие значительные изменения функциональной активности этих клеток, в частности усиливается продукция и секреция целого ряда ростовых факторов. В рамках такого подхода было изучено влияние БЦЖ и циклофосфамида на перитонеальные макрофаги мышей. Названные препараты выбраны как модельные в виду их достаточной изученности как индукторов противоопухолевой активности макрофагов in vitro и in vivo, а также достаточно широкого применения в клинической практике.
Известно, что цитотоксическая активность макрофагов in vitro достигает своего максимума к 48—72 ч культивирования, а затем быстро снижается. Была проведена оценка ростстимулирующей активности резидентных и БЦЖ-активированных макрофагов в процессе культивирования in vitro.
Установлено, что способность поддерживать рост опухолевых клеток прогрессивно снижается у резидентных макрофагов и нарастает у БЦЖ-активированных. Если в первые 3 дня прирост числа клеток на БЦЖ-активированных макрофагах достоверно ниже, чем на резидентных (что может быть объяснено цитотоксической активностью), то затем наблюдается противоположная ситуация.
Таким образом, если индуцированная БЦЖ-активация противоопухолевой активности макрофагов носит преходящий характер, то активация продукции ростовых факторов более устойчива во времени. Более того, при тестировании цитотоксической и ростстимулирующей активности макрофагов, выделяемых из перитонеальной полости мышей в различные сроки после введения БЦЖ, было выявлено, что цитотоксическая активность (цитолитическая и цитостатическая) максимальна на 10-й день. На 15-й и 20-й дни проявляется только цитолитическая, а цитостатическая активность исчезает. Ростстимулирующая активность максимальна на 15-й и 20-й дни. Следовательно, in vitro активация макрофагов БЦЖ приводит к транзиторной экспрессии противоопухолевой активности и длительной устойчивой ростстимулирующей активности.
С учетом этих данных становится понятным характер взаимодействия БЦЖ-активированных макрофагов и опухолевых клеток в процессе длительного ко-культивирования in vitro: в первые дни за счет цитотоксичности значительно снижается количество жизнеспособных клеток, одновременно цитостатические факторы тормозят их пролиферацию, но затем благодаря выделяемым ростовым факторам интенсивность пролиферации выживших опухолевых клеток значительно превышает таковую у резидентных макрофагов, в результате чего их общее количество достигает и даже превышает исходный уровень.
В ряде случаев при культивировании малых доз опухолевых клеток — до 10 на лунку (т. е. в соотношении эффектор/мишень 5000:1) — цитотоксической активности может быть достаточно для элиминации всей опухолевой популяции, однако в тех случаях, когда ростовая фракция превысит определенный порог цитотоксической активности, наблюдается интенсивный рост оставшихся опухолевых клеток. Именно это объясняет отсутствие достоверности результатов культивирования малых доз клеток, так как отклонения от средней величины отличались большей амплитудой.
Таким образом, способность макрофагов, активированных БЦЖ, контролировать рост опухолевых клеток in vitro ограничена и проявляется только в соотношениях эффектор/мишень, весьма далеких от реально возможного in vivo,— 500:1 — 5000:1. При этом противоопухолевая активность транзиторна, а опухольстимулирующая носит более длительный и устойчивый характер. Поэтому была предпринята попытка потенцировать противоопухолевую активность БЦЖ-активированных макрофагов путем воздействия на них циклофосфамидом. По данным литературы, этот противоопухолевый препарат является вполне «совместимым» с БЦЖ-агентом (т. е. стимулирует БЦЖ-индуцированную цитотоксичность) и, следовательно, может быть компонентом комбинированной химиоиммунотерапии на основе применения БЦЖ .
Циклофосфамид вводили мышам внутрибрюшинно в дозе 200 мг/кг, за 9 дней до того получивших также внутрибрюшинно 1 мг БЦЖ. На следующий день перитонеальные клетки выделяли и оценивали их цитотоксическую активность, способность поддерживать рост опухолевых клеток в субоптимальных концентрациях и влиять на рост автономно растущей опухолевой популяции в условиях ко-культивирования. Оказалось, что циклофосфамид самостоятельно индуцировал существенную цитолитическую и цитостатиче-скую активности, кроме того, достоверно усиливал БЦЖ-индуцированную цитотоксичность.. При этом в присутствии макрофагов, активированных комбинацией БЦЖ с циклофосфамидом, уровень пролиферации опухолевых клеток в зависимых от ростовых факторов концентрациях (102 клеток на лунку) был 2,9'104±3,25-103 и значительно превышал таковой при культивировании опухолевых клеток на БЦЖ-активированных макрофагах — 3,7-103±1,4-102, практически не отличаясь от их роста в присутствии нестимулированных макрофагов — 3,2-104±4,82-103.
Таким образом, несмотря на весьма высокий уровень цитотоксичности макрофагов, индуцированный их обработкой вслед за БЦЖ еще и циклофосфамидом, такие макрофаги теряли способность даже к ограниченному контролю ко-куль-тивируемой с ними популяции опухолевых клеток.
С учетом весьма значительной в этой серии экспериментов потери клеток под влиянием факторов цитотоксичности (в отдельных опытах цитолитическая активность достигала 70 %) и прироста клеток, сравнимого с таковым после ко-культивирования на резидентных макрофагах, можно считать, что совместное применение БЦЖ и циклофосфамида оказывает аддитивное действие на продукцию ростовых факторов макрофагами.
Таким образом, имеющиеся в литературе данные о совместимости БЦЖ и циклофосфамида, будучи совершенно справедливыми в отношении противоопухолевой активности активированных макрофагов, не отражают возможного конечного результата такого совмещения, явно нежелательного с клинической точки зрения. Следует отметить, что характер ответа макрофагов на активацию МБО in vivo имеет сходство с таковым in vitro. Как показано еще в первых работах по применению БЦЖ, иммунотерапия этим препаратом эффективна только в течение короткого срока, а затем происходит стимуляция опухолевого процесса, причем противоопухолевую активность макрофагов, достаточно быстро угасающую как in vitro, так и in vivo, как правило, не удается восстановить повторными введениями препарата, ее вызвавшего, и в случае успеха такая реактивация кратковременна.
Данные литературы достаточно однозначно указывают на отсутствие корреляции между цитотоксической активностью БЦЖ-активированных макрофагов in vitro и их влиянием на опухолевые клетки in.vivo. Если исходить из представленных нами данных, это становится вполне объяснимым: уничтожение in vitro даже большинства опухолевых клеток при последующем стимулировании роста оставшихся приводит к явному нивелированию эффекта цитолитического действия, особенно если учесть его относительную кратковременность по сравнению с ростстимулирующим действием. Кроме того, выявляемая in vitro цито-статистическая активность зависит от таких факторов, как аргиназа, истощение культуральной среды ввиду повышенного метаболизма активированных макрофагов, продукция токсических радикалов, атомарного кислорода и др. В условиях in vivo эти эффекты могут не проявляться в связи с притоком аргинина, других питательных веществ к клеткам, наличием антагонистов радикалов и т. д.
Как известно, при опухолевом процессе макрофаги способствуют развитию опухоли на «органном» уровне путем улучшения микроокружения (имеется в виду стимуляция ангиогенеза, формирование стромы опухоли, элиминация продуктов распада опухолевых клеток). Продуцируемые макрофагами иммуносупрессивные факторы, в частности простагландин Е2, способны инактивировать другие иммунологические механизмы рези-стентности к опухолевому росту. Такие факторы, как интерлейкин-1 (ИЛ-1), фактор некроза опухоли (ФНО), выделяемые активированными макрофагами, способны подавлять пролиферацию большей части известных линий опухолевых клеток, однако они являются и стимуляторами роста некоторых из них .
Учитывая высокую степень гетерогенности опухолевой популяции и усиление роста таковой под влиянием продуктов активированных макрофагов, нельзя исключить появления устойчивых и даже зависимых от ФНО и ИЛ-2 клонов. И если в относительно непродолжительных до времени сроках взаимодействия макрофагов и опухолевых клеток в условиях экспериментальных моделей такие эффекты не проявятся, то в реальных условиях вероятностью такого отбора пренебрегать нельзя. Это особенно актуально, если учитывать, что макрофаги практически неизбежно вовлекаются в реализацию любого иммунотерапевтического воздействия, поэтому продемонстрированные здесь негативные последствия их активации могут сказаться и на эффективности всей программы иммунотерапии, направленной изначально на другие эффекторы иммунной системы.
Таким образом, продукция макрофагами факторов, стимулирующих рост опухолевых клеток, является весьма существенным компонентом ответа этих клеток на МБО. Соответственно при оценке и отборе потенциальных МБО необходимо оценивать не только их способность к индукции противоопухолевых реакций, но и возможность экспрессии побочной ростстимулирующей активности. Только углубленное изучение этого вопроса., направленного на выявление факторов, стимулирующих рост опухолевых клеток, путей их биосинтеза в макрофагах и их регуляцию, может лечь в основу разработки методов селективного подавления нежелательных в онкологической ситуации ростстимулирующих свойств активированных МБО макрофагов при одновременной сохранности и активации их противоопухолевой активности.
ПЕРИТОНЕАЛЬНЫЕ МАКРОФАГИ КАК МОДЕЛЬ ДЛЯ ИЗУЧЕНИЯ АТЕРОГЕННОГО
ПОТЕНЦИАЛА СЫВОРОТКИ КРОВИ
Накопление липидов в гладкомышечных клетках (ГМК) и макрофагах интимы аорты является характерной чертой атеросклероза человека и экспериментальных животных. Было показано, что сыворотки крови больных ишемической болезнью сердца (ИБС) с ангиографи-чески подтвержденным коронарным атеросклерозом в отличие от сывороток крови здоровых лиц обладают способностью вызывать накопление липидов в культивируемых клетках интимы аорты человека. Это свойство было названо атерогенностью, поскольку накопление липидов сопровождалось другими атеросклеротическими проявлениями на клеточном уровне — усилением пролиферативной активности и синтеза внеклеточного матрикса. Однако связь между атерогенностью и атеросклерозом окончательно не выяснена.
Исследования по этой проблеме основаны на первичном культивировании субэндотелиальных клеток интимы аорты человека. Сложность работы обусловлена необходимостью постоянного обеспечения стерильным аутопсийным материалом, а также высокой стоимостью выделения и культивирования клеток.
Ранее было показано, что способностью аккумулировать внутриклеточно холестерин при культивировании с атерогенной сывороткой обладают ГМК аорты человека и мононуклеарные клетки периферической крови. Эти данные позволяют считать, что для определения атеро-генности сыворотки крови могут быть использованы не только субэндотелиальные клетки интимы аорты.
Целью работы было определение возможности использования легкодоступных перитонеальных макрофагов для определения атерогенности сыворотки крови.
Кровь для исследований бы взята у больных ИБС, подтвержденной при коронарной ангиографии, и здоровых доноров. ГМК были выделены из аорты мужчин, взятой в асептических условиях спустя 24 ч после внезапной смерти, ступившей от инфаркта миокарда. Человеческие перитонеальные макрофаги были выделены из асцитической жидкости больных недостаточностью кровообращения. Мышиные перитонеальные МФ получены от нестимулированных мышей.
Влияние сыворотки крови здоровых доноров и больных ИБС на уровень холестерина в клетках
|
Тип клеток |
Контроль, мкг |
Уровень холестерина в клетках, % контрольных величин |
|
на 1 мг белка |
здоровые доноры |
больные ИБС |
ГМК:
интимы аорты
59±5 110±7 370±43
67±4 118±13 179±10
32±3 95±8 264±31
44±4 108±3 284±28
медии аортыМышиные макрофаги
Человеческие макрофаги
В таблице приведено содержание общего холестерина в ГМК интимы и медии аорты человека, в перитонеальных мышиных и человеческих макрофагах, инкубированных 24 ч в присутствии 20 % сывороток крови здоровых доноров и больных ИБС. Видно, что инкубация клеток в 20 % сыворотке крови здоровых доноров не приводит к статистически значимому повышению уровня холестерина в клетках, а инкубация в 20 % сыворотке крови больных ИБС вызывает достоверное повышение содержания внутриклеточного холестерина как в ГМК, так и в перитонеальных макрофагах. Таким образом, так же, как ГМК интимы и медии аорты, перитонеальные макрофаги мышей и человека могут быть использованы для оценки атерогенного потенциала сыворотки крови. Кроме того, накопление холестерина в макрофагах происходит в 1,5—2,5 раза быстрее, чем в ГМК. Оба эти факта, а также более легкий способ получения культуры макрофагов позволяют считать, что эти клетки могут быть использованы для оценки атерогенного потенциала сыворотки крови значительно чаще, чем ГМК.
При культивировании мышиных, человеческих макрофагов и ГМК с 10 атерогенными и 10 неатерогенными сыворотками крови установлена прямая корреляционная связь между аккумуляцией холестерина в макрофагах и в ГМК интимы аорты. Кроме того, аккумуляция холестерина в человеческих и мышиных макрофагах находится в прямой зависимости от концентрации сыворотки (рис 1) и времени культивирования (рис 1А). При культивировании макрофагов с сывороткой здоровых лиц такого эффекта не выявлено.
Во время инкубации человеческих и мышиных макрофагов с сыворотками крови больных ИБС выявлено повышение содержания в клетке свободного холестерина и триглицеридов в 1,5— 2 раза, эфиров холестерина в 2,5—3 раза. При этом. уровень фосфолипидов не изменялся.
В группе здоровых доноров только у 22 (28 %) из 80 человек сыворотки крови вызывали аккумуляцию холестерина в культуре клеток. В группе больных ИБС у 83 % человек сыворотки крови вызывали достоверное повышение уровня общего холестерина в макрофагах, т. е. обладали атерогенными свойствами.
Не выявлено корреляции между атерогенностью крови больных ИБС и содержанием в сыворотке общего холестерина, триглицеридов, апо-А1, апо-В и ХС ЛПВП.
Полученные данные свидетельствуют о наличии прямой корреляции между аккумуляцией липидов в ГМК и макрофагах, а также о том, что способность выявлять атерогенность сыворотки крови. При использовании перитонеальных макрофагов не отличается от данных, полученных ранее при использовании ГМК интимы аорты. Во время культивирования перитонеальные макрофаги аккумулируют свободный и эстерифицированный холестерин, триглицериды, т. е. те же липиды, которые при инкубации включают в себя ГМК интимы аорты. Использование ГМК затруднено из-за сложностей получения асептического материала в достаточном для работы объеме. Человеческие и особенно мышиные макрофаги лишены этих недостатков. Эти факты доказывают, что культура перитонеальных макрофагов вместе с культурой ГМК интимы аорты может быть использована как тест-система для оценки атерогенного потенциала сыворотки крови.
Макрофаги, возможно, являются одним из главных клеточных акцепторов липидов в сосудистой стенке. Давно уже было показано наличие макрофагов, насыщенных липидами, в атеросклеротических бляшках. Экспериментальные работы, в которых использовалась культура клеток, выявили способность макрофагов интенсивно накапливать эфиры холестерина при инкубации с химически модифицированными липопротеидами.
Использование культуры макрофагов позволит продолжить изучение природы атерогенности сыворотки крови больных ИБС и ее роли в патогенезе атеросклероза.
Рис. 1 Связь между накоплением холестерина в клетках и концентрацией сыворотки
По оси абсцисс — концентрация сыворотки (в %); по оси ординат — содержание холестерина (в мкг на 1 мг белка) в человеческих (а) и мышиных (б) макрофагах. /, 2 — культивирование в сыворотках здоровыхдоноров,3,4— больных ИБС.
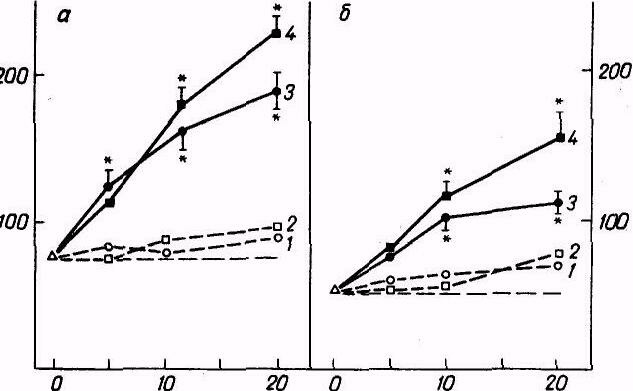
Рис.1А Связь между накоплением холестерина в клетках и временем инкубации.
По оси абсцисс—время инкубации (в ч). Остальные обозначения те же, что и на рис 1
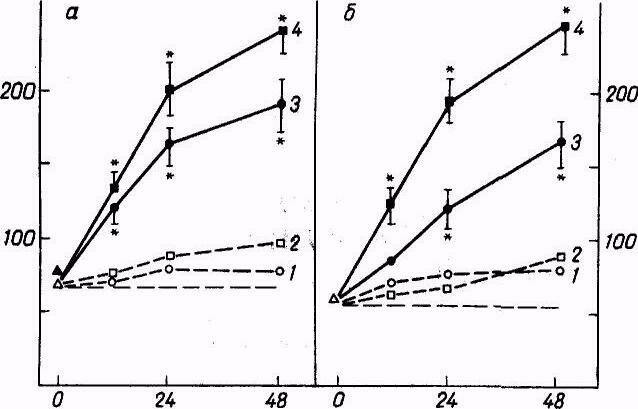
ВЛИЯНИЕ ГАМК, ГОМК И ГЛУТАМИНОВОИ КИСЛОТЫ НА ФУНКЦИОНАЛЬНУЮ
АКТИВНОСТЬ ФАГОЦИТОВ
За последние годы проводится интенсивное изучение роли нейроактивных аминокислот в иммунологических процессах. Это связано с широким использованием данных аминокислот в неврологической и психиатрической практике, однако одновременно с их прямыми эффектами получены данные об их влиянии на иммунологические процессы. Получены данные о влиянии гамма-аминомасляной (ГАМК), гамма-оксимасляной (ГОМК) и глутаминовой кислот на количество антителообразующих, розеткообразующих клеток и другие показатели иммунитета. Опыты проводились в двух сериях: в I серии на интактных мышах изучалось влияние нейроактивных аминокислот на функциональное состояние МФ и нейтрофилов. Во II серии влияние тех же веществ на состояние фагоцитов изучалось на фоне предварительной иммунизации животных. Иммунизацию проводили 5% взвесью эритроцитов барана в количестве 1 мл на 3-й день после введения препарата. Контрольную группу составляли интактные мыши, получавшие физиологический раствор (контроль I), и животные, получавшие физиологический раствор и иммунизированные эритроциты барана (контроль II).
Введение интактным животным (I серия) нейроактивных аминокислот сопровождалось существенными сдвигами активности кислой фосфатазы в МФ и нейтрофильных лейкоцитах. Необходимо отметить, что изменение активности в изученных группах носило разнонаправленный характер. Так, в условиях введения ГАМК активность фермента в МФ и лейкоцитах крови повышалась по сравнению с контрольной группой в 1,7 раза, при введении же ГОМК происходило достоверное снижение активности кислой фосфатазы лишь в МФ. Показатели активности фермента при введении интактным мышам глутаминовой кислоты не отличались от таковых контрольной группы.
Изучение влияния биологически активных веществ на фоне предварительной иммунизации эритроцитами барана во всех опытных группах II серии выявило существенное понижение активности кислой фосфотазы в нейтрофильных лейкоцитах. Относительно низкие показатели активности в изучаемых клетках крови были зарегистрированы при введении ГОМК и глутаминовой кислоты, которые были ниже соответственно в 2 и 1,4 раза контрольного уровня. Активность изучаемого фермента в МФ опытных групп II серии не отличалась от таковой в контроле, за исключением группы, в которой на фоне иммунизации вводили ГОМК, где содержание кислой фосфатазы понижалось почти в 2 раза.
В серии проведенных цитологических исследований было установлено, что нейроактивные аминокислоты в испытуемых дозах не вызывают в клетках фагоцитарного ряда дистрофических и деструктивных изменений. Так, ни в одной изучаемой группе I серии не было выявлено признаков распада, фрагментации и лизиса цитоплазмической и ядерной мембран, пикноза и рексиса ядер. В то же время в группах, где вводили ГАМК и ГОМК, ядра моноцитов выглядели гипертрофированными и гипохромными, цитоплазма—умеренно вакуолизированной. Не исключено, что нейроактивные аминокислоты в биологически допустимых дозах не оказывают цитотоксического влияния на моноциты и нейтрофильные лейкоциты в крови интактных мышей. Однако существенные сдвиги в активности основного маркёра лизосом—кислой фосфатазы наводят на мысль, что изучаемые биологически активные вещества, не оказывая непосредственного токсического действия на клеточную мембрану, вызывают все-таки дестабилизацию внутриклеточных мембранных структур, и, в первую очередь, лизосомальных. Принимая во внимание то обстоятельство, что активность лизосомальных ферментов во многом зависит от функционального состояния мембран, а точнее—от степени её проницаемости, была проведена специальная серия экспериментов по изучению проницаемости лизосомальных мембран при помощи прижизненного флюорохромирования фагоцитов акридиновым оранжевым (АО). Результаты исследований с применением АО показали, что изменение тинкториальных свойств клеток-мишеней наблюдалось при введении в организм мышей всех исследуемых нейроактивиых аминокислот. При введении ГАМК, ГОМК и глутаминовой кислоты период полураспада (Т >1/2>) заметно сокращается, а при экспозиции Т '/а, в течение которого происходит поэтапное изменение тинкторнальных свойств составных компонентов клеток (цитоплазма, ядро, участки локализации лизосом), количество фагоцитов с зеленой флюоресценцией ядер в опытных группах I серии достоверно снижалось. Аналогичная направленность четко прослеживалась во всех опытных группах II серии.
Резюмируя вышеизложенное, можно заключить, что нейроактивные аминокислоты оказывают существенное влияние на активность кислой фосфатазы в фагоцитах. Особо следует отметить, что полученный эффект в группах введения ГАМК и ГОМК был диаметрально противоположным: этот факт прослеживается наиболее четко в тех случаях, когда в качестве объекта изучения активности кислой фосфатазы выступали МФ. Введение же ГАМК и глутаминовой кислоты на фоне предварительной иммунизации не отражалось на активности кислой фосфатазы в МФ, в то время как в нейтрофильных лейкоцитах все изучаемые нейроактивные аминокислоты вызывали достоверное понижение активности фермента.
Эксперименты с применением в качестве индикатора проницаемости лизосомальных мембран АО показали, что нейроактнвные аминокислоты, не обладая цитодеструктивным действием па клетки-мишени, вызывают дестабилизацию лизосомальных мембран, направленную в сторону увеличения их проницаемости. Сопоставление показателей проницаемости лизосомальных мембран животных I и II серий показывает, что для повышения проницаемости под влиянием нейроактивных аминокислот формирование гуморального иммунитета является условием необязательным.
Таким образом, нейроактивные аминокислоты оказывают избирательное влияние на морфофункциональное состояние лизосомального аппарата фагоцитов. Это подтверждается полученными данными о проницаемости лизосомальных мембран в отношении токсического красителя и количественного определения активности основного маркера лизосом – кислой фосфотазы.
IV. Заключение
Приведенная лишь малая часть экспериментов по моделированию различных воздействий, говорит в пользу того, что процессы нарушений или изменения фагоцитирующей активности очень удобно изучать путем постановки опытов на перитонеальных МФ. Можно сказать, что данная модель давно стала базовой для проверки разнообразных химических и фармакологических препаратов в качестве агентов, воздействующих на клеточное звено иммунитета; для изучения процессов взаимодействия инфекционных агентов с фагоцитами. Также она используется для изучения не только фагоцитарной функции – но и наоборот, различных процессов, связанных с самими инфекционными агентами. Можно упомянуть, что проводятся исследования по изучению фагоцитов перитонеального экссудата у генетически диабетических мышей, у генетически иммунодепрессивных крыс и т.д.
Конечно те данные, которые получают исследователи достаточно относительны, ведь несмотря на высокое качество проводимых опытов, они все же проводятся in vitro, что неизменно накладывает свой отпечаток – клетки здесь лишены той гаммы цитокиновых воздействий, которая присутствует в живом организме, они не взаимодействуют со стромальными компонентами и другими клетками. Достоинством метода является его простота, доступность и, что немаловажно, наглядность, а также высокая скорость получения результатов и широкие возможности по постановке «регистрационных» реакций. Однако, как видно, этим в современной патологии мы не можем ограничиться. Поэтому в современных научных и клинических исследованиях применяют и другие методы и модели. О некоторых из них кратко будет рассказано ниже.
НЕКОТОРЫЕ ДРУГИЕ МОДЕЛИ ИЗУЧЕНИЯ ФАГОЦИТОЗА.
Одна из самых реалистичных моделей фагоцитоза – это модель in vivo. Данная модель позволяет получать достоверную информацию и моделировать процессы с учетом влияния внутренней среды. Однако она более сложна в реализации и порой не дает возможности работать с организмом человека. Модель in vivo существует в двух вариантах
Модель на фагоцитах сыворотки крови. Конечно, в этом случае объектом нашего внимания будут не МФ, а моноциты крови и нейтрофильные лейкоциты. Модель позволяет изучать влияние на состояние фагоцитов и фагоцитарной функции общих, системных процессов, таких как гипоксия, гипобария, радиация и проч. Также путем внутрисосудистого введения различных веществ можно получить данных об их воздействии. Изучается влияние и внутренних системных активных веществ: гистамина, простогландинов
Модель фагоцитоза в очаге хронического воспаления. Одна из самых «клинических» моделей. Здесь объектом внимания являются как тканевые МФ, так и эпителиоидные клетки, гигантские многоядерные клетки, клетки инородных тел и др. Большое внимание уделяется явлению незавершенного фагоцитоза, процессам подавления миграции, снижения кислород-зависимой цитотоксичности.
Очень оригинальная модель была разработана в лаборатории иммунологии Института акушерства и гинекологии им. Отта РАМН (авторы: О.А.Павлов,С.А.Сельков,А.В.Селютин, Е.Е.Андреева и др.). Они изучали роль плацентарных МФ в проблемах родовой деятельности и, особенно – в проблеме невынашивания беременности. Макрофагам фетоплацсптарного комплекса (МФК), по мнению исследователей, до недавнего времени уделялось неоправданно мало внимания. Лишь в последние годы появился ряд публикаций, посвященных этим клеткам. Многие функции МФК только предстоит выяснить, однако уже сейчас становится очевидным, что среди популяций различных клеток, составляющих ткань плаценты, МФК являются наиболее вероятными кандидатами на роль регуляторного (а, возможно, и ключевого) звена в ряде репродуктивных процессов. С точки зрения патогенеза развития родовой деятельности при преждевременных и срочных родах МФК представляют особый интерес в силу особенностей ряда свойств и положения этих клеток.
Плацентарные макрофаги (клетки Кащенко—Гофбауэра) принадлежат к системе мононуклеарпых фагоцитов, имеющих единое происхождение с обычными МФ. Эти клетки в значительном количестве содержатся в ткани плаценты и составляют подавляющее большинство ее иммунокомпетентных клеток. По данным некоторых авторов, макрофаги составляют до 40% популяции нетрофобластных клеток ворсин хориона.
Традиционно считалось, что прерогативой мононуклеарных фагоцитов в фетоплацентарном комплексе является удаление клеточного детрита, защита от микроорганизмов, участие в системе защиты плода от местного материнского иммунного ответа. Они также участвуют в реализации и модуляции иммунного ответа и представляют первую линию защиты против инфекции, обеспечивая неспецифические иммунные реакции и продуцируя регуляторные сигналы для специфических. Таким образом, уникальность положения МФК состоит в том, что, с одной стороны, они, как эффекторные клетки, вступают в непосредственный контакт с инфекционными агентами и иными продуктами, проникающими в организм хозяина (в данном случае в единую систему "мать—плацента—плод"), а, с другой стороны, будучи активированными в результате этого контакта, способны продуцировать множество растворимых сигнальных молекул, влияющих на функции близлежащих клеток.
В число этих факторов входят и цитокины, содержание которых меняется с началом спонтанной родовой деятельности — ФНОа, ИЛ-1, ИЛ-6 и ИЛ-8. Предполагается, что эти цитокины, продуцируемые макрофагами, стимулируют синтез ПГ и тем самым инициируют сократительную активность матки. Более того, показано, что макрофаги могут самостоятельно продуцировать ПГЕ2 ПГ2а, которые непосредственно воздействуют на миометрий. Макрофаги также способны к секреции трансформирующего ростового фактора-β (ТРФ-β), который играет важную роль в эмбриональном морфогенезе и влияет на функции клеток трофобласта и эндометрия. Недавно в экспериментах in vitro получены убедительные доказательства участия клеток плаценты в процессе родов. Авторы продемонстрировали связанное с родовой деятельностью увеличение секреции ФНО-α макрофагами, взятых у женщин в результате спонтанных родов или индуцированных родов путем искусственно родоразрешения, а также ИЛ-1 и ИЛ-6 эндотелиальными клетками плаценты. Все эти данные свидетельствуют о вкладе клеток плаценты, в том числе макрофагов, в повышение уровня цитокинов в пределах фетоплацентарного комплекса при спонтанной родовой деятельности. Предположения о возможной роли ПМ в преждевременном и срочном родоразрешении заставляют взглянуть на эту клеточную популяцию как на важнейший элемент, оказывающий влияние на гестационные процессы. В этой связи активация ПМ может рассматриваться как событие, ведущее к преждевременным или срочным родам. Установлена положительная корреляция между повышением уровня некоторых цитокинов (туморнекротизирующий фактор-α, интерлейкин-1 и -6), которые секретируются в том числе активированными макрофагами, и наступлением преждевременных и срочных родов. Попытались выявить связь между наличием родовой деятельности на разных сроках беременности и степенью активации ПМ in vitro, о которой судили по уровню экспрессии антигенов МНС II и фагоцитарной активности клеток, опосредованной Fс- и СЗ-рецепторами. при наличии родовой деятельности. Увеличивалось количество клеток, экспрессирующих МНС II и СКЗ и обладающих способностью к фагоцитозу. На более поздних сроках беременности не удалось выявить достоверного увеличения активности ПМ при родовой деятельности. Согласно данным снижение уровня фагоцитоза происходило за счет снижения активности отдельной клетки, в то время как количество фагоцитов существенно не изменялось. Для того, чтобы выяснить, отражает ли наблюдаемая тенденция существующую закономерность, или является следствием неоднородности исследуемого материала, необходимо исследовать дополнительное количество плацент.
Полученные результаты свидетельствуют о перспективности применения обогащенных культур ПМ для изучения ряда аспектов иммунологиии репродукции, в частности для выяснения клеточных механизмов, лежащих в основе нормальных и преждевременных родов.
Литература.
Д.И. Маянский, Клетка Купфера и система мононуклеарных фагоцитов, Москва, 1983
Методы изучения in vitro клеточного иммунитета, под ред. Блума и Глэйда, Москва,1974
И.И.Мечников, Лекции по сравнительной патологии воспаления, Москва, 1947
Терапевтический архив, 1990, Т62, N11
Вопросы медицинской химии, 1988, Т34, Вып.1
Лабораторное дело,1984, N5
Лабораторное дело,1985, N1
Лабораторное дело,1992, N2
Лабораторное дело,1989, N4
Иммунология, 1983, N1
Иммунология, 1983, N2
Иммунология, 1987, N6
Иммунология, 1989, N4
Иммунология, 1999, N1
Бюллетень экспериментальной биологии и медицины, 1988, N1
Бюллетень экспериментальной биологии и медицины, 1989,N11
Бюллетень экспериментальной биологии и медицины, 1990,N1
Бюллетень экспериментальной биологии и медицины,2000, Т129, N6
Бюллетень экспериментальной биологии и медицины,1999, Т127, N4
Журнал микробиологии эпидемиологии и иммунобиологии 1990,N5
Архив патологии, 1994, Т56, N1
Медицинская иммунология, 2001, Т3, N3
Экспериментальная и клиническая медицина, 1989, Т29, N3
Экспериментальная и клиническая медицина, 1989, Т29, N6
Цитология, 1992, Т34,N7
Санкт-Петербургский государственный университет
им. акад. И.П.Павлова
КАФЕДРА ПАТОФИЗИОЛОГИИ
РЕФЕРАТ
Макрофаги перитонеального экссудата как модель
фагоцитоза и нарушений фагоцитарной активности
Выполнил
студент Карпов С.А.
305 группы лечебного факультета
Преподаватель Степанян М.Л.
Санкт-Петербург
2002