Костистые рыбы
Реферат на тему:
Костистые рыбы
2009
Костистые рыбы произошли от костных ганоидов; ископаемые формы известны с триаса. Длина тела от 1 см до 5 м. Тело покрыто циклоидной или ктеноидной чешуёй либо костными пластинками. У большинства костистых рыб имеется луковица аорты. Некоторые костистые (например, каменные окуни) – гермафродиты.
Среди костистых рыб известны как хищники, так и растительноядные формы. Есть и паразиты других рыб. Костистые рыбы живут во всех водоёмах, где только могут жить рыбы; на их долю приходится 98 % общемирового улова (десятки миллионов тонн в год). К костистым рыбам относятся такие ценные промысловые породы, как тунец, палтус, лосось, сельдь, щука и другие. Обычные для наших речек караси и лещи – тоже костистые рыбы.
В настоящее время костистые рыбы достигли своего расцвета. Количество видов превышает 25 000 (это больше, чем количество видов всех остальных позвоночных, вместе взятых). Надотряд включает в себя 30–7 отрядов: тарпонообразные, конорылообразные, сельдеобразные, лососеобразные, миктофообразные, араванообразные, клюворылообразные, угреобразные, мешкоротообразные, спиношипообразные, карпообразные, сомообразные, карпозубообразные, сарганообразные, атеринообразные, перкопсообразные, трескообразные, бериксообразные, китовидкообразные, солнечникообразные, опахообразные, колюшкообразные, кефалеобразные, слитножаберникообразные, окунеобразные, скорпенообразные, камбалообразные, иглобрюхообразные, пегасообразные, батрахообразные, присоскообразные, удильщикообразные.
Жизнедеятельность костистых рыб приводит к сдвигу кислотности мирового океана из-за выделения больших количеств карбоната кальция, которое растет за счет глобального потепления, говорится в статье, опубликованной группой американских и британских ученых в журнале Science.
Рост кислотности мирового океана, связанный с увеличением количества поглощаемого водой углекислого газа, вызывает тревогу у экологов. Этот процесс ставит под удар многие морские экосистемы, существа, использующие кальций - в частности, кораллы, планктон, моллюски, некоторые водоросли. Из-за роста кислотности уменьшается толщина их кальциевых скелетов и оболочек.
Авторы публикации, Род Уилсон (Rod Wilson) из британского Эксетерского университета вместе с коллегами из Канады и США, обнаружили ранее неизвестный фактор, влияющий на кислотно-щелочной баланс океана.
С помощью компьютерного моделирования они установили, что синтез и выделение карбоната кальция в кишечнике костистых рыб, оказывает значительное, если не определяющее, влияние на кислотность океана.
Костистые рыбы, составляющие до 90% морской фауны (хрящевых рыб, к которым относятся акулы и скаты, значительно меньше) потребляют много воды, чтобы избежать обезвоживания. Вместе с водой в их организм попадает избыточное количество кальция, который выводится из организма в форме карбоната кальция, сдвигающего баланс воды в щелочную сторону.
Ранее океанологи полагали, что рыбы не могут значительно влиять на оборот химических элементов в экосистемах поскольку они находятся на вершине пищевой цепи и их суммарная биомасса мала по сравнению с массой фитопланктона и микроорганизмов. По их мнению, сдвиг кислотности морских вод в щелочную сторону вызван растворением карбоната кальция, из экзоскелетов микроорганизмов, а выделения рыб составляют доли процента от этой массы.
Однако в рамках этой модели океанологи долгое время не могли объяснить, почему воды океана становятся более щелочными, чем поверхностные, уже на глубине 500-1000 метров, тогда как экзоскелеты микроорганизмов не растворяются даже на существенно больших глубинах.
Костистые рыбы, в свою очередь, выделяют карбонат кальция в форме, значительно легче растворимой. Для того, чтобы показать, что именно они ответственны за повышение уровня щелочного показателя воды, ученые создали две математические модели, с помощью которых им удалось в первые получить оценочные значения биомассы всей рыбы в океане.
Согласно их расчетам сейчас в океане находится от 812 до 2050 миллионов тонн костистых рыб. Вся эта рыба производит около 110 миллионов тонн карбоната кальция в год. Это составляет от 3% до 15% всего карбоната кальция морского происхождения, однако в действительности эти цифры могут быть в три раза больше.
Ученые обеспокоены тем, что дальнейшее увеличение атмосферной концентрации углекислого газа только усилит влияние рыб на кислотность мировых вод. Во-первых, более теплый климат неизбежно приведет к ускорению всех физиологических процессов в их организме, а во-вторых увеличение концентрации СО2 в крови рыб напрямую стимулирует выработку ими карбоната кальция.
ОПИСАНИЕ СКЕЛЕТА
Осевой скелет (позвоночник). Осевой скелет костистых рыб составлен многочисленными костными позвонками. Тела позвонков спереди и сзади вогнутые — такие позвонки называют амфицельными. Пространство, образующееся между вогнутыми поверхностями соседних позвонков, и узкий канал, пронизывающий в центре тела позвонков, заполнены остатками хорды (рис. 1, 1), имеющей четковидную форму. Позвоночник делится на два отдела: туловищный (pars thoracalis) и хвостовой (pars caudalis); позвонки этих отделов отличаются своим строением.

Рис. 1. Схема продольного разреза хвостового отдела позвоночника судака: 1 — редуцированная хорда, 2 — тело позвонка, 3 — верхняя дуга, 4 — верхний остистый отросток, 5 — канал для спинного мозга, 6 — нижняя дуга, 7 — нижний остистый отросток, 8 — гемальный канал
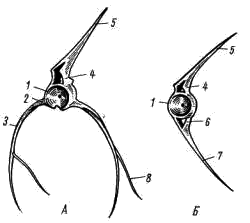
Рис. 2. Позвонки судака. А — туловищный позвонок; Б — хвостовой позвонок: 1 — тело позвонка, 2 — поперечный отросток, 3 — ребро, 4 — верхняя дуга, 5 — верхний остистый отросток, 6 — нижняя дуга, 7 — нижний остистый отросток, 8 — мышечная косточка
Туловищные позвонки (рис. 2, А) имеют округлое в поперечном сечении тело (corpus vertebrae, 2, 1), от которого в стороны отходят поперечные отростки (processus transversus, рис. 2, 2). К этим отросткам причленяются ребра (costa, рис. 2, 3), а к некоторым ребрам — тоненькие мышечные косточки (рис. 2, 8). От верхней (спинной) части позвонка отходят верхние дуги (arcus neuralis, рис. 2, 4), замыкающиеся остистым отростком (processus spinosus, рис. 2, 5). В канале, образованном верхними дугами позвонков, располагается спинной мозг.
Хвостовые позвонки (рис. 2, Б) также имеют тело и верхние дуги, но поперечные отростки их смещены вниз и образуют нижние дуги (arcus haemalis; рис. 2, 6), замыкающиеся непарным нижним остистым отростком (processus spinosus inferior; рис. 2, 7). Канал, образуемый нижними дугами хвостовых позвонков, носит название гемального; в нем располагаются хвостовые артерия и вена. Расположение этих крупных кровеносных сосудов внутри костного канала предохраняет их от сдавливания при сокращениях мощной мускулатуры хвоста.
Череп костистых рыб состоит из двух отделов: осевого черепа, или мозговой коробки (neurocranium), и лицевого, или висцерального, черепа (splanchnocranium). Но в отличие от хрящевых череп костистых рыб почти целиком образован костной тканью и состоит из многочисленных отдельных костей.
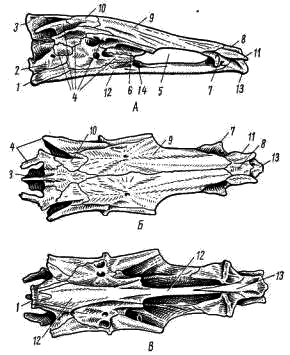
Рис. 3. Осевой (мозговой) череп судака. А — сбоку; Б — сверху; В — снизу: 1 — основная затылочная кость, 2 — боковая затылочная кость, 3 — верхняя затылочная кость, 4 — ушные кости, 5 — глазница, 6 — боковая клиновидная кость, 7 — боковая обонятельная кость, 8 — средняя обонятельная кость, 9 — лобная кость, 10 — теменная кость, 11 — носовая кость, 12 — парасфеноид, 13 — сошник, 14 — основная клиновидная кость
Осевой череп костистых рыб подразделяется на те же отделы, что и у хрящевых рыб. В каждом отделе образуется несколько костей. Затылочный отдел черепа составлен четырьмя костями: основной, или нижней, затылочной (basioccipitale; рис. 3, 2; рис 4, 1). Двумя боковыми затылочными (occipitale laterale; рис. 3, 2, рис. 4, 2) и верхней затылочной (supraoccipitale; рис. 3, 3; рис. 4, 3) Эти кости окаймляют большое затылочное отверстие (foramen occipitale magnum; рис. 4, 3), через которое головной мозг соединяется со спинным.
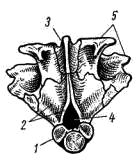
Рис. 4. Затылочный отдел осевого черепа судака: 1 — основная затылочная кость, 2 — боковая затылочная кость, 3 — верхняя затылочная кость, 4 — затылочное отверстие, 5 — ушные кости
В слуховом отделе черепа располагается по пять ушных костей (ossa otici; рис. 3, 4) с каждой стороны. В глазничном отделе имеются парные боковые клиновидные (laterosphenoideum; рис. 3, 6) и глазоклиновидные (orbitosphenoideum) кости. Последние расположены на тонкой межглазничной перегородке и при препарировке, как правило разрушаются; у судака их нет. В основании мозговой коробки образуется основная клиновидная кость (basisphenoideum); при рассматривании черепа сбоку виден лишь ее передний конец (рис. 3, А, 14). Впереди, в области обонятельного отдела, находятся кости парные боковые обонятельные (ectoethmoideum, рис. 3, 7) и непарная средняя обонятельная (mesethmoideum, рис. 3, 8). Все эти кости по происхождению замещающие (первичные), они возникли путем замещения соответствующих участков хрящевого черепа костной тканью.
Кости, прикрывающие череп сверху и снизу, по происхождению относятся к покровным (вторичным) костям. Они закладываются в соединительно-тканном слое кожи и позже, погружаясь под кожу, прирастают к хрящевой основе черепа. Среди них в крыше черепа бросаются в глаза крупные плоские лобные кости (frontale; рис. 3, 9), покрывающие большую часть черепной коробки. Сзади от них по обе стороны от гребня верхней затылочной располагаются относительно небольшие теменные кости (parietale; рис. 3, 10). Спереди от лобных в области обонятельного отдела находятся парные носовые кости (nasalе; рис. 3, 11), разделенные уже упоминавшейся средней обонятельной костью.
Дно черепа прикрыто крупной непарной костью, называемой парасфеноидом (parasphenoideum; рис. 3, 12). Впереди парасфеноида расположен непарный сошник (vomer; рис. 3, 13), в своей передней части снабженный зубами.

Рис. 5 Череп судака сбоку: 1 — окологлазничные кости, 2— слезная кость, 3 — нёбная кость, 4 — квадратная кость, 5 — крыловидные кости, 6 — сочленовная кость, 7 — предчелюстная кость, 8 — верхнечелюстная кость, 9 — зубная кость, 10 — угловая кость, 11 — подвесок, 12 — симплектикум, 13 — гиоид, 14 — копулагиоида, 15 — крышечная кость, 16 — предкрышечная кость, 17 — подкрышечная кость, 18 — межкрышечная кость
При рассматривании черепа сбоку помимо уже отмеченных костей видно полукольцо из мелких косточек, окаймляющих глазницу — окологлазничных (ossa infraorbitalia; рис. 5, 1). Передняя из них носит особое название — слезная кость (lacrimale; рис. 5, 2). Все эти кости покровные.
Висцеральный череп костистых рыб представлен серией висцеральных дуг: челюстной (самая передняя), подъязычной и пятью жаберными, из которых последняя (задняя) заметно редуцирована.
В челюстной дуге можно различить первичные и вторичные челюсти. Первичные челюсти образованы преимущественно замещающими костями и гомологичны хрящам нёбно-квадратному и меккелеву хрящевых рыб. В верхней челюсти они представлены спереди смешанной нёбной костью (palatinum; рис. 5, 3), сзади (в месте сочленения с нижней челюстью) — квадратной костью (quadratum; рис. 5, 4); между ними расположены три крыловидные кости (pterygoideum; рис. 33, 5). Из них лишь одна (задняя крыловидная — metapterygoicieum) замещающая, тогда как остальные две — наружная крыловидная (ectopterygoideum) и внутренняя крыловидная (entopterygoideum) — по происхождению покровные.
В нижней челюсти первична лишь одна сочленовная кость (articulare; рис. 5, 6), осуществляющая подвижное причленение нижней челюсти к верхней (через квадратную кость).
Вторичные челюсти представлены предчелюстными (praemaxillare; рис. 5, 7) и верхнечелюстными (maxillare; рис. 5, 8) костями в верхней половине и крупной зубной костью (dentale, рис. 5, 9) — в нижней. Кроме того, в нижней челюсти имеется еще маленькая угловая кость (angulare; рис. 5, 10), приросшая к задне-нижнему краю сочленовной. Предчелюстная и зубная кости снабжены довольно крупными зубами. Вторичным челюстям принадлежит главная хватательная функция. Дополнительную функцию удерживания добычи выполняют зубы, расположенные на нёбных костях и сошнике.
Все перечисленные кости челюстной дуги — парные; они симметрично повторяются на правой и левой половинах челюстей.
Подъязычная дуга целиком составлена первичными (замещающими) костями. Верхний элемент ее — подвесок (hyomandibulare, рис. 5, 11) верхним концом причленяется к слуховому отделу осевого черепа, а нижним — через маленькую дополнительную косточку (symplecticum; рис. 5, 12) — к квадратной кости челюстной дуги. Кроме этого, подвесок срастается с задней крыловидной костью. Таким образом, подвесок выполняет функцию причленения челюстной дуги к осевому черепу; иными словами, череп у костистых рыб, как и у акуловых, гиостилический. Нижняя ветвь подъязычной дуги, гомологичная гиоидному хрящу акуловых рыб, представлена несколькими окостенениями. Из них крупный — гиоид (hyoideum; рис. 5, 13). Гиоиды правой и левой сторон соединяются через непарную кость (copula; рис. 5, 14), одновременно поддерживающую язык. К гиоиду прикрепляются тонкие изогнутые косточки — лучи жаберной перепонки, поддерживающие кожистый край жаберной крышки.
Жаберные дуги представлены каждая четырьмя парными косточками, подвижно сочлененными между собой и объединяющимися снизу при помощи непарных костных элементов — копул. Исключение составляет лишь пятая, самая задняя жаберная дуга, имеющая лишь один парный (нижний) отдел.
Жаберные крышки являются по сравнению с хрящевыми рыбами новообразованием. Каждая состоит из четырех вторичных (покровных) костей: крышки (operculum; рис. 5, 15), предкрышки (praeoperculum; рис. 5, 16), подкрышки (sub>operculum; рис. 5, 17) и межкрышки (interoperculum; рис. 5, 18). Жаберная крышка каждой стороны через предкрышечную кость прикрепляется к соответствующему подвеску и к квадратной кости.
Парные конечности и их пояса. Парные конечности представлены грудными и брюшными плавниками. Опорой грудных плавников в теле рыбы служит плечевой пояс (рис. 6). Он представлен двумя небольшими замещающими (первичными) и несколькими покровными костями. Верхняя из замещающих костей — лопатка (scapula; рис. 6, 1) — расположена в области причленения свободной конечности (ее легко отличить по небольшому круглому отверстию в центре кости). Сразу под ней находится врановая кость, или коракоид (coracoideum; рис. 6, 2). Эти два элемента составляют первичный пояс. Они неподвижно соединены с крупной покровной костью клейтрум (cleithrum; рис. 6, 3), верхний конец которой направлен несколько вперед; к нему присоединяется небольшая кость надклейтрум (supracleithrum; рис. 6, 4).

Рис. 6. Плечевой пояс и грудной плавник судака: 1 — лопатка, 2 — коракоид, 3 — клейтрум, 4 — надклейтрум, 5 — заднетеменная кость, 6 — заднеключичная кость, 7 — радиалии, 8 — костные кожные лучи
Клейтрум
в свою очередь соединяется с заднетеменной
(posttemporale; рис. 6, 5) костью. Направленные
вперед нижние концы правого и левого
клейтрумов соединяются друг с другом.
Позади клейтрума неподалеку от лопатки
и коракоида расположена небольшая
заднеключичная кость (postcleithrum; рис. 6,
6). Все названные кости парные; они
составляют вторичный плечевой пояс.
Правая и левая заднетеменные кости
причленяются к осевому черепу, что
обеспечивает более прочную фиксацию
пояса и таким образом усиливает его
опорную функцию.
Грудной плавник в
своем основании имеет один ряд мелких
косточек — радиалии (radialia; рис. 6, 7),
отходящих от лопатки (частично и от
коракоида). Вся свободная лопасть
плавника состоит из членистых кожных
лучей1 (lepidotrichia; рис. 6, 8). Особенность
скелета грудных плавников костистых
рыб, по сравнению с хрящевыми, заключается
в редукции базалий. Подвижность грудных
плавников увеличивается потому, что
мышцы прикрепляются к расширенным
основаниям кожных лучей, подвижно
сочленяющихся с радиалями.
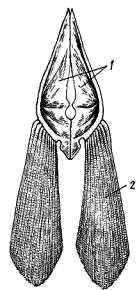
Рис. 7. Тазовый пояс и брюшной плавник судака: 1 — тазовые кости, 2 — кожные костные лучи
Тазовый пояс (рис. 7) представлен сливающимися друг с другом парными плоскими треугольными костями, лежащими в толще брюшной мускулатуры и не связанными с осевым скелетом. К боковым сторонам тазового пояса причленяются брюшные плавники. У большинства костистых рыб в скелете брюшных плавников отсутствуют базалий и полностью редуцировались радиалии: лопасть плавника поддерживается кожными костными лучами (lepidotrichia), расширенные основания которых непосредственно причленяются к тазовому поясу. Такое упрощение скелета брюшных плавников, видимо, связано с их ограниченными функциями.
Непарные конечности. Непарные конечности представлены спинными, подхвостовым (анальным) и хвостовым плавниками. Анальный и спинные плавники состоят из костных лучей, подразделяющихся на внутренние (скрытые в толще мускулатуры) птеригофоры и наружные плавниковые лучи — лепидотрихии.
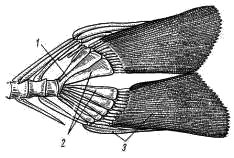
Рис. 8. Хвостовой плавник судака: 1 — конец позвоночного столба — уростиль, 2 — гипуралин, 3 — костные кожные лучи — лепидотрихии
Хвостовой плавник, как это было видно уже при наружном осмотре, имеет внешне равнолопастное строение, однако при рассмотрении его внутреннего скелета (рис. 8) видно, что концевые позвонки позвоночного столба сливаются в палочковидную косточку — уростиль (urostyl; рис. 8, 1), которая заходит в основание лишь верхней лопасти плавника, а основание нижней лопасти поддерживается разросшимися, довольно широкими нижними дугами позвонков — гипуралиями (рис. 8, 2). Такой тип строения хвостового плавника, вполне или почти симметричного по внешней форме, но асимметричного по расположению внутреннего скелета, носит название гомоцеркального. Наружный скелет хвостового плавника составлен многочисленными кожными лучами — лепидотрихиями (рис. 8, 3).
ЗАКЛЮЧЕНИЕ
Общий тип организации водных челюстноротых позвоночных, сформировавшийся у хрящевых рыб, претерпевает дальнейшее развитие в классе костных рыб. Сохраняя общий план строения предков, костные рыбы в своей эволюции приобрели ряд прогрессивных черт, способствовавших повышению уровня жизнедеятельности и широкой адаптивной радиации этих животных.
Наиболее существенное отличие костных рыб от хрящевых заключается в развитии у них костного скелета. Биологическое значение этого явления очень велико: превосходя хрящ в прочности, костная ткань придает скелету еще и большую легкость и подвижность.
Развитие костной ткани происходит двумя независимыми путями. С одной стороны, имеют место довольно сложные гистологические процессы, в результате которых хрящевая ткань, составляющая основу скелета зародыша, постепенно замещается костной. Так возникают замещающие (хрящевые, или первичные) кости. Первичный костный скелет принимает на себя практически все те функции, которые у хрящевых рыб принадлежали хрящевому скелету.
С другой стороны, костная ткань может формироваться непосредственно в соединительнотканном слое кожи независимо от хряща. Таким путем развиваются покровные (кожные, или вторичные) кости. Именно так возник в эволюции костных рыб ряд новых элементов скелета, имеющих большое биологическое значение. Не будучи связанными с первичными (исходными) функциями хрящевого скелета, вторичные костные образования открывают возможность большого разнообразия форм, отражающих приспособления к разным условиям жизни. Таковы, например, вторичные челюсти, принимающие на себя хватательную функцию и отличающиеся большим разнообразием строения у разных видов в связи с приспособлением их к различным типам питания, гораздо более многообразным, чем у хрящевых рыб. Только вторичные кости образуют жаберную крышку, имеющую большое значение в активизации дыхания. Вторичный плечевой пояс осуществляет связь первичного пояса с черепом. При характерном для рыб неподвижном соединении черепа с позвоночником такая связь определяет собой более прочную фиксацию пояса и усиление его опорной функции.
В эволюции рыб сначала покровные кости возникали в добавление к хрящевому скелету (состояние, характерное, например, для осетровых рыб), а позднее начался процесс окостенения хряща и образование замещающих костей; одновременно покровные окостенения начали погружаться под кожу, соединяясь с замещающими (хрящевыми) костями.
Во внутреннем строении костистых рыб наиболее бросающаяся в глаза особенность — появление плавательного пузыря — гидростатического органа, увеличивающего «плавучесть» и позволяющего рыбам маневрировать без существенной затраты энергии. У хрящевых рыб это возможно только при движении, что, естественно, требует значительной затраты энергии. Плавательный пузырь выполняет и некоторые добавочные функции: служит резонатором издаваемых рыбой звуков, может служить резервуаром для накопления резервного запаса кислорода (а у некоторых видов — и органом воздушного дыхания) и т. п.
Отсутствие спирального клапана, свойственного хрящевым рыбам, компенсируется у костистых рыб увеличением относительной длины кишечника и развитием у многих видов пилорических придатков, также увеличивающих общую всасывающую поверхность кишечника. Эти преобразования способствуют усилению интенсивности и эффективности пищеварения.
Своеобразно строение мочеполовой системы костистых рыб. Они, как и хрящевые рыбы, обладают мезонефрическими (туловищными) почками с мочеточниками, соответствующими вольфовым каналам. В отличие от хрящевых костистые рыбы имеют мочевой пузырь. Что же касается половых протоков костистых рыб, то они представляют собой особые образования, не гомологичные ни вольфовым, ни мюллеровым каналам. Эти особенности возникают в результате изменений хода эмбрионального развития гонад и, по-видимому, связаны с приспособлением к выведению большого количества половых продуктов; плодовитость костистых рыб значительно выше, чем хрящевых. Впрочем, рассмотренные особенности мочеполовой системы являются специфическим свойством только костистых (и некоторых других костных) рыб и не получили дальнейшего развития в эволюции позвоночных животных.
Перечисленные морфологические особенности, а также и ряд прогрессивных физиологических свойств (большая, чем у хрящевых рыб, кислородная емкость крови, некоторые особенности пищеварения, осморегуляции и т. п.) обеспечили костным рыбам возможность значительно более широкой адаптивной радиации как по характеру местообитаний, так и по пищевой специализации.
Костные рыбы заселяют в гидросфере практически все жизненные ниши. Среди этих животных имеются как пелагические морские виды, так и придонные формы, в том числе обитатели больших глубин, хорошо приспособившиеся к высокому давлению и отсутствию освещения. Столь же широко они заселяют и пресные водоемы — как текучие (реки, ручьи), так и стоячие (в том числе и пересыхающие).
Широкий диапазон осморегуляционных процессов позволяет костным рыбам заселять водоемы с разной соленостью и даже в течение индивидуальной жизни переходить из морей в пресные водоемы и наоборот (так называемые проходные рыбы, например многие лососевые). Осморегуляция у костистых рыб построена на совершенно ином принципе, чем у хрящевых. Пресноводные формы имеют хорошо развитую систему фильтрации воды в почках и выделяют большое количество жидкой мочи, избавляя тем самым организм от избытка воды, постоянно поступающей осмотическим путем через жабры и кожу. Морские рыбы, организм которых в силу разницы его осмотического давления с таковым среды все время обезвоживается, имеют слабо развитую клубочковую систему в почках, выделяют небольшое количество сильно концентрированной мочи и пьют много морской воды, выделяя избыток получаемых при этом солей через почки и через жабры.
Ряд физиологических адаптации позволяет костным рыбам жить в водоемах, резко отличающихся содержанием кислорода, температурой и другими условиями.
Очень многообразны адаптации костных рыб к различным типам и способам питания. Этот класс включает и плотоядные, и растительноядные формы (среди хрящевых рыб — только плотоядные); многие виды характеризуются смешанным питанием. Среди костных рыб имеются хищники (которые активно охотятся за крупной добычей или подстерегают ее в засаде) и питающиеся микроскопическими планктонными организмами. Разные виды рыб добывают пищу на поверхности воды, в толще ее, среди подводных зарослей, камней, в коралловых рифах, на поверхности дна и в толще ила и т. д. Все эти формы обладают специальными приспособлениями в строении вторичных челюстей, форме тела, устройстве плавников, окраске и т. п.
Комплекс прогрессивных особенностей строения костных рыб особенно отчетливо и полно выражен у наиболее молодой и прогрессивной ветви этого класса — костистых рыб Teleostei, которая включает подавляющее большинство ныне живущих форм этого класса (около 20 000 видов).
Литература:
Гуртовой Н.Н., Матвеев Б.С., Дзержинский Ф.Я. Практическая зоотомия позвоночных. Низшие хордовые, бесчелюстные, рыбы. М., 1976.
Никольский Г.В. Частная ихтиология. М., 1971.
Никольский Г.В. Экология рыб. М., 1974.
Строганов Н.С. Экологическая физиология рыб. М., 1962.
Суворов Е.К. Основы ихтиологии. М., 1948.
Шмальгаузен И.И. Основы сравнительной анатомии позвоночных. М., 1947.