Архебактерии в биокомпьютерах
Архебактерии - первые на Земле, первые в биокомпьютерах?
Архебактерии
достаточно давно известны как объект
самых разнообразных микробиологических,
генетических и биохимических исследований
(Stoeckenius et al., 1979, Sapienza, Doolittle, 1982, Pfeifer,
Betlach,1985). Общепризнанно, что этот интерес
связан, прежде всего, с их способностью
использовать энергию солнечного света
для частичного обеспечения энергетических
потребностей внутриклеточных процессов
биосинтеза (Katznelson, Lochhead, 1952), а также с
уникальными требованиями к ростовым
условиям, выраженными потребностью в
необычайно высоком содержании хлорида
натрия (отсюда название наиболее
изученного рода архебактерий -
галобактерии).
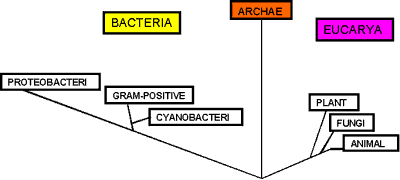
Таксономические
исследования представителей Archaebacteria
практически всегда подчёркивали
своеобразие и необычность этих
микроорганизмов. Новейшие системы
классификации, основанные на сопоставлении
биохимических и генетических признаков,
чаще всего подтверждают обособленность
представителей данного таксона как от
истинных бактерий, так и от эукариотических
микроорганизмов (Woese et al., 1990, May, Dennis,
1990, Mukohata et al., 1991, Jochi, Dennis, 1993, Ruepp et al., 1995,
Goldman, Kranz,1998). В частности, одним из важных
таксономических признаков микроорганизмов
является состав клеточных мембран.
Оказалось, что в отличие от эубактериальных
и эукариотических организмов, имеющих
липиды на основе спиртов и жирных кислот,
липиды архебактерий в основном
представлены изопраниловыми эфирами
глицерина и синтезируются путём
конденсации глицерина с изопреноидными
спиртами с числом углеродных атомов
20, 25 или 40 (DeRosa, Gambacorta, 1988).
Многими
исследователями высказывалась мысль,
что именно архебактерии были основной
формой жизни на Земле в первый её период.
Одним из важных аргументов в пользу
этой идеи может быть то обстоятельство,
что представители многих видов
архебактерий используют в качестве
единственного источника углерода - то
есть в качестве ростового субстрата
для биосинтеза всех компонентов клеточной
массы - различные аминокислоты. Как
известно, именно обнаружение аминокислот
в Космосе и в модельных первичных
бульонах - коацерватных каплях - считается
важным доводом в пользу возможности
зарождения биологической жизни вообще
и на нашей планете в частности. Можно
предположить, что питательная среда
для архебактерий на Земле могла
сформироваться на самых ранних этапах
биологической эволюции.
Сейчас, по
мнению ряда специалистов, продолжение
исследований археобактерий требует
пристального изучения других таксономически
значимых признаков, например, ферментативных
реакций новых видов. (Oren, 1994).
К
сожалению, на сегодняшний день генетические
и биохимические исследования архебактерий,
и в частности, галобактерий отличает
заметная "однобокость" - большинство
полученных и охарактеризованных
мутантов, а также клонированных
структурных и регуляторных участков
генов так или иначе связаны с одной,
действительно очень интересной
биологической системой - системой
фоторецепции. Справедливо было бы
отметить, что к настоящему времен
накоплено довольно много информации и
о других клеточных системах представителей
Archaebacteria, например, транспорта
экзогенных соединений в клетки. Однако
эти сведения нельзя считать исчерпывающими.
Сказанное относится как к транспорту
сахаров - веществ, оказывающих сравнительно
слабое влияние на рост и функционирование
клеточных систем представителей этого
таксона, так и к транспорту аминокислот
- основного источника углерода, азота
и энергии. Это обстоятельство, казалось
бы, должно было послужить поводом для
тщательного изучения особенностей
поступления и последующих превращений
аминокислот в клетках архебактерий.
Тем не менее, данных о проведении
подобного рода целенаправленных
исследований не появлялось в научной
периодике уже несколько лет.

Скопление
соли на берегу Мертвого моря -
идеального
места для обитания представителей рода
Halobacterium.
Вместе
с тем, по мере расширения использования
галофильных бактерий в качестве
промышленных продуцентов тех или иных
биологически активных соединений (БАС),
наблюдается всё более пристальный
интерес и к накоплению новых знаний о
биохимических и других особенностях
этих микроорганизмов. На сегодняшний
день наибольший объём информации собран
о галобактериях вида Halobacterium salinarium
(ранее данный вид носил название H.
halobium).
Следует отметить, что, как
показали исследования, галобактерии
довольно чувствительны к изменениям в
соотношениях компонентов ростовых
сред. Выше уже отмечалась способность
большинства представителей Archaebacteria
расти на питательных средах с аминокислотами
в качестве единственного источника
углерода и одного из двух источников
энергии - имеется в виду, что вторым
источником энергии является фотопроцесс
(MacDonald et al., 1977, Lany 1978). Для выращивания
галобактерий в значительных объёмах
используются ростовые среды, разнообразные
по составу и содержанию компонентов (в
зависимости от задач эксперимента), но
с двумя обязательными составляющими -
хлоридом натрия в высокой концентрации
и тем или иным набором аминокислот.
Наиболее широко применяемым источником
последних, при культивировании
галобактерий, являются всевозможные
гидролизаты белков (или биомассы)
микробного, растительного или животного
происхождения. Многие биохимические
фирмы выпускают сейчас превосходные
препараты белковых гидролизатов с
достаточно точно охарактеризованным
и воспроизводимым из года в год составом.
Подавляющее большинство лабораторий
успешно проводит исследования, нарабатывая
биомассу на средах с такими препаратами.
Идеальным вариантом гидролизата может
считаться препарат казаминовых кислот
- продукт полного кислотного гидролиза
казеина. В ряде случаев используют более
дешевые гидролизаты, зачастую с
неконтролируемым составом. Применение
сред, приготовленных на таких ростовых
субстратах, разумеется, оправдано
экономически, но в ряде случаев может
приводить к невоспроизводимости
результатов. В ряде работ отмечалось,
что, например, варьирование солевого
состава полноценных сред, приводит к
изменению уровня накопления биомассы
и формы клеток галобактерий (Brown, Gibbons,
1955). В целом, однако, бактерии Halobacterium
на полноценных средах обнаруживают
хороший рост. Витамины выступают как
стимуляторы на ранних стадиях развития
культуры, углеводы являются стимуляторами
только при отсутствии лимита по ионам
калия. Таким образом, современные
галобактерии, унаследовавшие свои
уникальные свойства от первых архебактерий,
можно, в общем и целом, считать довольно
"не привередливыми" в отношении
ростовых субстратов.
Особого
внимания, на мой взгляд, заслуживает
рассмотрение применения синтетических
сред, то есть сред с абсолютно чётко
заданным аминокислотным и солевым
составом. Многие исследователи применяли
простую тактику - добавляли в ростовую
среду галофилов смесь чистых препаратов
аминокислот (Оnishi et al., 1965). Соотношение
концентраций отдельных аминокислот в
статьях разных авторов несколько
отличалось, отражая, по-видимому, то
обстоятельство, что у них не было
необходимости оптимизировать данные
параметры (Gochnaker, Kushner, 1969). Было установлено,
что рост клеток сильно замедляется,
если из среды исключить аргинин, валин,
лейцин или лизин. При отсутствии метионина
и глицерина рост не ухудшался, но заметно
снижался уровень красных пигментов в
клетках (Kushaza, Kates, 1979). Нуклеиновые
компоненты, как оказалось, стимулируют
рост только при нехватке в среде
источников фосфатов (Оnishi et al., 1965).
Возможно то обстоятельство, что
галобактерии сохранили способность
самостоятельно синтезировать из
нескольких аминокислот большинство
необходимых для жизнедеятельности
клеток БАС, отражает высокую
распространённость именно этих
аминокислот в первичном бульоне на
Земле.
Совершенно особое значение
приобретают синтетические ростовые
среды для архебактерий, если они
применяются в прецизионных
молекулярно-биологических или
биохимических исследованиях тех или
иных БАС. Такие научно-экспериментальные
работы зачастую сопряжены с введением
в изучаемые молекулы атомов изотопов.
Поскольку довольно часто изотопные
метки по азоту или углероду вводят в
состав белков именно через введение в
ростовую среду меченых аминокислот,
несколькими исследовательскими группами
было определено "минимальное
необходимое" их число и требуемые
концентрации. В настоящий момент широко
используется ростовая среда, содержащая
десять аминокислот: аргинин, валин,
изолейцин, лейцин, лизин, метионин,
серин, тирозин, треонин, фенилаланин
(Оnishi et al., 1965). Потребность бактерий в
остальных аминокислотах восполняется
за счёт внутриклеточных метаболических
превращений, происходящих по известным
схемам. Так, по литературным данным
[4-13C]-аспарагиновая кислота, как
метаболический предшественник метионина,
треонина и лизина, не может обеспечить
специфического мечения молекул
бактериородопсина, а включает 13C-метку
в этот белок по аминокислотным остаткам
ещё и глутамина, пролина, треонина,
лизина (Engeihard et al., 1985). "Перекрёстное"
мечение наблюдали и при введении в
синтетическую ростовую среду
15N-аминокислот: 15N-метка
валина обнаруживалась в лейциновых
остатках, 15N-лейцин давал
15N-изолейцин, 15N-валин и
15N-глютамин, 15N-фенилаланин
давал 15N-триптофан и следы
15N-тирозина (Orechov et al., 1992).
Скорее
всего, все-таки существует принципиальная
корреляция между наличием или отсутствием
определённых метаболических систем
архебактерий, составом ростовых сред
для галофильных бактерий и аминокислотным
составом "первичного бульона"
Земли. Однако, прямого экспериментального
пути проверить это предположение на
сегодня, к сожалению, нет. Возможно, что
дальнейшие исследования помогут его
найти. В настоящее время ряд мировых
научных центров и лабораторий проявляет
значительный интерес к галофильным
бактериям как источнику бактериородопсина
(БР) - белка с уникальными свойствами,
имеющего хорошие возможности для
применения в оптоэлектронных устройствах.
Следует надеяться, что необходимость
получения значительных количеств БР и
его мутантных и меченых стабильными
изотопами аналогов потребует существенного
продвижения по части изучения генетических
и биохимических особенностей представителей
этого рода архебактерий.
Автор
статьи - главный научный сотрудник ФГУП
"ГосНИИгенетика",
д.б.н. Складнев
Д.А