Антиатеросклеротическое действие смеси масел льна и расторопши с селенопираном
ФЕДЕРАЛЬНОЕ АГЕНТСТВО ПО ОБРАЗОВАНИЮ
Пензенский государственный педагогический университет
имени В.Г. Белинского
Факультет Кафедра
Естественно-географический Биохимии
ДИПЛОМНАЯ РАБОТА НА ТЕМУ:
Антиатеросклеротическое действие смеси масел льна и расторопши с селенопираном
Студент________________________________________Краснова Ю.В.
Руководитель___________________________________Петрушова О. П.
________________Кравченко Ю. В.
К защите допустить. Протокол № от «_____» ____________2007 г.
Зав. Кафедрой______________________________________Генгин М. Т.
Пенза, 2007 год
СПИСОК СОКРАЩЕНИЙ
АО - антиоксиданты
АКМ – активные кислородные метаболиты
АС - атеросклероз
БОТ – бутилокситолуол
ВЖР – высокожировой рацион
ГМК - гладкомышечные клетки
ДК – диеновые коньюгаты
ИА - индекс атерогенности
ЛНП – липопротеиды низкой плотности
ЛОНП - липопротеиды очень низкой плотности
ЛП – липопротеиды
ЛВП - липопротеиды высокой плотности
мГМК - модифицированные ГМК
МДА – малоновый диальдегид
мЛНП - модифицированные ЛНП
ПНЖК – полиненасыщенные жирные кислоты
ПОЛ – перекисное окисление липидов
СМЛР - смеси масел льна и расторопши с селенопираном
СП - селенопиран
ТГ - триглицеридов
ТФ - токоферол
ХС - холестерин
ХС- ЛВП – холестерин липопротеидов высокой плотности
ХС- ЛНП - холестерин липопротеидов низкой плотности
ХС- ЛОНП - холестерин липопротеидов очень низкой плотности
ХС>общ> -> >общий холестерин
ЭХС – эфиры холестерина
ВВЕДЕНИЕ
В настоящее время в диетотерапии и профилактике различных заболеваний, особенно сердечно-сосудистой системы, широко используются полиненасыщенные жирные кислоты (ПНЖК) семейства Омега-3. В многочисленных клинических и экспериментальных исследованиях выявлен положительный эффект применения данных жирных кислот, связанный с их влиянием на липидный обмен, иммунологическую реактивность и механизмы свертывания крови. Большинство подобных исследований [26, 27, 31] посвящено изучению эффективности ПНЖК, входящих в состав рыбных жиров. Растительным жирам в этом смысле уделено гораздо меньше внимания, хотя традиционным в России сырьем для получения масла, богатого альфа-линоленовой кислотой (семейства Омега-3), является семя льна. Комбинируя масло семян льна с другими растительными маслами, можно добиться оптимального соотношения ПНЖК класса Омега-3 и Омега-6, рекомендованного для целей диетотерапии. Существенным недостатком пищевого масла, богатого ПНЖК, является низкий срок хранения из-за высокой способности этих жирных кислот к окислению, поэтому актуальным является поиск способа стабилизации продукта. Второй аспект проблемы заключается в биологической целесообразности употребления такого масла. Известно, что в условиях недостатка в организме антиоксидантов поступление в него ПНЖК может приводить к индукции перекисного окисления липидов [36]. Решением этих вопросов может служить введение в композицию масла расторопши пятнистой с комплексом флаволигнанов [10] и синтетического антиоксиданта (АО). В качестве жирорастворимого АО в данной работе исследовалось органическое соединение селена – 9-фенил-симметричный октагидроселеноксантен (селенопиран – СП). Обладая высокой водородо- и электронодонорной активностью, данное соединение препятствует образованию перекисей [3]. Кроме того, СП в живом организме подвергается метаболизации в системе микросомального окисления [4], после чего из него высвобождается атом селена, встраивающийся в селенопротеины, основной функцией которых является антиоксидантная защита.
Целью нашей работы было исследование антиоксидантных свойств селенопирана in vitro, а также изучение антиатеросклеротического действия смеси масел льна и расторопши, стабилизированной селенопираном при моделировании экспериментального атеросклероза.
При выполнении работы были поставлены следующие задачи:
Исследовать количество липидов плазмы крови и их соотношение.
Изучить влияние смеси масел льна и расторопши, стабилизированной селенопираном, на процессы перекисного окисления липидов.
Выявить влияние смеси масел льна и расторопши, стабилизированной селенопираном, на процесс атерогенеза.
Научная новизна. Впервые выявлено антиатеросклеротическое действие смеси масел льна и расторопши, стабилизированной селенопираном in vivo при моделировании экспериментального атеросклероза. Выявлены различия действия исследуемого масла в клетках (эритроцитах, печени) и плазме крови.
Апробация работы. Материалы работы доложены на Научной студенческой конференции ПГПУ им. В.Г. Белинского.
ГЛАВА 1. ОБЗОР ЛИТЕРАТУРЫ
Атеросклероз (АС) – патологическое изменение внутреннего слоя (интимы) и мышечного слоя (медии) артерии. Слово образовано от греч. «атере» - кашица, основной компонент которой – эфиры холестерина (ЭХС), включенные в уплотненную склерозированную бляшку [2]. Клиническими проявлениями АС являются различные широко распространенные сосудистые заболевания разных органов: сердца, аорты, головного мозга, почек, нижних конечностей. Атеросклеротическая бляшка затрудняет кровоток и вызывает ишемию и гипоксию органа, а при полной закупорке сосуда бляшкой или индуцированным ею тромбом возникают грозные осложнения АС: инфаркт миокарда, инсульт, гангрена конечностей [2].
1.1.Биохимические ОСНОВЫ ПАТОГЕНЕЗА АТЕРОСКЛЕРОЗА
По мере накопления экспериментальных данных по атеросклерозу, возникли разные гипотезы об этиологии заболевания и патогенезе. Все гипотезы связаны с транспортом липопротеидов (ЛП) и холестерина (ХС) в сосудистую стенку. Повышенный уровень ХС является, бесспорно, главным патогенетическим фактором развития атеросклероза, существенную роль играют и другие факторы риска: факторы повреждения эндотелия (инфекции, воспаления, гипертония, токсические факторы, например никотин, высокие содержания липопротеинов низкой плотности (ЛНП) и липопротеинов очень низкой плотности (ЛОНП), повышенная свертываемость крови, гемодинамические факторы (бляшки возникают в местах изгибов, ветвления, стенозов сосудов) и изменение содержания гормонов, но, пожалуй, важнейшим фактором можно назвать химическую и физико-химическую модификацию ЛНП (гликозилирование, перикисное окисление ХС и ПНЖК в ЛНП, десиалирование, частичный протеолиз, агрегация) [2].
В состав атеросклеротических бляшек входят: ЭХС, триацилглицерины, гликозаминогликаны, коллаген, эластин, кальций, макрофаги, модифицированные гладкомышечные клетки (мГМК), погибшие клетки. В атеросклеротических бляшках с большой частотой обнаруживаются ДНК герпесвирусов (цитомегаловируса и простого герпеса), а также хламидии. Роль этих микроорганизмов остается пока невыясненной. При разрыве фиброзной капсулы под действием механических и токсических факторов обнажается холестериновая кашица, и такая злокачественная бляшка становится центром агрегации тромбоцитов, отложения сгустков фибрина и тромбообразования [2].
Учитывая все перечисленные этиологические и патогенетические факторы, можно привести пример механизма интегральной модели АС артериальной стенки. В норме ЛНП крови транспортируют в эндотелий и другие клетки сосуда ХС, необходимый им для формирования мембран. При гиперхолестеринемии, обусловленной повышенным уровнем ЛНП, или при модификации ЛНП, главный поток ХС, во внутреннюю стенку сосудов, осуществляют моноциты. Последние фагоцитируют ЛНП, модифицированные ЛНП (мЛНП), аутоиммунные комплексы мЛНП с антителами и переносят ХС в субэндотелиальный участок интимы. ЛНП разрушаются в эндолизосоме, освобождая ХС, который накапливается в цитозоле макрофагов в виде капель ЭХС, придавая клеткам пенистый вид («пенистые» клетки). Этерификацию ХС катализирует ацетил-коэнзим А-холестеринацилтрансфераза, активируемая свободным ХС. «Пенистые» клетки, погибая освобождают ХС в межклеточное пространство для формирования бляшек [2].
Источником ХС бляшек так же служат гладкомышечные клетки (ГМК). Активированные макрофаги продуцируют в интиме цитокины (интерлейкин – 1 и др.), которые стимулируют миграцию ГМК из медии сосуда в интиму, а так же их пролиферацию и трансформацию в ГМК синтезирующего типа («миома сосуда»). Последние продуцируют коллаген и другие структуры соединительной ткани фиброзной бляшки. мГМК также фагоцитируют ЛНП и мЛНП, превращаясь в «пенистые » клетки [2].
Около 70% ХС крови содержится в составе ЛНП. Уровень ХС в крови зависит от огромного количества факторов: поглощаемой пищи, синтеза и регуляции, транспорта, расходования на синтез жирных кислот и других соединений с выведением. При патологических процессах нарушается динамическое равновесие между указанными факторами, что может привести к гиперхолестеринемии. Следовательно, гиперхолестеринемия – главный фактор риска развития атеросклероза, как правило, связана с увеличением содержания в крови ЛНП и их предшественников ЛОНП – двух атерогенных ЛП. Поэтому более строгим фактором риска следует считать не гиперхолестеринемию, а дислипопротеинемию – количественное преобладание ЛНП, ЛОНП и качественные изменения в структуре ЛНП (мЛНП) [2]. В формировании модифицированных липопротеидов центральную роль играют процессы свободнорадикального окисления (перекисное окисление липидов (ПОЛ)).
1.2. ПЕРЕКИСНОЕ ОКИСЛЕНИЕ ЛИПИДОВ
Из всех классов ЛП ПОЛ затрагивает в первую очередь именно ЛНП [14]. ПОЛ в частицах ЛНП - сложный многоступенчатый и еще не до конца выясненный процесс. Постоянно возникающие в животном организме свободные радикалы О->2>, HO*>2> и НО- генерируют образование гидроперекисей ненасыщенных жирных кислот, входящих в состав фосфолипидов, триглицеридов (ТГ) и ЭХС ЛНП [14].
ПОЛ – цепная свободнорадикальная реакция. На первых стадиях 92-98 % всех продуктов окисления составляют гидроперекиси, образованию которых предшествует гидроперекисный радикал ROO* (образуется в акте одноэлектронного окисления с появлением сопряженных двойных связей) [Камышников В.С., 2000]. Преобразование липидов в гидроперекиси жирных кислот (ацилгидроперекиси – первичные продукты ПОЛ) приводит к образованию «пор» в мембранах, через которые наружу выходит содержимое клетки и органелл. Первичные продукты ПОЛ неустойчивы и разрушаются. Вторичные продукты ПОЛ: альдегиды, кетоны, спирты и эпоксиды. Продукты ПОЛ способствуют агрегации тромбоцитов, уменьшению синтеза простагландинов, антикоагулянтному действию и разрушению мембран [12].
Трудно точно назвать место, где образуются перекисно-модифицированные ЛНП. По-видимому, значительная часть их образуется в крови в результате окисления липидов в этих комплексах при участии клеток белой крови, генерирующих активные кислородные радикалы. Поскольку гранулоцитарные лейкоциты не проникают в интиму артерий, есть основания полагать, что их участие в окислении ЛНП происходит не в артериальной стенке, а именно в кровотоке. Другая потенциальная возможность перекисной модификации частиц возникает в процессе циркуляции в кровяном русле в результате контакта ЛП с клетками эндотелия. Аналогичной способностью модифицировать ЛНП обладают и другие клетки: ГМК, активированные моноциты и лейкоциты и даже фибробласты. Речь идет, следовательно, о перекисной модификации ЛП, индуцированной клетками [14]. .В образовании мЛНП участвуют так же ферменты, объединенные общим названием липоксигеназы. Нельзя исключить и то, что процесс пероксидации ЛНП может продолжаться и в самой интиме вследствие контакта с другими клетками (макрофагами и др.) и недостатка в интиме АО [14].
Таким образом, множество данных указывает на то, что реакции свободнорадикального окисления липидов являются необходимым атрибутом процесса атерогенеза.
1.3. АТЕРОГЕНЕЗ
Многие исследователи отмечают, что необходимым условием атерогенеза является повреждение стенки сосуда, что приводит к секвестрации и накоплению в интиме фагоцитирующих клеток с последующим их перерождением в «пенистые» клетки. Тот факт, что формирование «пенистых» клеток происходит при инкубации макрофагов только с окисленными или модифицированными («атерогенные ЛНП»), но не с нативными ЛНП, послужил основой концепции, согласно которой, начальным этапом атерогенеза является возникновение окисленных ЛНП, цитотоксичных для эндотелиоцитов и усиливающих адгезию нейтрофилов, что вызывает повреждение эндотелия; захват окисленных ЛНП макрофагами и приводит к образованию «пенистых» клеток, их накоплению в интиме сосудов и в последующем – к формированию атеросклеротической бляшки [20, 33, 37, 38]. Таким образом, свободнорадикальные реакции с участием активированных кислородных метаболитов (АКМ) являются атерогенным фактором, так как вызывают модификацию ЛП (преимущественно ЛНП) плазмы крови; кроме того АКМ и продукты их реакции, в том числе и окисленные ЛП, оказывают цитотоксическое и деструктивное воздействие на клетки и ткани [23, 29], что может лежать в основе повреждения сосудов [16]. Множество экспериментальных, клинических и эпидемиологических исследований позволяют отнести АС к классическим свободно радикальным патологиям [1, 6].
В моноцитах (макрофагах) и в эндотелии сосудов основную роль в поглощении окисленных ЛНП играют не ЛНП-рецепторы, а особые нерегулируемые скэвенджер-рецепторы, (рецепторы – «мусорщики») для модифицированных ЛНП [21]. Общим признаком всех лигандов для скэвенджер – рецепторов является наличие полианионных комплексов («рецептор-липучка»). Скэвенджер – рецепторы связывают только модифицированные ЛНП, и их экспрессия не регулируется внутриклеточным содержанием холестерина [21]. Больше всего скэвенджер – рецепторов выявляется в моноцитах/ макрофагах [34].
Проникновению моноцитов в эндотелиальную стенку предшествует их адгезия на поверхности эндотелия, по-видимому, в ответ на появление в интиме мЛНП и секрецию хемотаксических веществ - хемоатрактантов. Адгезивные молекулы обеспечивают прилипание моноцитов на поверхности эндотелия. Часть проникших в интиму моноцитов/макрофагов подвергается в ней пролиферации [35].
Значительная часть макрофагов после захвата ими мЛНП и накопления в них ЭХС трансформируется в «пенистые» клетки и остается в интиме. Подавляющая часть «пенистых» клеток гибнет, при этом в интиму изливаются накопленные в них ЭХС, ненасыщенных эфиров холестерина и кристаллы моногидрата ХС. Образуются очаговые скопления ХС и создается угроза развития сначала липидных пятен, а затем и атеросклеротических бляшек [13].
Оценивая участие макрофагов в атерогенезе, следует сказать, что эти клетки, захватывая мЛНП, стремятся выжить и покинуть артериальную стенку вместе с захваченными ЛП. Об этом свидете6льствует активация пролиферации макрофагов в интиме при появлении в ней мЛНП и миграция макрофагов из интимы в кровяное русло еще до превращения в «пенистые» клетки. Есть и другие пути избавления стенки сосуда от ХС [13]:
- гидроксилирование ХС и превращение его в более растворимые производные, которые клетка секретирует в окружающую среду.
- усиленный синтез макрофагами аполипопротеина Е и фосфолипидов с последующим образованием дискоидальных комплексов, способных связывать и удалять ХС из клетки.
- гидролиз ЭХС и удаление ненасыщенных эфиров холестерина при участии ЛВП.
- уменьшить содержание ХС и ЛНП в крови могут такие гормоны, как иодтиронины и эстрогены. Иодтиронины индуцируют на уровне транскрибции образование ЛНП-рецепторов и 7-гидроксилазы печени – регуляторного фермента синтеза желчных кислот из ХС. Эстрогены ингибируют образование последнего фермента, но активируют синтез ЛНП-рецепторов и ГМГ-СоА-редуктазы печени (регуляторный фермент синтеза ХС) [2].
Одним из важных защитных механизмов можно считать действие различных АО, которые являются ингибиторами образования мЛНП.
1.4. АНТИОКСИДАНТЫ В ЖИВОМ ОРГАНИЗМЕ
В живом организме функционирует внутренняя система антиоксидантной защиты, представленная ферментами (глутатионпероксидаза, супероксиддисмутаза, каталаза и глутатионредуктаза) и низкомолекулярными соединениями (тиоловые соединения, мочевая кислота, некоторые пептиды). Помимо этого, многие низкомолекулярные соединения, образующиеся в организме или поступающие с пищей (водорастворимые или жирорастворимые), способны служить в качестве компонентов антиоксидантной защиты клеток, внутри- и межклеточной жидкости (аскорбиновая кислота, мочевая кислота, токоферол, флавоноиды, каротиноиды, и др.). Неферментные АО- низкомолекулярные соединения, перехватчики радикалов и активных кислородных метаболитов. Общим признаком очень многих АО является их способность выступать в качестве донора протона функциональной группы. По строению функциональной группы важнейшие для человека и млекопитающих низкомолекулярные АО можно разделить на соединения, содержащие SH-группы (тиоловые АО) и ОН- группы (фенольные АО) [15].
Тиоловые соединения
В свете современных данных становится все более очевидной ведущая роль тиоловых соединений в механизме антиоксидантной защиты. К такому заключению приводят несколько обстоятельств [19]. К примеру, основная часть функциональных компонентов антиоксидантной системы представлена веществами тиоловой природы. Так, в состав неферментного звена входят низкомолекулярные тиолы (глутатион, тиоредоксин и др.) и тиол содержащие белки, которые по некоторым данным [32] даже более реактивны по отношению к АКМ, чем глутатион. К тиоловым компонентам сыворотки крови млекопитающих относятся альбумины, представляющие собой важные внеклеточные АО [28]. Ферменты, принимающие участие в противоокислительной защите, либо относятся к собственному числу тиоловых энзимов, либо нуждаются в присутствии тиолов для проявления каталитической активности. Во-вторых, уникальные химические свойства тиолов наделяют их высокой антиокислительной способностью [15].
Фенольные соединения, имеющие в своей структуре ароматическое кольцо с несколькими ОН-группами, также являются мощными перехватчиками радикалов, эффективность которых возрастает в зависимости от количества гидроксильных заместителей цикле [15].
Важную роль в защите клеток, и главным образом, их мембран от окислительных повреждений играют токоферолы (ТФ), антиокислительное действие которых реализуется двумя путями. Во-первых, ТФ способны стабилизировать мембраны за счет хорошей растворимости в фосфолипидах и взаимодействия их с жирнокислыми цепями, что увеличивает плотность упаковки фосфолипидов в мембране и снижает вероятность их окисления. Вторым проявлением антиоксидантных свойств токоферолов является их способность перехватывать АКМ и ингибировать ПОЛ, в результате чего образуются малоактивные токоферильные радикалы, которые легко восстанавливаются аскорбиновой кислотой, убихиноном или мочевой кислотой, что обеспечивает регенерацию витамина Е [11, 22].
Одним из наиболее важных водорастворимых низкомолекулярных АО считают аскорбиновую кислоту, действие которой распространяется на широкий спектр АКМ. Способность аскорбиновой кислоты восстанавливать токоферил-радикалы обеспечивает синергическое действие системы аскорбат-токоферол в гетерогенных средах, содержащих водную и липидную фазы [24].однако в присутствии ионов металлов переменой валентности аскорбиновая кислота способна восстанавливать их, и, таким образом проявлять прооксидантное действие [8].
Мочевая кислота, помимо способности хелатировать ионы железа и меди, обладает прямым антиоксидантным действием за счет способности ингибировать оксиды азота, супероксид-анион радикал, гидроксильный радикал и синглетный кислород, а также гемовые оксиданты. Реакция мочевой кислоты с гидроксильным радикалом при физиологических рН в большинстве случаев приводит к конформационной перестройке молекулы с образованием аллантоина, в свою очередь способного окисляться гидроксильным радикалом до парабановой кислоты [11]. Ввиду высокого содержания мочевой кислоты в плазме крови (0,12-0,48 мМ), некоторые исследователи считают, что на ее долю приходится 35-65% защиты ЛП от окисления, 10-15% ингибирования гидроксильного радикала и 12% ингибирования синглетного кислорода.
В плазме крови имеется большое количество других соединений, способных взаимодействовать с АКМ и тормозящих развитие свободнорадикальных реакций посредством хелатирования ионов переменой валентности. К ним относятся железосвязывающие белки: ферритин, гемосидерин и трансферрины; медьсвязывающий фермент церулоплазмин; молочная и мочевая кислот; некоторые пептиды, гормоны и гормоноподобные вещества [11].
Таким образом, учитывая ведущую роль в патогенезе атеросклеротических повреждений процессов окислительной модификации ЛП, весьма интересным представляется изучение влияния пищевых АО (особенно жирорастворимых) в условиях гиперхолестеринемии, сопровождающейся окислительным стрессом, на атерогенез. Не менее важным представляется проанализировать комплексное воздействие эссенциальных жирных кислот класса Омега-6, Омега-3 и АО, так как известно, что ПНЖК являются быстроокисляемым субстратом (т.е. могут провоцировать развитие антиоксидантной системы), с одной стороны, и незаменимыми регуляторами состояния эндотелия, тонуса сосудов и процессов свертывания крови (т.е. могут препятствовать развитию антиоксидантной системы), с другой стороны.
2. МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
2.1. ЭКСПЕРИМЕНТАЛЬНЫЕ ЖИВОТНЫЕ И УСЛОВИЯ ОПЫТОВ
Изучение антиатеросклеротического действия СМЛР выполнено в эксперименте на животных в общепринятой модели. Эксперимент проведен на 30 половозрелых белых крысах-самцах Вистар весом 270-410 г. В зависимости от условий эксперимента животные были разделены на 3 равные по численности группы: контрольную и две опытные.
Таблица 1
Состав экспериментальных рационов
|
Наименование |
Рацион (количество в г.) |
|||
|
1 |
2 |
3 |
4 |
|
|
Казеин |
197 |
197 |
197 |
197 |
|
Метионин |
3 |
3 |
3 |
3 |
|
Крахмал |
445 |
445 |
280 |
280 |
|
Сахар |
175 |
175 |
175 |
175 |
|
Отруби |
50 |
50 |
50 |
50 |
|
Масло подс. контр.* |
45 |
35 |
10 |
- |
|
Холина хлорид |
5 |
5 |
5 |
5 |
|
Солевая смесь** |
70 |
70 |
70 |
70 |
|
Витаминная смесь*** |
10 |
10 |
10 |
10 |
|
Холестерин |
- |
- |
10 |
10 |
|
СМЛР**** |
- |
10 |
- |
10 |
|
Масло окисленное***** |
- |
- |
190 |
190 |
* масло в 45 г содержит: ретинола ацетата 15000 МЕ, эргокальциферола – 500 МЕ
** состав солевой смеси представлен в таблице 2
*** в 1 г содержит: тиамина (В>1>) – 0,6 мг, рибофлавина (В>2>) – 0,6 мг, пиридоксина (В>6>) – 1,5 мг, никотиновой кислоты – 3,0 мг, пантотената кальция – 1,6 мг, фолиевой кислоты – 1,0 мг, цианкобаламина (В>12>) – 0,003 мг, викасола – 0,1 мг, Д-биотина – 0,02 мг, токоферола ацетата – 15 мг, глюкозы – до 1 г.
**** - СМЛР содержит (г/100 г): нерафинированное масло льна – 14,3 , нерафинированное масло расторопши – 85,7, селена - 0,001 в составе селенопирана, жирнокислотный состав см. в таблице 3
***** - масло окисленное в 200 г содержит: ретинола ацетата 15000 МЕ, эргокальциферола – 1500 МЕ
Таблица 2
Состав солевой смеси
|
№ |
Название соли |
Химическая формула |
Количество, г |
|
1 |
Хлористый натрий |
NaCl |
139,3 |
|
2 |
Калий фосфорнокислый, однозамещенный |
KH3PO4 |
388,8 |
|
3 |
Магний сернокислый |
MgSO4 |
57,4 |
|
4 |
Кальций углекислый |
CaCO3 |
380,4 |
|
5 |
Железо сернокислое |
FeSO4*7H3O |
26,4 |
|
6 |
Калий йодистый |
KI |
0,77 |
|
7 |
Марганец сернокислый |
MnSO4*7H30 |
4,55 |
|
8 |
Цинк сернокислый |
ZnSO4*7H3O |
0,53 |
|
9 |
Медь сернокислая |
CuSO4*5H3O |
0,48 |
|
10 |
Кобальт хлористый |
CoCl2*6H3O |
0,024 |
|
11 |
Натрий фтористый |
NaF |
0,50 |
|
12 |
Алюмокалиевые квасцы |
K>2>SO4Al2(SO4)3*24H3O |
0,11 |
|
ИТОГО |
1000 |
На первом этапе эксперимента (14 дней) животные контрольной группы и первой опытной группы получали рацион, содержащий (по калорийности) 20% белка (казеин), 70% углеводов, 10% жиров (подсолнечное масло) (таблица 1, рацион 1). Животные второй опытной группы получали рацион с замещением части жира СМЛР (таблица 1, рацион 2). Витамины и минеральные вещества добавляли в соответствии с физиологическими нормами (таблица 1 и таблица 2). Корм животным давали вволю, доступ к воде был свободным.
На втором этапе эксперимента (28 дней) животные контрольной группы получали рацион 1, животные опытных групп получали высокожировой рацион (ВЖР), содержащий 10 г/кг холестерина и 40% жиров по калорийности (подсолнечное масло, перекисное число которого составляло 38 ммоль активного кислорода на 1 кг продукта). В первой опытной группе 5% жира замещали обычным (неокисленным) подсолнечным маслом (таблица 1, рацион 3), во второй опытной группе – СМЛР (таблица 1, рацион 4).
Таблица 3
Жирнокислотный состав смеси масел льна и расторопши (СМЛР)
|
Наименование |
Количество, % |
|
Сумма насыщенных жирных кислот |
13,71 |
|
Сумма мононенасыщенных жирных кислот |
27,39 |
|
Сумма ПНЖК, из них: |
58,99 |
|
Линолевая (-6) С18:2 |
50,1 |
|
-линоленовая (-6) С18:3 |
0,03 |
|
-линоленовая (-3) С18:3 |
8,86 |
|
Соотношение -6/-3 |
5,66 |
2.2. ПОДГОТОВКА БИОЛОГИЧЕСКОГО МАТЕРИАЛА ДЛЯ БИОХИМИЧЕСКОГО И ГИСТОЛОГИЧЕСКОГО ИССЛЕДОВАНИЯ
После окончания эксперимента животных выдерживали 12 часов без корма, затем наркотизировали тиопенталом натрия в дозе 6 мг на 100 г веса. После чего вскрывали брюшную полость, пунктировали брюшную аорту и собирали кровь в пробирки с гепарином, центрифугировали 15 мин при 2000 об/мин. Готовые гемолизаты эритроцитов (разведение 1:1) и плазму затем замораживали и хранили при – 18°С- – 20°С до исследования. Затем перфузировали печень in situ через соустье полой вены охлажденным физиологическим раствором. Далее извлекали печень, промывали охлажденным физраствором, продавливали через металлический пресс с диаметром отверстий 0,2 мм и готовили гомогенаты. Печень гомогенизировали в стеклянном гомогенизаторе Поттера-Эльвейма с тефлоновым пестиком (зазор 0,18-0,2 мм) в течение 90 с при 1200 об/мин. Гомогенат хранили при –18°С- – 20°С.
Для гистологического исследования брали грудной и брюшной отдел аорты. Аорту фиксировали 10% формалином, вскрывали по длине, проводили макроскопическое исследование, далее готовили срезы методом парафиновой проводки, окрашивали гематоксилином + эозином, проводили микроскопическое исследование
2.3. СПЕЦИАЛЬНЫЕ МЕТОДЫ ИССЛЕДОВАНИЯ
2.3.1. Изучение антиоксидантных свойств селенопирана in vitro
Изучение антиоксидантных свойств СП in vitro проводили в системе нерафинированных растительных масел: подсолнечного, смеси масел льна и расторопши пятнистой. В модельной системе с подсолнечным маслом для сравнения использовали бутилокситолуол (БОТ) и ТФ. При этом концентрация АО (в том числе и СП) составляла 0,01% от массы. Контролем служило чистое масло. Окисление проводили при температуре 80 С, пропуская через масло воздух со скоростью 5 л/ч. Автоокисление смеси масел льна и расторопши (1:6, v/v) проводили в мягких условиях – при комнатной температуре без принудительного пропускания воздуха. СП или ТФ добавляли в смесь масел в концентрации 0,005%. Контролем служила смесь масел без АО. Количество пероксидов определяли йодметрическим титрованием [ГОСТ 26593-85].
2.3.2. Определение перекисного числа йодометрическим титрованием по ГОСТ 26593-85
Метод основан на окислении йодистого калия перекисями и гидроперекисями, содержащимися в образце, в растворе уксусной кислоты и хлороформа и титровании выделившегося йода раствором тиосульфата натрия.
Пробу отвешивают в колбу или пробирку, помещают навеску в колбу. Добавляют 10 см3 хлороформа, растворяют навеску, приливают 15 см3 уксусной кислоты и 1 см3 раствора йодистого калия, колбу сразу же закрывают, перемешивают содержимое в течение 1 мин и оставляют на 5 мин в темном месте при температуре 15-25°С. Затем добавляют 75 см3 воды, тщательно перемешивают и добавляют пять капель раствора крахмала. Выделившийся йод титруют раствором тиосульфата натрия, используя раствор следующей концентрации в зависимости от предполагаемого значения перекисного числа. С (½Na>2>S>2>O>3>) = 0,002 моль\дм3 , если перекисное число не более 6,0 моль\кг или с (½Na>2>S>2>O>3>) = 0,01 моль\дм3, если перекисное число равно или более 6,0 моль\кг [Издательство стандартов, 1994]. Массу пробы, необходимой для измерений, в зависимости от предполагаемого перекисного числа определяют по таблице:
|
Предполагаемое значение перекисного числа, моль\кг |
Масса испытуемой пробы, г |
|
От 0 до 6,0 |
5,000-2,000 |
|
От 6,0 до 10 |
2,000-1,200 |
|
От 10,0 до 15,0 |
1,200-0,600 |
|
От 15,0 до 25,0 |
0,600-0,500 |
|
От 25,0 до 40,0 |
0,500-0,300 |
Для каждой испытуемой пробы выполняется два измерения. Контрольные измерения проводят параллельно с основными измерениями. Если на контрольное измерение пойдет более 0,1 см3 (0,01 моль\дм3) раствора тиосульфата натрия, то проверяют соответствие реактивов стандарту.
Вычисление результатов измерения: Перекисное число Х в миллимолях активного кислорода на килограмм пробы вычисляют по формуле
Х = (V>1>-V>0>)*c*1000 \ m
V>0>> >–объем раствора тиосульфата натрия, использованный при контрольном измерении, см3
V>1 >-> >объем раствора тиосульфата натрия, использованный при измерении, см3
C – концентрация использованного раствора тиосульфата натрия моль\дм3
1000 – коэффициент учитывающий пересчет результата измерения в ммоль\кг
m – масса испытуемой пробы, г
За результат измерения принимают среднее арифметическое значение двух параллельных измерений.
2.3.3. Определение концентрации общего холестерина (ХС>общ>) в сыворотке и плазме крови энзиматическим колориметрическим методом
ПРИНЦИП МЕТОДА. При гидролизе ЭХС холестеразой образуется свободный ХС. Образуется и имеющийся в пробе ХС: окисляется кислородом воздуха под действием холестеролоксидазы с образованием эквимолярного количества перекиси водорода. Под действием (РОД) перекись водорода окисляет хромогенные субстраты с образованием окрашенного продукта. Интенсивность окраски пропорциональна концентрации в пробе.
Набор реагентов: CHOLESTEROL «E – D» («Vital-Diagnostics»)
ПРОВЕДЕНИЕ АНАЛИЗА
|
Реагенты |
Опытные пробы,(мл.) |
Калибровочные пробы, (мл.) |
Контрольные пробы, (мл.) |
|
Сыворотка или плазма |
0,02 |
_ |
_ |
|
Физ. Раствор (H>2>O) |
_ |
_ |
0,02 |
|
Реагент 2 |
2,0 |
2,0 |
2,0 |
|
Раствор холестерина |
_ |
0,02 |
_ |
Реакционную смесь перемешивают и инкубируют не менее 5 мин при комнатной температуре (20-25°С) или при 37°С и измеряют оптическую плотность опытных и калибровочных проб против контрольной пробы в кювете с толщиной поглощающего слоя 5 см (1 см.) при длине волны 500 нм (ФЭК- 490 нм). Окраска стабильна не менее 2-х часов после окончания инкубации при предохранении от прямого солнечного света.
Расчет концентрации холестерина проводят по формуле:
С = [Е>о>/Е>ст >]> * >5,17 (ммоль/л) или С =[ Е>о>/Е>ст >]> * >200 (мг/100 мл)
Е>о> и Е>ст> – экстинция образца и стандарта, измеряется относительно контрольной пробы.
2.3.4. Определение концентрации триглицеридов (ТГ) в сыворотке и плазме крови энзиматическим колориметрическим методом
Т глицериды
липаза глицерин + жирная
кислота
глицериды
липаза глицерин + жирная
кислота
Г лицерин
+ АТФ глицерокиназа
глицерил-3-фосфат +2 H>2>O
лицерин
+ АТФ глицерокиназа
глицерил-3-фосфат +2 H>2>O
Г лицерил-3-фосфат
+ O>2> ГФО диоксиацетон
фосфат +2 H>2>O
лицерил-3-фосфат
+ O>2> ГФО диоксиацетон
фосфат +2 H>2>O
H>2>O>2
> + 4-ААР + 4-хлорфенол пероксидаза
хинонимин + 4H>2>O
Набор реагентов: TRIGLYCERIDES «E – D» («Vital-Diagnostics»)
ПРОВЕДЕНИЕ АНАЛИЗА
|
Реагенты |
Опытные пробы, (мл.) |
Калибровочные пробы, (мл.) |
Контрольные пробы, (мл.) |
|
Сыворотка или плазма |
0,02 |
_ |
_ |
|
Физ. Раствор (H>2>O) |
_ |
_ |
0,02 |
|
Реагент |
2,0 |
2,0 |
2,0 |
|
Стандартный раствор |
_ |
0,02 |
_ |
Реакционную смесь перемешивают и инкубируют не менее 5 мин при комнатной температуре (20-25°С) и измеряют оптическую плотность опытных и калибровочных проб против контрольной пробы в кювете с толщиной поглощающего слоя 5 см (1 см.) при длине волны 505 нм (ФЭК- 490 нм). Окраска стабильна не менее часа после окончания инкубации при предохранении от прямого солнечного света. (таблица).
Расчет концентрации триглицеридов проводят по формуле:
С = [Е>о>/Е>ст *>250] – 10 (мг/100мл) или С = [Е>о>/Е>ст *>2,85] – 0,11ммоль/л
Е>о> и Е>ст> – экстинция образца и стандарта, измеряется относительно контрольной пробы. 10 мг/100мл (0,11ммоль/л) – поправка на содержание свободного глицерина в сыворотке (плазме крови).
2.3.5. Определение концентрации ХС-ЛВП (холестерина-липопротеидов высокой плотности) (HDL) в сыворотке и плазме крови
ПРИНЦИП МЕТОДА. Хиломикроны, липопротеиды низкой плотности (LDL) и липопротеиды очень низкой плотности (V LDL) осаждаются при добавлении к образцу фосфовольфрамат/Mg²+. После центрифугирования в супернатанте остаются только HDL, концентрация которых определяется так же как концентрация общего холестерина.
Набор реагентов: HDL CHOLESTEROL «FL - E» («Vital-Diagnostics»)
ПРОВЕДЕНИЕ АНАЛИЗА
1. Преципитация
|
Реагенты |
Опытные пробы, (мл.) |
Калибровочные пробы, (мл.) |
Контрольные пробы, (мл.) |
|
Сыворотка или плазма |
0,15 |
_ |
_ |
|
H>2>O |
_ |
_ |
0,15 |
|
Осаждающий реагент |
0,3 |
0,3 |
0,3 |
|
Раствор холестерина |
_ |
0,15 |
_ |
Хорошо перемешать и оставить на 10 мин при комнатной температуре. Опытные пробы отцентрифугировать в течение 10 мин при 4000g или 1-2 мин 1000g. Прозрачный супернатант использовать для определения концентрации HDL. Калибровочные и контрольные пробы центрифугировать не нужно. Определение холестерина во всех пробах в течение часа.
2. Определение концентрации HDL
|
Реагенты |
Опытные пробы, (мл.) |
Калибровочные пробы, (мл.) |
Контрольные пробы, (мл.) |
|
Супернатант |
0,2 |
_ |
- |
|
H>2>O+ реагент №1 |
_ |
_ |
0,2 |
|
Рабочий реагент для определ. холестерина |
0,2 |
0,2 |
0,2 |
|
Раствор холестерина+реагент №1 |
_ |
0,2 |
_ |
Реакционную смесь тщательно перемешивают и инкубируют не менее 10 мин при комнатной температуре (20-25°С) или 5 мин при 37°С и измеряют оптическую плотность опытных и калибровочных проб против контрольной пробы в кювете с толщиной поглощающего слоя 5 см (1 см.) при длине волны 500 нм (ФЭК- 490 нм). Окраска стабильна не менее 2-х часов после окончания инкубации при предохранении от прямого солнечного света.
Расчет концентрации (С) холестерина HDL проводят по формуле:
С = Е>о>/Е>ст * >1,29 (ммоль/л) или С = Е>о>/Е>ст * >50 (мг/100 мл) Е>о> и Е>ст> – экстинция образца и стандарта, измеряется относительно контрольной пробы.
2.3.6. Определение ХС-ЛОНП (холестерина-липопротеидов очень низкой плотности) расчетным методом
Уровень ХС-ЛОНП находят путем деления показателя концентрации триацилглицеринов на 2,2 (ТГ/2,2). Вывод этой формулы расчетов базируется на том, что, во-первых, при отсутствии в крови хиломикронов (а они после 12-часового голодания у большинства пациентов не выявляются) практически все количество ТГ крови сосредоточено в ЛОНП и ,во-вторых, при не слишком высокой концентрации в крови (когда содержание ТГ не привышает 4,4 ммоль/л) в частицах ЛОНП на 22 молекулы ТГ приходится 10 молекул холестерола [12].
2.3.7. Определение ХС-ЛНП (холестерина липопротеидов низкой плотности) расчетным методом
Зная уровень ХС>общ> плазмы, ХС-ЛВП и ХС-ЛОНП, нетрудно определить содержание ХС-ЛНП по формуле [12]:
ХС-ЛНП = ХС - (ХС-ЛВП + ХС-ЛОНП)
2.3.8. Определение индекса атерогенности (ИА) расчетным методом
Расчет индекса атерогенности призводят по формуле:
ХС>пл >– £-ХС /£-ХС Х (ЭХС/СХС)
ХС>пл >– общий холестерол плазмы
£-ХС (альфа-ХС) – холестерол супернатанта (ЛПВП)
ЭХС/СХС – отношение содержания эфиросвязанного и свободного холестерола супернатанта плазмы [12] .
2.3.9. Определение содержания диеновых коньюгатов(ДК) в эритроцитах
За основу взят классический метод Placer в модификации Гаврилова В. Б. [9] для эритроцитов. 0,5 мл гемолизата эритроцитов (1:1) смешивают в пробирках с притертой крышкой с 4 мл чистоперегнанных растворов изопропанол : гептан (1:1). Смесь встряхивают постоянно не менее одного часа затем добавляют 1 мл HCl (pH=2) после чего еще раз встряхивают две минуты, добавляют 2 мл чисто перегнанного гептана и снова встряхивают 10-15 мин. Примерно через 1-2 часа отбирают верхнюю фазу и фотометрируют при 232нм против контрольной пробы (А>фон>) где вместо гемолизата взято 0,5 мл H>2>O. Коэффициент экстинции при 233 нм –
2,2 * 10 -5 см-1 М-1. Формула для расчета: С(нмоль/мл эр)= (А>оп >- А>фон>) * 36,4
2.3.10. Определение содержания ДК в плазме крови и гомогенате печени
За основу взят классический метод Гаврилова В.Б. и соавт [9], Владимирова Ю.А. и соавт. [7]. 0,4 мл плазмы вносят в пробирку с плотной крышкой и добавляют 4 мл чисто перегнанной смеси изопропанол : гептан (1:1). Смесь встряхивают в течение 20-30 мин затем добавляют 1 мл HCl (pH=2) после чего еще раз встряхивают две минуты, добавляют 2 мл чисто перегнанного гептана и снова встряхивают 10-15 мин. Примерно через час отбирают верхнюю фазу и фотометрируют при 232 нм против контрольной пробы где вместо гемолизата взято 0,2 мл H>2>O. Коэффициент экстинции 2,2 * 10 -5 см-1 М-1. Формула для расчета (при l=1 см):
С(нмоль/мл пл) = [(∆А* 106)/(2,2*105*1)]*[4/V>проб>], где: ∆А = А>оп >- А>фон, >2,2*105 – коэффициент экстинции, 1 (см) = L (толщина кюветы), 2 (мл) = V>гепт >(объем гептана)
2.3.11. Определение содержания малонового диальдегида (МДА) в эритроцитах
За основу взят классический метод Ernster et al [25] в модификации для эритроцитов. 0,5 мл гемолизата эритроцитов (1:1) смешивают с 1 мл охлажденной 5% ТХУ и тщательно растирают смесь стеклянной палочкой. После выдерживают при 0±5 °С не менее 1-2 часов смесь центрифугируют при 4000 g 30 мин. Далее тщательно отбирают 0,4мл надосадка в пробирки для кипячения и добавляют 1,0 мл 0,5% тиобарбитуровой кислоты. Смесь инкубируют в кипящей воде более 10 мин после чего охлаждают и фотометрируют при 532 нм против соответствующего контроля. В расчет принимают коэффициент молярной экститнции – 1,56*105 см-1 М-1. С(нмоль/мл эр) = ∆А * 115,8
2.3.12. Определение содержания МДА в плазме крови и печени
За основу взят классический метод Mihara et al. [30]. 0,4 мл плазмы крови смешивают с 4 мл охлажденной 1,4% ортофосфорной кислоты и 2 мл 0,5% тиобарбитуровой кислоты. Смесь инкубируют в кипящей бане 45 мин после чего охлаждают и добавляют 4 мл н-бутанола. Смесь встряхивают до образования белой суспензии, после чего центрифугируют при 4000 g 20 мин. Верхнюю фазу фотометрируют при 532 нм против контрольной пробы. Коэффициент экстинции – 1,56*105 см-1 М-1. Формула для расчета (при L=1 см). С (нмоль/мл )= ∆А * 64,2
2.3.13. Определение жирнокислотного состава масел методом газожидкостной хроматографии (метод внутренней нормализации)
1 мл масла смешивают с 5 мл метанола, добавляют 1 мл метилирующего агента (раствор гидроксида тетраметиламмония) и 3 мл эфира серного (диэтиловый). Выдерживают в течение 10-15 мин при комнатной температуре, аликвотную часть отбирают для анализа (3-5 мкл) и хроматографируют.
Режим разделения: Температура колонки 170-190°С, Температура впрыскивающей камеры 250°С, Колонка насадочная 3 м, Содержание колонки: 5% диэтиленгликольсукцинат нанесенный на хроматон N-AW. Анализ проводят относительно количества жирных кислот.
2.3.14. Гистологический анализ
Выполняли для подтверждения наличия атеросклеротических изменений в опытных группах. Для гистологического исследования брали грудную и брюшную аорту, фиксировали 10% нейтральным формалином. Гистологическое исследование выполняли сотрудники гистологической лаборатории.
3. РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
3.1. ИССЛЕДОВАНИЯ АНТИОКСИДАНТНЫХ СВОЙСТВ СЕЛЕНОПИРАНА
Данные исследования антиоксидантных свойств СП по сравнению с БОТ, ТФ и СП при термическом окислении подсолнечного масла (Таблица 1) свидетельствуют о том, что антиоксидантная активность СП не уступает активности БОТ, и даже превосходит ее.
Рисунок 1
Динамика накопления пероксидов в смеси нерафинированных растительных масел в присутствии бутилокситолуола, селенопирана и токоферола
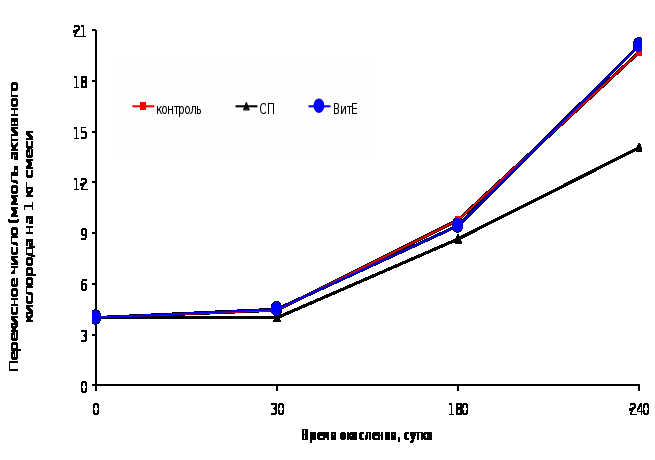
Преимущество СП становится заметным уже в течение 1-го часа окисления. Отмечено, что введение в смесь СП приводило к снижению концентрации пероксидов, и только к 6 часу их накопление достигло величин, превышающих исходные. К 26 часу окисления концентрация пероксидов в системе с БОТ составляла 95,65%, с СП – 85,51%, с ТФ – 111,6% по отношению к контролю.
Таблица 1
Динамика накопления пероксидов в подсолнечном масле в присутствии бутилокситолуола, селенопирана и токоферола
|
Часы |
Концентрация пероксидов, ммоль/г |
|||
|
контроль |
БОТ |
СП |
ТФ |
|
|
0 |
0,0060 |
0,0060 |
0,0060 |
0,0060 |
|
1 |
0,0090 |
0,0070 |
0,0045 |
0,0060 |
|
3 |
0,0100 |
0,0080 |
0,0055 |
0,0100 |
|
6 |
0,0131 |
0,0130 |
0,0090 |
0,0170 |
|
11 |
0,0300 |
0,0250 |
0,0210 |
0,0280 |
|
18 |
0,0480 |
0,0460 |
0,0380 |
0,0520 |
|
26 |
0,0690 |
0,0660 |
0,0590 |
0,0770 |
При щадящем режиме окисления смеси растительных масел СП в качестве АО также имел преимущества перед ТФ (Рис. 1), что позволило выбрать его для стабилизации итогового продукта – композиции нерафинированных масел льна и расторопши.
3.2. ИЗМЕНЕНИЯ БИОХИМИЧЕСКИХ ПОКАЗАТЕЛЕЙ ТКАНЕЙ ЭКСПЕРИМЕНТАЛЬНЫХ ЖИВОТНЫХ
Результаты комплексного биохимического и гистологического анализа биоматериала, полученного от экспериментальных животных, свидетельствуют о существенном торможении развития атеросклеротических процессов при введении СМЛР в качестве добавки к ВЖР.
Достоверные различия между опытными группами обнаруживались как в количестве липидов плазмы крови, так и в их соотношении (Таблица 2). В первой опытной группе наблюдались характерные признаки нарушения липидного обмена: повышение ХС>общ> на 54% и ХС-ЛНП на 350 % по сравнению с контрольной группой, на фоне снижения содержания ХС-ЛВП на 57 %. При этом ИА повышался до 6,29 по сравнению с 1,03 в контроле. Введение в ВЖР животных второй опытной группы СМЛР привело к торможению накопления в плазме ХС, снижение на 38 % и снижение концентрации ХС атерогенных ЛП: ХС-ЛНП на 272% и ХС-ЛОНП на 55% по сравнению с контролем, а также к повышению содержания ХС-ЛВП 42%, что отразилось на величине ИА – 3,91.
Таблица 2
Липидограмма плазмы экспериментальных животных
|
Группа |
ТГ ммоль/л |
ХС общ., ммоль/л |
ХС-ЛВП, ммоль/л |
ХС-ЛНП, ммоль/л |
ХС-ЛОНП, ммоль/л |
Индекс Атероген., у.е. |
|
Контроль n=10 |
0,906 0,031 |
1,74 0,052 |
0,858 0,024 |
0,459 0,049 |
0,421 0,019 |
1,03 0,069 |
|
Опытная 1 n=10 |
0,512# 0,044 |
2,68# 0,083 |
0,372# 0,01 |
2,07# 0,08 |
0,240# 0,018 |
6,29# 0,393 |
|
Опытная 2 n=10 |
0,442# 0,015 |
2,40# * 0,083 |
0,496# * 0,025 |
1,71# * 0,077 |
0,191# * 0,007 |
3,91# * 0,235 |
Примечание: * - p<0,05 относительно опытной группы 1
# - p<0,05 относительно контрольной группы
Было также установлено, что применение СМЛР существенно влияло на процессы ПОЛ. Выявлено достоверное снижение ДК плазмы на 12% , МДА эритроцитов на 37% и печени на 11% животных второй опытной группы по сравнению с животными, не получавшими СМЛР (Таблица 3).
Таблица 3
Содержание ДК и МДА в плазме, эритроцитах и печени экспериментальных животных
|
Группа |
ДК плазмы, нмоль/мл |
ДК эритроц., нмоль/мл |
ДК печени, нмоль/г |
МДА плазмы, нмоль/мл |
МДА эритроц., нмоль/мл |
МДА печени, нмоль/г |
|
Контроль n=10 |
3,06 0,097 |
13,2 0,482 |
47,0 1,14 |
11,6 0,068 |
53,8 0,914 |
36,7 0,656 |
|
Опытная 1 n=10 |
3,24 0,101 |
14,3# 0,168 |
85,5# 3,21 |
13,4# 0,107 |
54,2 3,30 |
50,6# 1,52 |
|
Опытная 2 n=10 |
2,68 # * 0,127 |
16,3# * 0,655 |
87,9 2,32 |
13,1 0,051 |
33,9 # * 1,48 |
32,8# * 0,362 |
Примечание: * - p<0,05 относительно опытной группы 1
# - p<0,05 относительно контрольной группы
3.3. ГИСТОЛОГИЧЕСКОЕ ИССЛЕДОВАНИЕ ПРЕПАРАТОВ АОРТЫ ЭКСПЕРИМЕНТАЛЬНЫХ ЖИВОТНЫХ
Результаты гистологического исследования препаратов аорты представлены в таблице 4. При макроскопическом исследовании аорты у 100% животных первой опытной группы наблюдали умеренно выраженные уплотнения в брюшном и грудном отделе аорты и снижение эластичности стенки аорты.
При микроскопическом исследовании аорты животных первой опытной группы во всех образцах обнаружен склероз ветвей брюшной отдела у 100% с наложением фибрина у 100% и частичной облитерацией их просвета у 40 %. При исследовании грудного отдела аорты выявлены слабо выраженные очаговые утолщения стенки у 50 %. В целом в препаратах аорты наблюдали признаки атеросклероза в стадии сформированной фиброзной бляшки (Рис. 2).
Таблица 4
Макро- и микроскопические изменения стенки аорты экспериментальных животных
|
Признаки |
Опытная 1 |
Опытная 2 |
|||
|
% |
степень |
% |
степень |
||
|
МАКРО-скопическое исследование |
Уплотнения в брюшном отделе |
100 |
++ |
70 |
+ |
|
Уплотнения в грудном отделе |
100 |
++ |
- |
- |
|
|
Снижение эластичности стенки |
100 |
++ |
70 |
+ |
|
|
МИКРО- скопическое исследование |
Очаговые утолщения стенки в брюшном отделе |
100 |
++ |
70 |
+ |
|
Очаговые утолщения стенки в грудном отделе |
50 |
+ |
- |
- |
|
|
Наложения фибрина |
100 |
++ |
- |
- |
|
|
Склероз ветвей брюшной аорты |
100 |
+++ |
70 |
++ |
|
|
Облитерация просвета ветвей брюшной аорты |
40 |
++ |
- |
- |
Примечание: +++ сильно выражены, ++ умеренно выражены, + слабо выражены
У животных второй опытной группы при микро- и макроскопическом исследовании аорты все указанные изменения были выражены в меньшей степени.
Рисунок 2
Фиброзная бляшка в стенке брюшного отдела аорты (из препаратов 1 опытной группы), окраска гематоксилин-эозин х 250.
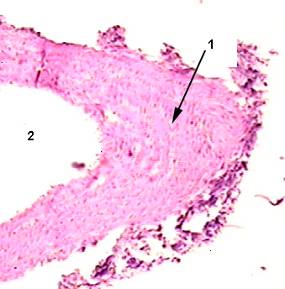

1- грубая соединительная ткань, сформированная фиброзная бляшка
2 – нарушение целостности эндотелия.
Обнаруживались лишь начальные признаки повреждения интимы и специфические «завихрения» в слое гладкомышечных волокон, ни в одном препарате не находили сформированных «бляшек», просвет ветвей аорты свободен, а в 30% случаев (у 3-х животных) признаков атеросклероза аорты вообще не выявлено.
4. ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Введение СП в сочетании с ПНЖК в атерогенный рацион оказало существенное влияние на развитие атеросклеротических процессов и на процессы ПОЛ. Вероятнее всего, это связано с антиоксидантными свойствами СП.
Липидограмма плазмы эксперементальных животных более наглядно показала действие СП, так как отличия второй опытной группы от первой было значительным. Концентрация ХС>общ>, ХС-ЛНП и ХС-ЛОНП второй опытной группы была достоверно ниже, чем у животных первой опытной группы. Данные изменения в липидограмме, в совокупности с повышением ХС фракции ЛВП, можно трактовать как положительные, так как ЛНП и ЛОНП являются атерогенными, а ЛВП - антиатерогенными частицами. ЛВП осуществляют обратный транспорт ХС из периферических органов и тканей в печень для окисления в желчные кислоты и выведения с желчью [14]. Этим можно объяснить более низкую концентрацию ХС>общ> у животных второй опытной группы в отличие от первой. Понижение концентрации ТГ объясняется снижением количества ЛОНП, которые являются транспортной формой эндогенных ТГ [14].
При исследовании продуктов ПОЛ было выявлено достоверное увеличение концентрации ДК в эритроцитах и печени, и снижение ДК в плазме у животных, получавших СМЛР. Это объясняется тем, что вдыхание кислорода приводит к десятикратному увеличению количества перекисей в печени [18], эритроциты же осуществляют транспорт кислорода и содержат мощный катализатор ПОЛ – гемоглобин [7]. Между тем, большинство тканей в норме содержит некоторое количество липидных перекисей [17]. Так, по данным Нейфаха и Кагана в печени, мозге, сальнике крыс обнаруживается от 15 до 80 нмолей перекисей на 1 г липидов или от 1 до 4,5 нмоль перекисей на 1 г ткани [18]. Следовательно, концентрация ДК, как первичных продуктов ПОЛ, высока именно в печени и эритроцитах. Накопление ДК в клетках приводит к тому, что в мембранах появляются участки («поры»), через которые наружу выходит содержимое самих клеток и органелл, а в эритроцитах возрастает перекисный и осмотический гемолиз. ДК являются неустойчивыми и через образование липоперекисей превращаются в МДА [12]. СП взаимодействует именно с перекисями жирных кислот, причем это взаимодействие имеет двойной характер. Во-первых, в нашем исследовании была показана способность СП тормозить накопление перекисей жирных кислот, а ранее автором соединения [3] было показано, что увеличение содержания перекисей в системе приводит к рециклизации соединения, протоны расходуются на инактивацию перекисей и обрыв цепи свободнорадикального окисления. При дальнейшем разрыве ковалентных связей в системе микросомального окисления с участием ферментов цитохрома р-450 из состава СП высвобождается восстановленный селен, который участвует в образовании селеноцистеина и таким образом стимулирует синтез глутатионпероксидазы [4] – главного внутри- и внеклеточного фермента, обезвреживающего липопероксиды. Таким образом, при обрыве цепи и инактивации липопероксидов, следующая степень повреждения жирных кислот – образование МДА – не наступает, чем и объясняется достоверно более низкое содержание МДА в печени и эритроцитах у животных второй опытной группы.
По результатам гистологического анализа у животных первой опытной группы были обнаружены сформировавшиеся фиброзные бляшки в отличие от второй опытной группы, у которой были найдены только липидные пятна. Большинство морфологов считают, что развитие атеросклероза протекает по схеме: липидные пятна или полоски фиброзные бляшки осложненные поражения (изъязвления, кальциноз, тромбоз), хотя отдельные авторы не исключают возможности образования фиброзных бляшек без предварительного формирования липидных пятен [5], но так же известно и то, что липидные пятна могут исчезнуть, если произойдет значительное и длительное снижение уровня ХС и ЛНП в плазме крови, и достигнутый уровень будет поддерживаться на протяжении полутора-двух лет у людей [14]. Важную роль в регрессии играют так же ЛВП, благодаря их способности осуществлять транспорт ХС в печень [14]. Можно предположить, что время опыта было слишком мало для того, чтобы увидеть, какие изменения произойдут с липидными пятнами во второй опытной группе, но даже полученные данные подтверждают положительное действие СМЛР. Возможно, СМЛР в сочетании с физическими нагрузками имела бы более выраженный положительный эффект, так как из-за малой подвижности у экспериментальных животных наблюдалось ожирение [14].
ВЫВОДЫ
1. В исследованиях in vitro доказано, что селенопиран, благодаря электроно- и водорододонорным свойствам, обладает способностью тормозить накопление пероксидов в системе нерафинированных растительных масел с различным содержанием полиненасыщенных жирных кислот в большей степени, чем бутилокситолуол и токоферол.
2. Введение в высокожировой рацион экспериментальных животных смеси масел льна и расторопши с селенопираном способствует торможению атеросклеротических процессов: снижению концентрации холестерина и атерогенных липопротеидов в плазме крови, повышению содержания антиатерогенных липопротеидов.
3. Полученные данные свидетельствуют о стабилизации перекисных процессов в липопротеидах низкой и очень низкой плотности, что подтверждается положительной корреляцией между показателями содержания диеновых конъюгатов плазмы и холестерина в липопротеидах низкой и очень низкой плотности.
4. Гистологический анализ препаратов аорты подтверждает полученные биохимические данные, и свидетельствует о значительном торможении развития морфологических изменений, вызванных атерогенной диетой, при внесении в нее испытуемой смеси масел льна и расторопши с селенопираном.
СПИСОК ЛИТЕРАТУРЫ
Абрамова Ж.И., Оксенгендлер Г.И. Человек и противоокислительные вещества. – Л.: Наука, 1985.-232с.
Биохимические основы патологических процессов / Под ред. Е.С. Северина / – М.: Медицина, 2000. –304 с.
Боряев Г.И., Жуков О.И., Блинохватов А.Ф., Древко Б.И. Оценка антиоксидантной активности селен органических соединений // Сб. науч. трудов Саратовского университета. – Саратов, 1996. – С.199.
Боряев Г.И., Галочкин В.А., Блинохватов А.Ф. Функциональная активность монооксигеназной системы печени цыплят – бройлеров при введение в рацион селеноорганического соединения СП-1 // Бюлл.ВНИИФБиП с.-х. животных. – 1990. – Вып.3 (99). – С.70-73.
Вихтер А. М. Атеросклероз // Руководство по кардиологии / Под ред. Е. И.Чазова. – М.: Медицина, 1982. – Т. 1. – С. 417 - 443.
Владимиров Ю.А., Азизова О.А., Деев А.И. и др. Свободные радикалы в живых системах // Итоги науки и техники. Сер. Биофизика. - Т.29. - М., 1991. - С.1-249.
Владимиров Ю.А., Арчаков А.И. Перекисное окисление липидов в биологических мембранах. - М.: Наука, 1972. - С.237-238.
Владимиров Ю.А. Свободные радикалы в биологических системах // Соросовский образовательный журнал. - 2000. - Т.6. - №12. - С.13-19
Гаврилов В.Б., Мишкорудная М.И. // Лаб. дело. - 1983. -№3. - С.33-35.
Гильмиярова Ф.Н. Патент РФ № 2131672, кл. А 23 D 9/00, 1998.
Зенков Н.К., Ланкин В.З., Меньщикова ЕБ. Окислительный стресс: Биохимический и патофизиологический аспекты. – М.:МАИК «Наука\Интерпереодика», 2001.-343 с.
Камышников В.С. Справочник по клинико-биохимической лабораторной диагностике. В 2 т. – Минск: Беларусь, 2000. - Т.1-2
Климов А.Н., Никульчева Н.Г Липиды и липопротеиды и атеросклероз. – СПб.: Питер, 1995.–298с.
Климов А.Н., Никульчева Н.Г. Обмен липидов и липопротеидов и его нарушения. - С-Пб: « Питер – Ком », 1999.–512с.
Кравченко Ю.В. Экспериментальное исследование системы антиоксидантной защиты на этапах онтогенеза при токсическом и алиментарном воздействии: // Дис. к.б.н.-М., 2005
Ланкин В.З., Вихерт А.М., Тихазе А.К. и др. Роль перекисного окисления липидов в этиологии и патогенезе атеросклероза // Вопр. мед. химии. - 1989. -№3. - С.18-24.
Нейфах ЕА Биофизика. – 1971. - С.16-560.
Нейфах ЕА., Каган ВЕ. Биохимия. – 1969. - С.34-511.
Соколовский В.В. Тиолдисульфидное соотношение крови как показатель состояния неспецефической резистентности организма: Учебное пособие. – С-Пб., 1996.-30 с.
Aviram M. Modified forms of low-density lipoprotein and atherogenesis // Atherosclerosis.-1993. - Vol.98.- P.1-9.
Brown M.S., Goldstein J.L. Lipoprotein metabolism in the macrophage: implication for cholesterol deposition in atherosclerosis // Annu. Rev. Biochem. - 1983. - Vol.52.- P.223-261.
Burton G., Ingold K. Vitamin E: application of the principles of physical organic chemistry to the exploration of its structure and function. // Acc. Chem. Res. – 1986. - Vol.19.- P.194 -201
Cthcart M.K., Menally A.K., Morel D.W., Chisolm G.M. Superoxid anion participation in human monocyte-mediated oxidation of low-density lipoprotein and conversion of low-density lipoprotein to a cytotoxin // J. Immunol. - 1989. -Vol.142. - P1963-1969.
Doba T., Burton G., Ingold K. Antioxidant and co- antioxidant activity of vitamin C. The effect of vitamin C, either alone or in the presence of vitamin E analogue, upon the peroxidation of agueous multilamellarphospholipid liposomes. // Biochem. biophys. аcta .- 1985. - Vol.835. - P.298 -303.
Ernster L, Nordenbrandt K. Microsomal lipid peroxidation // Methods Enzymol., 1967. - Vol. 10. - P. 575-576.
Fan Y.Y., Chapkin R.S. Importance of Dietory γ-Linolenic Acid in Human Health and Nutrition // The Journal of Nutrition. – 1998. – Vol.128. – P.1411-1414.
Frenoux J-M.R., Prost E.D., Belleville J.L., Prost J.L. A Polyunsaturated Fatty Acid How us Blood Pressure and Improves Antioxidant Status in Spontaneously Hypertensive Rats // The Journal of Nutrition – 2001. – Vol.131. – Р.39-45.
Halliwel B. Oxidation of low-density lipoproteins: guestions of initiation, propagation, and the effect of antioxidants // Amer. J. Clin. Nutr. - 1995. - Vol.61 (suppl). - P.670S-677S.
Henning B., Chow C.K. Lipid peroxidation and endothelial cell injury: Implications in atherosclerosis. Review // Free Radical Biol. and Med. - 1988. - Vol.4. - P.99-105.
Michara M., Uchiyama M., Fukuzava K. Thiobarbituric acid value on fresh homogenate of rat as a parameter of lipid peroxidation in aging, CCL4 intoxication, аnd vitamin E deficiency // Biochem. Med. 1980. - Vol. 23 (3). P. 302-311.
Nair S.D., Leitch J.W., Falconer J., Gard M.L. Prevention of Cardiac Arrhythmia by Dietory (n-3) Poly un saturated Fatty Acid and Their Mechanism of Action // The Journal of Nutrition – 1997. – Vol.127. – Р.383-393.
Park E., Thomas J. The mechanisms of reduction of protein mixed disulfides (dethiolation) in cardiac tissue. // Arch. Biochem. Biophys. – 1989. - Vol.274 (1). - P.47 -54.
Parthasarathy S., Steiberg D., Witzturm J.L. The role of oxidized low-density lipoprotein in the pathogenesis of atherosclerosis // Annu. Rev. Med. - 1992. - Vol.43. - P.219-225.
Pitas R.E. Expression of the acetyl low-density lipoprotein receptor by rabbit fibroblasts and smooth muscle cells. Up-regulation by phorbol esters // J. Biol. Chem. -1990. - Vol.265. - P.12722 -12727.
Sakai M., Miyazaki A., Hakamata H. et al. The scavenger receptor serves as a route for internalization of lysophospatidilcholine in oxidized low density lipoprotein - induced macrophage proliferation // J. Biol. Chen. – 1996. - Vol.271. - P.27346 -27352.
Schwarz K., Porter L.A., Fredga A. Some regularities in the structure-function relationship of organoselenium compounds effective against dietary liver necrosis // Ann.N.Y.Acad.Sci. – 1972. – Vol.14. – Р.192-200.
Steinbrecher U.P., Zang H.F., Lougheed M. Role of oxidatively modified LDL in atherosclerosis // Free Radical Biol. and Med. - 1990. - Vol.9. - P.155-168.
Yla-Herttuala S. Macrophages and oxidized low-density lipoprotein in the pathogenesis of atherosclerosis // Ann. Med. - 1991. - Vol.23. - P.561 -566.