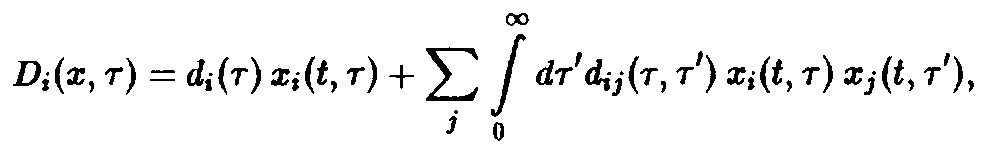Индивидуальное развитие как новая стратегия эволюции
Реферат
"Индивидуальное развитие как новая стратегия эволюции"
1. Роль онтогенеза в эволюции
Живые организмы, появившиеся 3–4 миллиарда лет назад в условиях Праземли, были еще весьма примитивны. "Протоклетка" была своего рода микрореактором, содержавшим в растворе полинуклеотиды и полипептиды, а также другие вещества. Этот реактор был отделен от окружающей среды мембраной и одновременно связан с окружающей средой путем обмена водой и растворенными в ней веществами.
Протоклетки обладали способностью к самовоспроизводству с небольшими ошибками, конкурировали друг с другом и были подвержены давлению отбора. В силу уникальных физико-химических свойств полинуклеотидов протоклетки были наделены способностью к хранению и передаче информации, закодированной в последовательности оснований. Новые протоклетки возникали в результате деления клеток. Дочерние клетки после непродолжительной физико-химической фазы роста становились неотличимыми от материнских клеток по всем свойствам, дочерние клетки не претерпевали индивидуального развития и по крайней мере в принципе были бессмертны. Многие из ныне, живущих простейших микроорганизмов, например, вирусы и бактерии, лишены индивидуального развития, они "рождаются в готовом виде" и живут столько, сколько позволяют условия окружающей среды. У более сложных одноклеточных организмов, например, у дрожжевых клеток, наблюдаются отчетливо выраженные жизненные циклы с фазами юности, зрелости, размножения и старости. С возникновением многоклеточных организмов индивидуальное развитие занимает все более важное место.
Наиболее ярким примером может служить человек. Между грудным ребенком и зрелым взрослым человеком имеются существенные качественные различия не только с точки зрения морфологии. Младенец и взрослый отличаются и по своей работоспособности, и по своим интеллектуальным возможностям.
На ранней фазе эволюции живого первые многоклеточные организмы появились лишь как результат случайного образования кластеров. По своим свойствам они еще несущественно отличались от одноклеточных организмов. Индивидуальное развитие как новая стратегия складывалось лишь постепенно, и последующий успех этой стратегии опирался на ее значительные эволюционные преимущества. Онтогенез, характеризуемый как процесс индивидуального развития, является большим достижением эволюции, типичным лишь для более поздних стадий филогенеза – исторического развития таксономических групп. Онтогенез – непременная предпосылка не только образования более сложных морфологических структур, но и процессов обучения, на которых основывается работоспособность всех высших организмов. Поскольку ген вследствие ограниченной емкости его "памяти" может передавать только информацию, важную для выживания, основной поток информации осуществляется через процессы обучения. Возможность обучения является важным преимуществом при отборе. Процессы обучения открывают совершенно новые возможности обработки информации. Поскольку онтогенез занимает в эволюции центральное положение, его эволюционное моделирование имеет большое значение.
2. Дискретные модели циклов жизни
Важные свойства систем, элементы которых претерпевают индивидуальное развитие, могут быть изучены даже на простых дискретных моделях. В простейшем случае жизненный цикл состоит только из двух стадий – молодость и зрелость, которые мы обозначим номерами 1 и 2.
Рассмотрим в качестве примера популяцию клеток прокариотов, например, бактерий. Индивидуальное развитие клеток прокариотов можно разделить на две стадии.
1) Молодость клетки; на этой стадии, продолжающейся около 40 минут, делится ДНК клетки.
2) Зрелость клетки.
Развитие популяций эукариотов, например, дрожжевых клеток, может быть разделено на следующие фазы.
1) Молодость клетки: клетка сосредоточивается на синтезе белка.
2) Зрелость клетки: синтез ДНК; после различных дальнейших этапов, наконец, наступает
3) Митоз.
Если ограничиться двумя стадиями, то процесс моделируется следующими уравнениями;
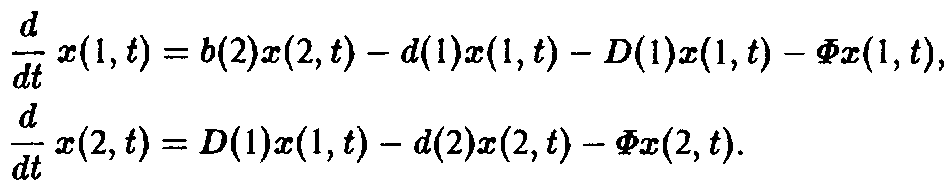
Здесь 6 характеризует размножение, а также порождение молодых клеток зрелыми клетками, d – смертность и D – переход молодость–зрелость.
Величина Ф означает разбавление потока для создания давления отбора. В простейшем случае неумирающих клеток имеем:
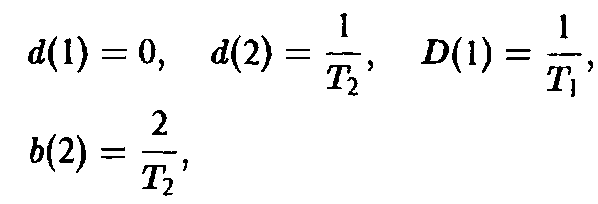
где Г, – продолжительность i-й фазы.
Скорости размножения могут в определенных пределах замедляться. Такое торможение регулируется производством чалонов – ингибиторов клеточного деления. Это приводит к зависимости скорости роста от концентрации. Если ингибиторы испускаются только на стадии зрелости, то скорость роста можно описать следующим выражением:
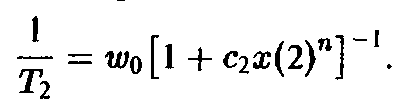
В этом случае в системе существует и колебательный режим. Если же ингибиторы производятся главным образом на стадии молодости клетки, то скорость роста описывается следующим выражением:
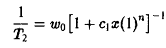
где п – надлежащим образом выбранный показатель.
Конкуренция и отбор могут быть введены следующим образом. Рассмотрим систему, состоящую из различных сортов, в две стадии:
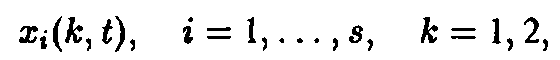
и введем конкуренцию с помощью зависящего от времени разбавления потока:
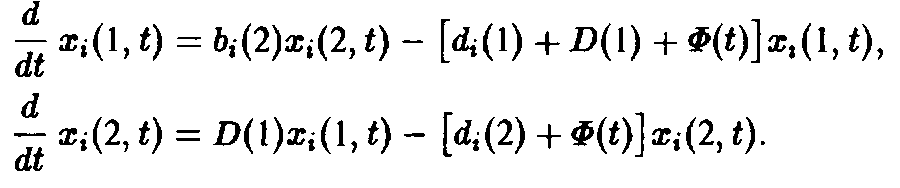
При условии постоянного общего числа частиц получаем:
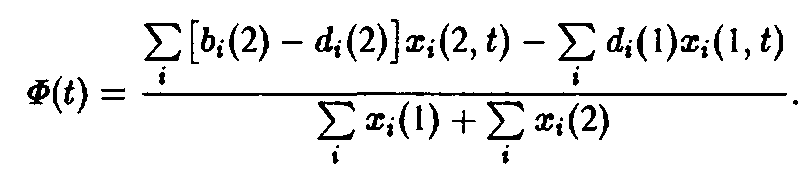
Вторая возможность состоит в том, чтобы создать давление конкуренции посредством связи через субстрат, т.е.
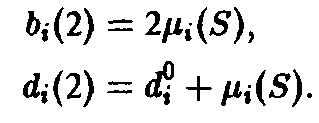
Здесь
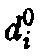 – естественная смертность зрелых
клеток,
– естественная смертность зрелых
клеток,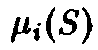 – скорость деления в зависимости от
концентрации субстрата. Для описания
расхода субстрата используется
дополнительное уравнение:
– скорость деления в зависимости от
концентрации субстрата. Для описания
расхода субстрата используется
дополнительное уравнение:
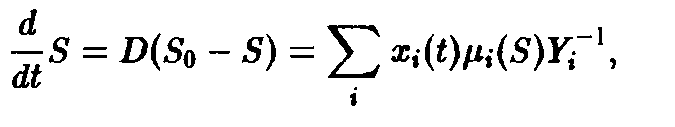
где Y-1>i> – мера эффективности использования субстрата.
Более подробное обсуждение моделей с двумя и более стадиями индивидуального развития, а также их приложений можно найти в литературе.
3. Модель старения Маккендрика фон Фёрстера
Для математического описания индивидуального развития Маккендрик и фон Фёрстер разработали рекуррентную модель, в которой непрерывный процесс старения рассматривается как последовательность отдельных, изолированных, актов. С математической точки зрения речь идет о том, чтобы вывести систему дифференциальных уравнений для функции плотности x, описывающей, сколько особей возраста г существуют в системе в момент времени t. Рассмотрим такую систему
в момент времени t + At. Число особей, которые к этому времени достигнут возраста г, равно числу особей, которые к моменту времени t достигли возраста г – At, за вычетом особей, умерших за интервал времени At. В результате мы получаем:
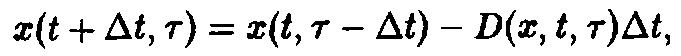
где D – смертность. Совершая предельный переход при At – О, приходим к дифференциальному уравнению
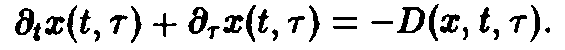
Важную роль в динамике системы играют также процессы воспроизводства, характеризуемые рождаемостью В. Эта величина показывает, сколько потомков производят в момент времени t особи, достигшие возраста т. В качестве начального условия по г для уравнения мы получаем величину
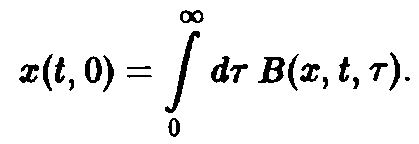
Оно дополняется заданием второго начального условия
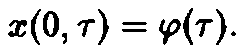
Уравнения, с начальными условиями, задают модель Маккендрика–фон Фёрстера для популяций с возрастной структурой. Эта модель позволяет определить временную эволюцию функции плотности x по заданному начальному распределению у, если известны скорость воспроизведения и смертность. Решения, получаемые в рамках этой модели при различных конкретных функциях D и В, подробно изучены (McKendrick, 1926; von Foerster, 1959; Полу-эктов, 1974; Полуэктов и др., 1980; Романовский и др., 1984). Смертность D и рождаемость В, вообще говоря, сложным образом зависят от плотности x и), что приводит к существованию разнообразных структур решений. Однако уравнения в этом случае также становятся весьма сложными, и их аналитическое решение становится невозможным. Поэтому мы сначала рассмотрим простейший случай, когда рождаемость и смертность линейны по плотности x и не зависят явно от времени:
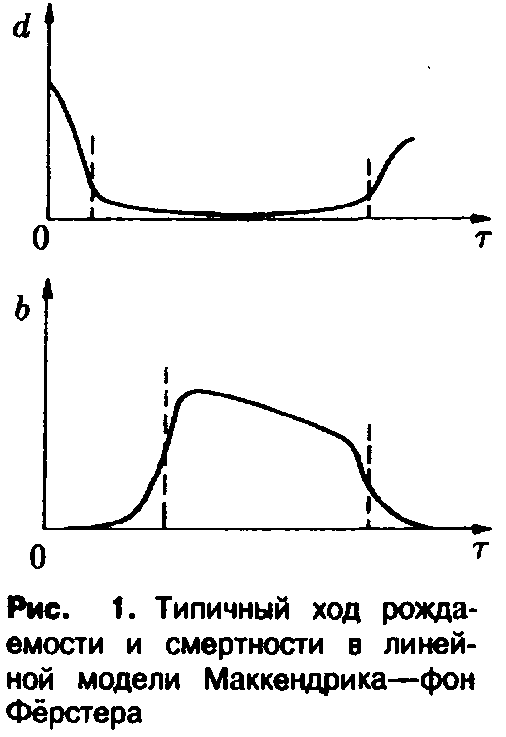
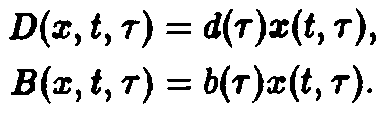
Реалистические
примеры функций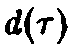 и
и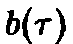 представлены на рис. 1. Смертность по
прошествии первого времени после
рождения стабилизируется на относительно
низком уровне и начинает снова возрастать
лишь при больших г; в дальнейшем мы,
кроме того, принимаем предположение
представлены на рис. 1. Смертность по
прошествии первого времени после
рождения стабилизируется на относительно
низком уровне и начинает снова возрастать
лишь при больших г; в дальнейшем мы,
кроме того, принимаем предположение

Рождаемость, как правило, достигает своего максимального значения лишь по истечении определенного периода после начала фазы зрелости и в старости снова понижается.
Учитывая соотношения, мы получаем из уравнений – систему уравнений
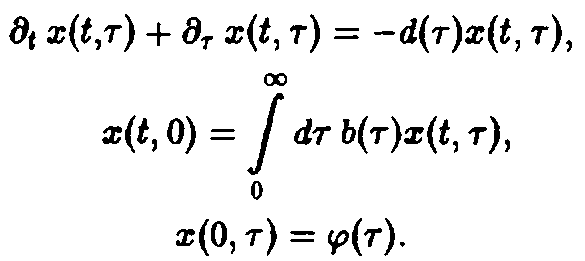
Поведение решений системы уравнений вполне очевидно. Из первого уравнения мы получаем, прежде всего, вероятность W того, что особь достигает возраста т:
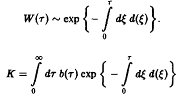
В зависимости от величины К возможны три качественно различных случая. При К > 1 число новорожденных в единицу времени больше числа умерших, доминируют процессы воспроизводства, и при t → оо мы получаем при всех г расходящуюся плотность x – оо. Наоборот, при К<1 воспроизводство слишком слабо, и при t – оо мы получаем x –* О при всех т, т.е. вид вымирает. Наконец, при К = I оба процесса находятся в равновесии, соответственно, существует бесконечно много стационарных состояний, и только от начального условия <р зависит, какое из них реализуется. Разумеется, в случаях К > 1 и К < 1 результат не зависит от начального условия <р.
Тем самым мы получаем качественную характеристику динамики индивидуального развития внутри отдельного вида при упрощающем предположении. Исследуем теперь, какие модификации возникают в том случае, когда п видов развиваются в соответствии с уравнениями, аналогичными уравнению, и, кроме того, взаимодействуют между собой посредством процесса отбора. Затронутая проблема связана с вопросом оптимальной стратегии старения, сложившимся в ходе эволюции.
4. Процессы отбора в моделях с непрерывным старением
Прежде всего систему п не взаимодействующих между собой видов можно описать уравнениями, обобщающими уравнение:
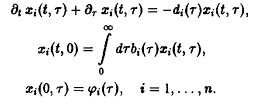
В качестве простого метода создания давления отбора мы по аналогии с моделью Эйгена потребуем постоянства общего числа особей в системе:
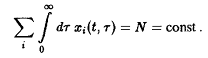
Чтобы условие выполнялось, необходимо модифицировать систему уравнений 13, что можно осуществить различными способами. Особый интерес представляют две возможности.
1. Введение потоковых членов в модель Эйгена.
Такая операция соответствует подстановкам

в уравнение, причем во избежание патологии, например, отрицательных концентраций, должны выполняться неравенства
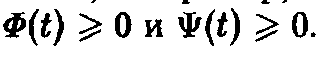
2. Регуляция скорости воспроизведения.
Регуляция достигается с помощью подстановки
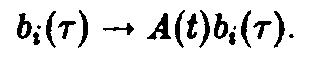
И в том, и в другом случае существенно, что модификации либо видо-, либо возрастоспециф ич ны.
Если равенство продифференцировать по времени и воспользоваться уравнением с подстановками, то получится следующее:
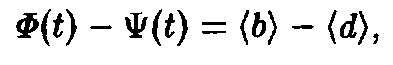
где по определению
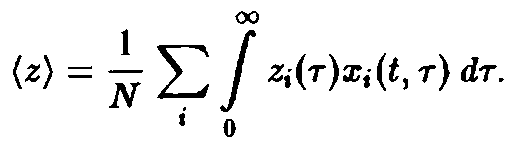
Учитывая положительность
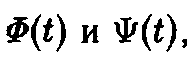
мы получаем

и, наконец, приходим к системе уравнений

Проводя аналогичные вычисления с использованием подстановок, получаем, полагая
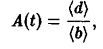
систему уравнений
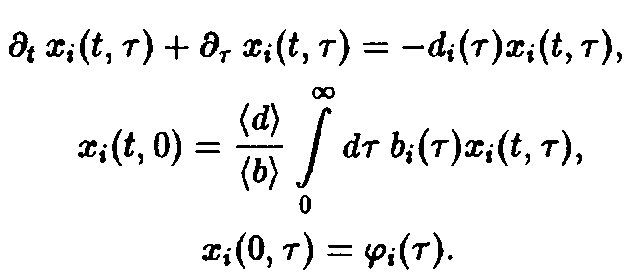
Уравнения и описывают временную эволюцию систем стареющих конкурирующих между собой видов и тем самым удобны для математического анализа индивидуального развития и отбора.
В отличие от системы уравнений для независимых видов дифференциальные уравнения и связаны между собой через определенное соотношением среднее значение. С одной стороны, эта связь выступает как математическое выражение взаимодействия между видами, а с другой – исключает возможность получения аналитических решений и обусловливает тем самым весьма широкое применение численных методов.
Ряд интересных утверждений может быть высказан и без явного решения системы уравнений. В частности, необходимо выяснить, каким образом, зная функции d, и Ь, можно определить те виды, которые замещают другие и поэтому доминируют при больших временах.
Необходимый для этого качественный анализ динамики удается осуществить с помощью подстановок
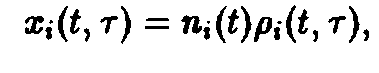
где
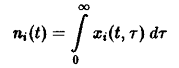
– общее число частиц,
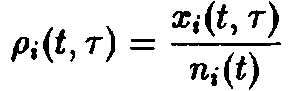
– нормированная возрастная структура i-ro рода. Рассмотрим сначала ситуацию, описываемую уравнением. Пользуясь подстановкой, получаем следующие уравнения для п, и pi:
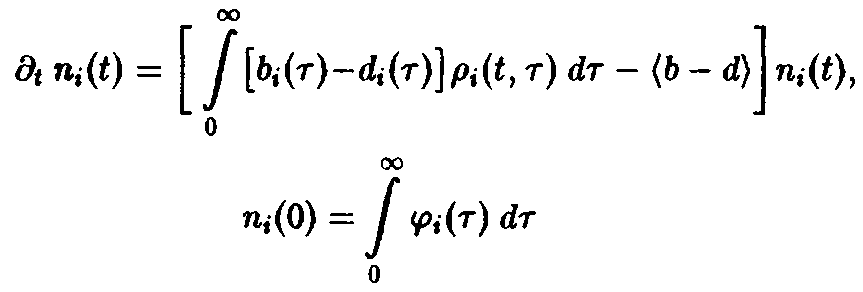
и
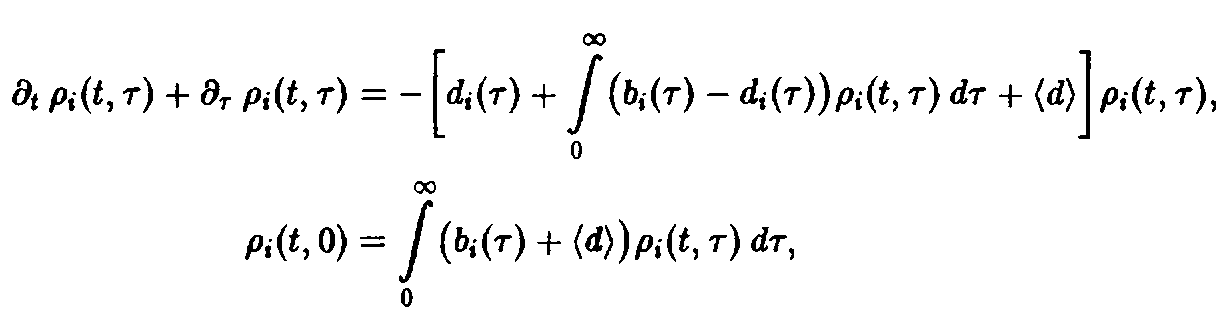
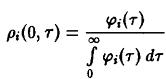
Уравнение имеет в точности такую же структуру, как уравнение Эйгена, с тем лишь различием, что теперь приспособленность
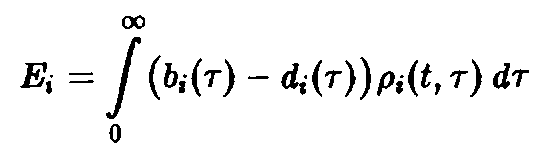
– функционал
нормированной возрастной структуры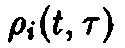 и
поэтому может изменяться
во времени.
и
поэтому может изменяться
во времени.
Его
временная эволюция определяется
изменением во времени возрастное
структуры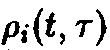 ,
которая в свою очередь зависит от
динамики чисел через уравнение.
Зависимость приспособленности
нормированной возрастной структуры
приводит к тому, что виды могут повысить
свои шансы на успех в ходе отбора за
счет
подходящего
распределения особей по возрастным
группам; иначе говоря, в ходе эволюции
происходит замещение одних видов другими
с оптимальной возрастное структурой.
Каким образом по заданным функциям
,
которая в свою очередь зависит от
динамики чисел через уравнение.
Зависимость приспособленности
нормированной возрастной структуры
приводит к тому, что виды могут повысить
свои шансы на успех в ходе отбора за
счет
подходящего
распределения особей по возрастным
группам; иначе говоря, в ходе эволюции
происходит замещение одних видов другими
с оптимальной возрастное структурой.
Каким образом по заданным функциям
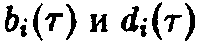 >
>можно
определить какие виды выживут в конце
концов?
Чтобы ответить на этот вопрос, рассмотри
стационарные решения уравнений и при
больших временах
>
>можно
определить какие виды выживут в конце
концов?
Чтобы ответить на этот вопрос, рассмотри
стационарные решения уравнений и при
больших временах
Мы получаем
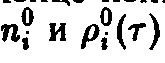
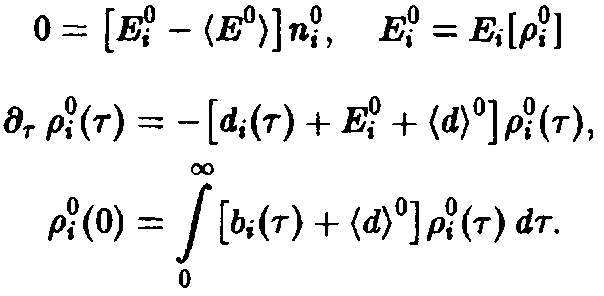
При рассмотрении уравнения Эйгена мы обнаружили, что уравнение допускает п различных стационарных решений вида

т.е. стационарны только такие ситуации, в которых все N особей представлены одним видом. С учетом соотношения из формулы следует, что
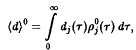
поэтому
уравнение для
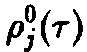 >
>замыкается.
Таким
образом, мы получаем:
>
>замыкается.
Таким
образом, мы получаем:
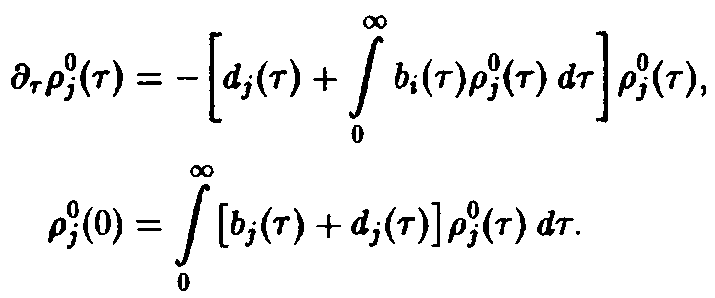
Вводя сокращенные обозначения
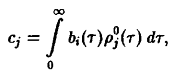
запишем уравнение в виде
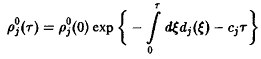
и далее, с помощью соотношения,
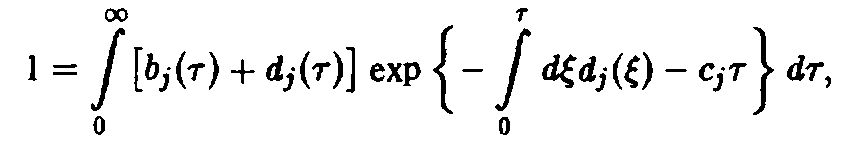
а также

вследствие того, что по определению р,
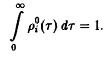
Тем
самым мы полностью охарактеризовали
стационарные решения систем уравнений.
Можно показать, что при заданных
 >
>и
>
>и
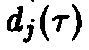 >
>уравнение
всегда допускает ровно одно решение
>
>уравнение
всегда допускает ровно одно решение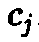
Величины
Cj
в
силу соотношения определяют,
поэтому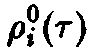 однозначно
определяются соотношением.
однозначно
определяются соотношением.
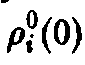
Из стационарных решений устойчиво только одно, и при t –* оо именно оно описывает поведение системы. Для этого решения справедливы неравенства
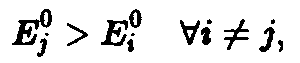
т.е. выживает вид, обладающий наибольшей приспособленностью. Соотношения – позволяют определить этот вид посредством формулы
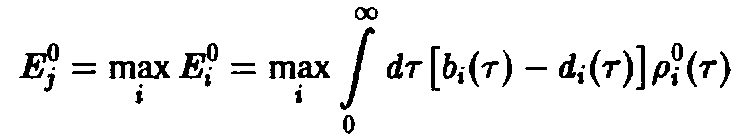
по
известным функциям
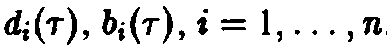 .
Тем самым для системы конкурирующих
видов с возрастной структурой, описываемой
уравнением, становится возможным
определять на основе заданных зависящих
от возраста скорости воспроизведения
и смертности тот из видов, который
побеждает в ходе отбора. Используя
соотношения, получаем
.
Тем самым для системы конкурирующих
видов с возрастной структурой, описываемой
уравнением, становится возможным
определять на основе заданных зависящих
от возраста скорости воспроизведения
и смертности тот из видов, который
побеждает в ходе отбора. Используя
соотношения, получаем
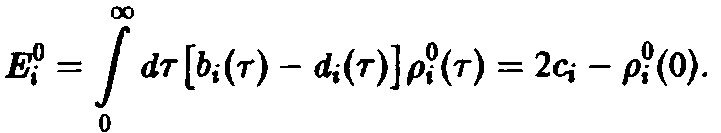
Можно показать, что
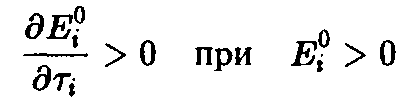
Таким
образом, если мы ограничимся видами с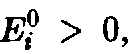 >
>т.е.
такими, которые в отсутствие отбора
сами не вымирают и то максимум величины
>
>т.е.
такими, которые в отсутствие отбора
сами не вымирают и то максимум величины
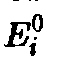 >
>достигается
для того вида j,
для
которого величина
>
>достигается
для того вида j,
для
которого величина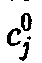 также максимальна. Следовательно,
доминирующий вид может определяться
вместо соотношения соотношением
также максимальна. Следовательно,
доминирующий вид может определяться
вместо соотношения соотношением
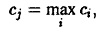
где с, – определяется формулой. Аналогичные соображения применимы и к модели (Эбелинг и др., 1986). Следует отметить, что в рассматриваемом случае соотношение получается и как определяющее уравнение для выживающего вида. Если воспользоваться определением параметров с, то соотношение можно рассматривать как разумное обобщение соотношения.
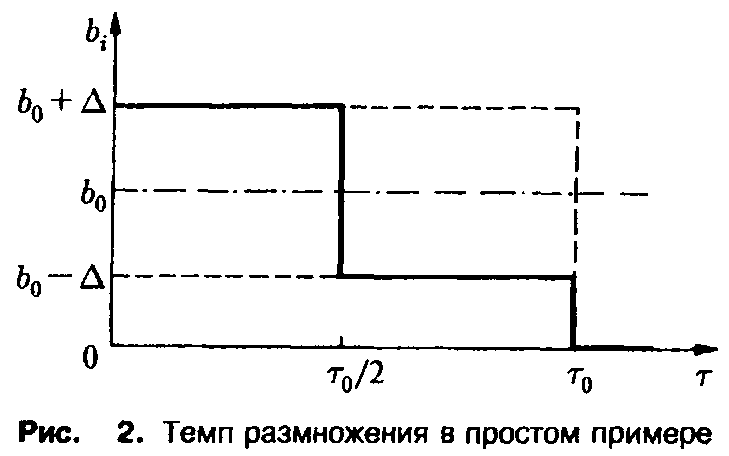
В заключение продемонстрируем важность внутривидовой возрастной структуры для процесса отбора на простом примере динамики, описываемой уравнением. Рассмотрим два вида с одинаковой и постоянной смертностью
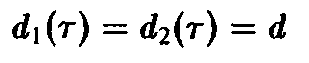
и рождаемостью
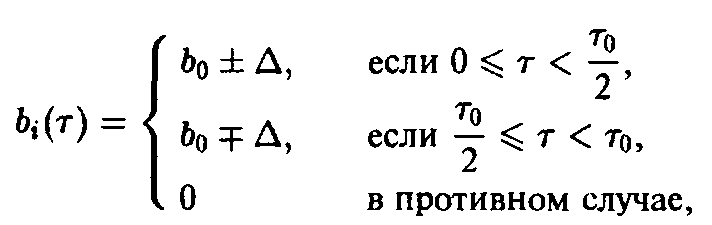
где в и в последующих соотношениях верхние знаки относятся к первому виду, а нижние – ко второму виду. Выбранные рождаемости представлены на рис. 2. Чтобы выяснить, какой вид побеждает в процессе отбора, необходимо исследовать, какая из определяемых соотношением величин с, больше. В рассматриваемом случае равенство упрощается до
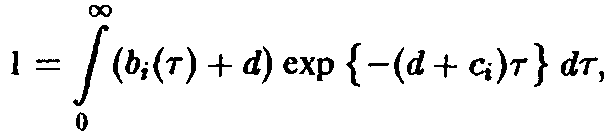
или
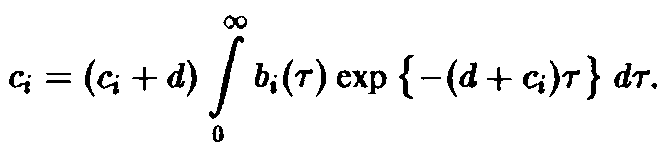
Отсюда с учетом формулы получаем
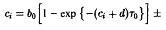
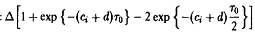
Подставляя
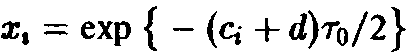 находим
находим
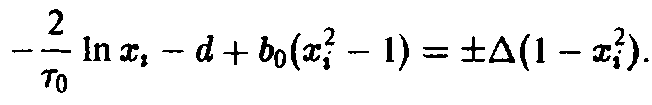
Графическое решение этого уравнения представлено на рис. 3. Из хода кривой мы заключаем, что Х < х>2> вследствие монотонности подстановки приводит к неравенству с > С2 – Таким образом, первый вид выживает, а второй вымирает; грубо говоря, те преимущества, которыми второй вид обладает в старости, не перевешивают его недостатков в юности. Этот пример наглядно демонстрирует упрощенное описание процесса отбора с учетом возрастной структуры. В случае динамики Эй-гена оба вида характеризовались бы усредненной приспособленностью, в результате чего никакого отбора не происходило бы, и оба вида могли бы сосуществовать.
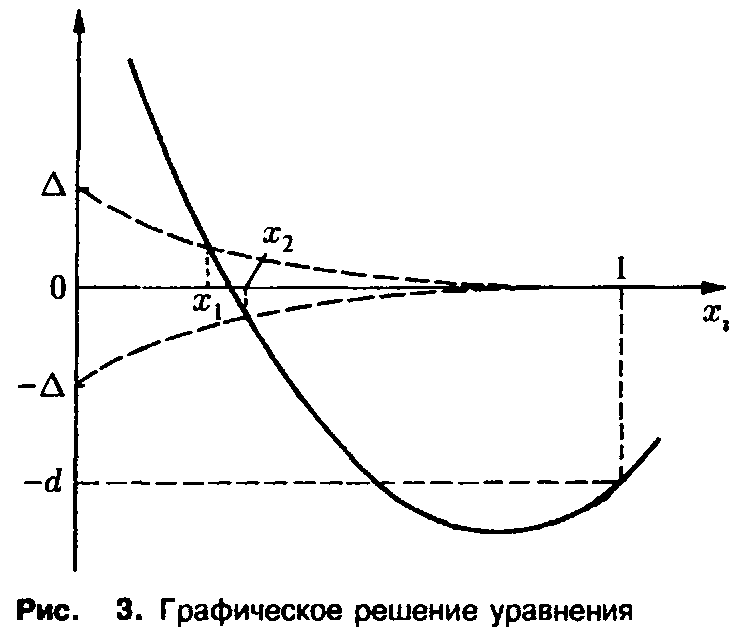
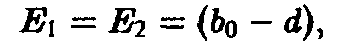
Действительно, при одинаковых начальных условиях
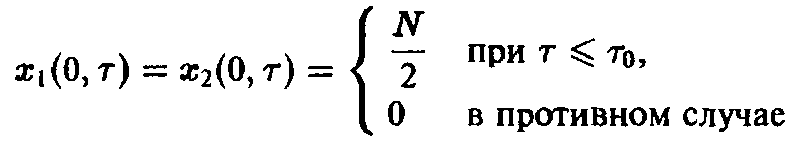
мы получаем из соотношения
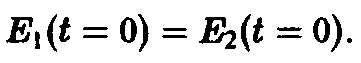
Однако
динамика внутривидовой возрастной
структуры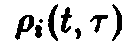 приводит
к временному изменению приспособленности:
приводит
к временному изменению приспособленности:
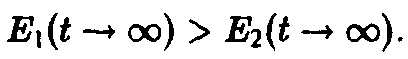
Тем
самым даже в простейших моделях
индивидуальное развитие внутри видов
имеет решающее значение для исхода
протекающих
процессов отбора. Другие простые примеры
для функций
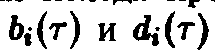 показывают, что обычно высокая скорость
воспроизведения и низкая смертность в
сравнительно молодом возрасте является
преимуществом в борьбе за отбор. Это
обстоятельство тесно связано с тем, что
стационарные возрастные структуры
показывают, что обычно высокая скорость
воспроизведения и низкая смертность в
сравнительно молодом возрасте является
преимуществом в борьбе за отбор. Это
обстоятельство тесно связано с тем, что
стационарные возрастные структуры монотонно
затухают по
монотонно
затухают по .
.
5. Сложные возрастные структуры
Можно указать несколько случаев, когда более сложные и, следовательно, более реалистические возрастные структуры удается описать с помощью модели Маккендрика фон Фёрстера и ее обобщения. Мы не будем предпринимать попыток решить соответствующие уравнения, а ограничимся изложением возможностей, присущих формализму.
Прежде всего, напомним наиболее общую форму – модели в случае одного отдельного вида:
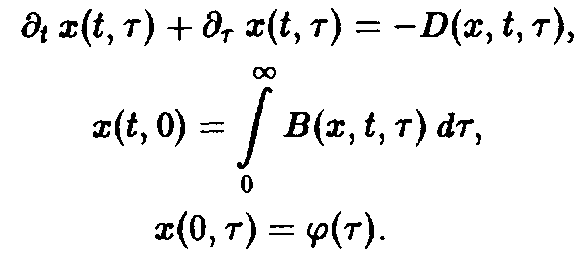
Множество самых различных решений определяется выбором функций D (x, t, т) и В. В дальнейшем мы обсудим лишь несколько принципиальных вариантов. Соображения, развиваемые ниже относительно функции D, могут быть по аналогии перенесены на рождаемость В.
Прежде всего, мы предположим существование явной зависимости от времени. Это позволит учитывать изменения внешних условий. Колебания могли бы моделировать годичные или более длительные изменения, чередования теплых и холодных периодов, а скачкообразные изменения – влияние природных катаклизмов на экосистемы. Существуют и другие разнообразные условия, приводящие как к положительным, так и к отрицательным последствиям, но их вряд ли уместно классифицировать более подробно.
В дальнейшем мы всегда будем предполагать, что система всегда находится в стационарном окружающем поле. Явная зависимость от времени в этом случае не возникает, но изменение величины D в зависимости от плотности х может быть весьма разнообразным. Рассмотрим сначала простейший случай – зависимость от х,

Несколько более реалистическим является учет ограничивающего члена в виде
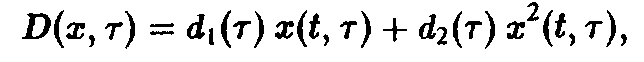
препятствующем
для К
> 1)
расходимости
 >
>при
>
>при
 >
>Если
число особей очень велико, то смертность
сильно возрастает и система стабилизируется
на уровне, зависящем от функций
>
>Если
число особей очень велико, то смертность
сильно возрастает и система стабилизируется
на уровне, зависящем от функций
 i.
В
i.
В
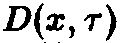 >
>могут
быть включены члены более высокого)
порядка. Включение кубической нелинейности
может привести к возникновению
бистабильных ситуаций, в которых число
особей
>
>могут
быть включены члены более высокого)
порядка. Включение кубической нелинейности
может привести к возникновению
бистабильных ситуаций, в которых число
особей
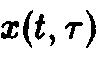 >
>может
устанавливаться на двух, вообще говоря,
не зависящих от г устойчивых значениях.
>
>может
устанавливаться на двух, вообще говоря,
не зависящих от г устойчивых значениях.
Однако возможные режимы этим отнюдь не исчерпываются. Особенно большой интерес наряду с перечисленными выше вариантами нелокальных зависимостей представляют такие, которые в простейшей форме могут быть записаны следующим образом:

Ядро
интеграла
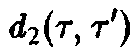 >
>описывает
взаимодействие между особями самых
различных возрастов внутри одного вида.
При
>
>описывает
взаимодействие между особями самых
различных возрастов внутри одного вида.
При
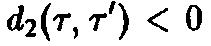 >
>эффективная
смертность особей, достигших возраста
г, понижается за счет наличия особей
возраста
>
>эффективная
смертность особей, достигших возраста
г, понижается за счет наличия особей
возраста
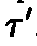 .
Примером может служить, например,
особенно тщательная забота о потомстве
на протяжении первых отрезков жизни
или поддержка старых, весьма опытных
особей. Соответственно,
.
Примером может служить, например,
особенно тщательная забота о потомстве
на протяжении первых отрезков жизни
или поддержка старых, весьма опытных
особей. Соответственно,
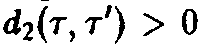 >
>приводит
к внутривидовой конкуренции, проявляющейся
в борьбе за верховенство или в "конфликтах
поколений". Как и в соотношение, в
формулу могут
входить члены высшего порядка, описывающие
взаимодействие между тремя и более
различными возрастными группами.
>
>приводит
к внутривидовой конкуренции, проявляющейся
в борьбе за верховенство или в "конфликтах
поколений". Как и в соотношение, в
формулу могут
входить члены высшего порядка, описывающие
взаимодействие между тремя и более
различными возрастными группами.
Введение давления отбора с помощью условия постоянства организации в целом позволяет моделировать только простейшую форму межвидовой конкуренции. Взаимодействие хищник-жертва можно рассматривать как обобщение взаимодействия вида.