Эволюция адаптивных стратегий
Эволюция адаптивных стратегий
Введение
Биологи-эволюционисты пытаются объяснить, каким образом путем естественного отбора жизнь достигла той степени развития, которую мы наблюдаем сегодня. Чтобы объяснить существование конкретного генетического признака, они высказывают предположения по поводу того, что было до появления этого признака. Затем постулируют, что какой-то редкий ген появился у отдельной особи или что такой же ген был привнесен иммигрантом и что особи, несущие этот ген, стали обладателями данного признака. Затем они задаются вопросом, какие условия среды будут благоприятствовать распространению этого гена в популяции. Если естественный отбор благоприятствует этому гену, то у особей, генотип которых содержит упомянутый ген, приспособленность увеличится. Можно сказать, что этот ген вторгся в популяцию. В этой главе мы рассмотрим, как аргументы такого рода можно использовать при изучении поведения животных.
1. Эволюционные стратегии
Чтобы сохраниться в популяции, ген не только должен выдержать конкуренцию с другими генами генофонда, но и выдержать вторжение других мутантных генов. Действительно эволюционные биологи, рассуждая о конкретной эволюционной ситуации, часто постулируют вторжение гипотетических мутантов, чтобы выяснить, устоит ли данная теория при наличии конкуренции. Перед тем как вникать в детали этой процедуры, рассмотрим несколько примеров. Высокая степень соответствия животного его окружению может быть результатом отбора по признакам, которые позволяют животному избегать хищников или хищникам оставаться незамеченными и нападать из засады на приближающуюся жертву. Этот тип стратегии иногда принимает форму удивительного сходства между животным и растением, с которым оно обычно связано. Например, напоминающий лист кузнечик Arantia rectifolia может сидеть неподвижно с плотно сжатыми крыльями, имитируя лист, или, сидя на листе, может держать крылья расправленными и казаться частью листа. Эти маскировочные позы считаются защитой против хищников. Наоборот, богомол Phyllocrania paradoxa может замаскироваться точно так же, но его маскировка будет частично защитной и частично агрессивной, поскольку он сидит неподвижно, поджидая свою жертву, чтобы схватить ее.
Помимо сходства с растениями или субстратом животные также могут иметь сходство с отдельными частями других животных. Часто это проявляется в виде особых демонстраций, которыми животные пытаются отпугнуть своих врагов. Например, гусеница южноамериканского бражника Leucorampha обычно держится спиной вниз под веткой или листом. Потревоженная, она поднимает и раздувает голову, на нижней поверхности которой хорошо видны пятна, похожие на глаза, а общим видом гусеница напоминает голову змеи. У многих ночных и дневных бабочек на крыльях есть пятна, напоминающие глаза; если бабочку потревожить, она внезапно раскрывает крылья, демонстрируя пятна-глаза, и хищник обычно пугается.
Истинная мимикрия - это сходство одного животного, называемого имитатором, с другим, называемым моделью, так что их обоих путает третье животное, обычно хищник. Существуют разные формы мимикрии, из которых наиболее иллюстративна бейтсовская мимикрия. При этой форме мимикрии животное-имитатор напоминает модель, которая ядовита или неприятна на вкус для хищника. Для животного-модели такая мимикрия бесполезна, но, судя по экспериментальным данным, имитаторы в конечном счете получают преимущество, поскольку хищники не отличают имитатора от модели. Например, птицы быстро научаются избегать саламандры Notophthalmus viridescens, мясо которой неприятно на вкус. Однако они охотно поедают сходно окрашенную саламандру Pseudotriton ruber при условии, что они прежде не сталкивались с несъедобной Notophthalmus. Таким образом, Pseudotriton извлекает некоторую пользу из своего сходства с Notophthalmus при условии, что птицы уже знают вид последней и что они не так опытны, чтобы суметь различить два этих вида саламандр. Некоторые другие примеры бейтсовской мимикрии показаны на рис. 1.
Иногда ряд ядовитых видов имеет одинаковую предупреждающую окраску. Эта форма мимикрии называется мюллеровской мимикрией; она дает преимущество всем видам-участникам, поскольку, когда хищник научается избегать один вид, он будет избегать и все виды-имитаторы. На рис. 3 показаны некоторые примеры мюллеровской мимикрии.
Замечательный случай агрессивной мимикрии обнаружен у рыб. Губан-чистильщик, окрашенный в черно-белую полоску, живет среди коралловых рифов в Тихом океане. Другие рыбы узнают его по заметной окраске и привлекающему внимание поведению. Крупные рыбы позволяют губану приближаться и удалять паразитов с поверхности тела и из полости рта. Эти отношения носят характер симбиоза, поскольку губан извлекает пользу, получая пищу, а рыба-хозяин избавляется от паразитов. Однако у губана-чистильщика есть имитатор, весьма сильно его напоминающий, - саблезубая морская собачка. Другие рыбы часто по ошибке принимают ее за губана-чистильщика и позволяют приблизиться. Тогда саблезубая морская собачка быстро откусывает кусок плавника у рыбы-хозяина и поспешно уплывает.
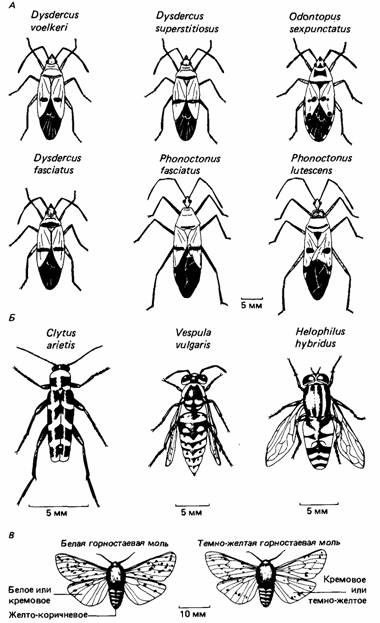
Рис. 1. Некоторые примеры мюллеровской и бейтсовской мимикрии.
А. Мюллеровская мимикрия у западноафриканских Heteroptera. Клоп-хищнец Phonoctonus fasciatus обычно живет в колониях Dysdercus fasciatus, D. superstitiosus и D. voelkeri. Phonoctonus lutescens обычно живет в колониях Odontopus, иногда также с D. voelkeri. Все эти клопы имеют в своей окраске черный, красный и серовато-оранжевый цвет.
Б. Бейтсовская мимикрия. Оса Vespula vuIgaris и два имитатора: жужелица Clytus arietis и Helophilus hybridus. У всех трех тело темно-коричневое или черное, с желтыми полосками.
В. Неприятная на вкус белая горностаевая моль и ее бейтсовский имитатор - темно-желтая горностаевая моль. Брюшко у обеих желтоватое, с темными отметинами, тогда как крылья белые или кремовые, с черными пятнами.
В случае бейтсовской мимикрии предупреждающая хищника окраска одного вида-модели имитируется другим видом-имитатором. Это будет выгодно в том случае, если хищники не в состоянии отличить имитатора от модели и если животные-имитаторы не будут встречаться слишком часто по сравнению с моделями. Если модель неприятна на вкус для хищника, то имитатор выигрывает, поскольку хищники, по-видимому, будут избегать как модели, так и имитатора. При этом животному-имитатору не нужно нести расходы на выработку отпугивающих веществ. В некоторых случаях одной модели могут подражать несколько видов, и тогда существует опасность, что хищники будут выбирать больше животных-моделей, чем могли бы. Например, большая и бросающаяся в глаза бабочка данаида Danaus plexippus, обитающая на западе США, неприятна на вкус для птиц, и они научаются избегать ее после одной или двух проб. Гусеницы данаиды питаются разными видами молочаев, которые ядовиты для большинства позвоночных. Гусеницы накапливают токсины, которые сохраняются в куколке и, таким образом, остаются в бабочке. У хищников, съевших эту бабочку, обычно наблюдается рвота. Вызывающую рвоту данаиду имитирует ленточник Limenitis archippus. Ленточники съедобны, и, по некоторым данным, сойки с удовольствием поедают их, если предварительно не сталкивались с данаидами. А съев один раз данаиду, сойки впредь избегают и данаид, и ленточников. У данаиды есть и другие имитаторы, поэтому не исключено, что в какой-либо области число имитаторов может превысить число моделей. В такой ситуации хищники не скоро научаются избегать данаид и их имитаторов, поскольку их первым опытом может быть встреча со съедобными бабочками; таким образом, уровень поедания хищниками как модели, так и имитатора, вероятно, будет выше.
Пытаясь проследить эволюцию такого состояния, биолог должен постараться рассчитать последствия возможных комбинаций частот различных генотипов в популяциях модели и имитатора. Например, что может случиться, если некоторые данаиды перестанут питаться на ядовитых растениях и станут съедобными для хищников? С одной стороны, можно ожидать, что такие особи получают преимущество, оставаясь совершенно похожими на съедобных особей своего вида и при этом не неся затрат на выработку устойчивости к поеданию ядовитых молочаев; это дает основание для увеличения частоты такого генотипа.
С другой стороны, если бы съедобные данаиды стали слишком обычны, хищники начали бы активно поедать их. На основе некоторых допущений можно подсчитать вероятность выживания съедобных данаид как функцию соотношения съедобной и несъедобной форм в популяции. Допущения касаются степени хищничества и числа попыток, которые требуются хищнику, чтобы начать избегать данаиду после поедания особи, вызывающей рвоту. Фактически содержание ядовитых веществ в молочаях сильно различается, как различается и съедобность данаид, гусеницы которых питались на разных молочаях. Таким образом, в естественных популяциях все же имеется некоторое число данаид, поедающих малоядовитые виды молочаев и съедобных для птиц; эта форма мимикрии называется автомимикрией. В некоторых местностях отношение автоимитаторов и модели составляет 3:1, и все же хищники избегают их поедать.
Если переход к нетоксичной диете имеет генетическую основу, а не просто случаен, то такое изменение можно считать эволюционной стратегией. Таким образом, эволюционным биологам иногда полезно считать, что гены применяют какую-либо стратегию, чтобы увеличить свою частоту за счет других генов, даже если в действительности вся эта процедура проводится пассивно естественным отбором.
Рассматривая эволюционные стратегии, которые влияют на поведение, мы должны отчетливо представить себе ситуацию, при которой изменения в генотипе ведут к изменениям в поведении. Совсем необязательно считать, что поведение непосредственно контролируется генами, поскольку существует много способов, посредством которых изменения в генетическом аппарате могут влиять на поведение. Однако лучше связывать поведение с генами, понимая, что это делается просто для удобства. Так, говоря о «гене заботы о родственниках», мы предполагаем, что в популяции существуют генотипические различия, которые заставляют одних особей помогать своим родственникам, а других - не помогать. Точно так же под «стратегией голубя» мы подразумеваем поведение в популяции особей, которые не вступают в конфликт и передают этот признак от одного поколения другому.
На первый взгляд может показаться, что наиболее успешная эволюционная стратегия распространится во всей популяции и в конце концов вытеснит все остальные. Иногда это случается, но далеко не всегда. Во многих ситуациях невозможно определить, какая стратегия является наилучшей, поскольку эффективность любой стратегии зависит от поведения других животных, т.е. конкурирующие стратегии зависят друг от друга так, что успех одной зависит от существования другой и от частоты, с которой эта другая представлена в популяции. Например, стратегия, использующая мимикрию, не имеет никакой ценности, если неэффективна стратегия отпугивания моделей.
У некоторых видов есть совместные территории для спаривания, называемые токовищами, на которых доминантные самцы защищают небольшие участки. Например, у турухтанов обнаружены два типа самцов: территориальные самцы, которые агрессивны и соперничают друг с другом из-за самок, и самцы-сателлиты, которые не агрессивны, держатся около территориальных самцов и спариваются с самками украдкой, пока территориальный самец занят чем-нибудь другим. У самцов-сателлитов имеется белый бросающийся в глаза воротник, тогда как у территориальных самцов окраска темнее. Полагают, что территориальные самцы терпят присутствие самцов-сателлитов потому, что их бросающееся в глаза оперение привлекает самок к токовищу. Существуют две точки зрения по поводу происхождения самцов-сателлитов. Во-первых, это могут быть самцы, потерпевшие неудачу в конкуренции за территорию, или молодые и неопытные самцы, которые принимают стратегию сателлитов, чтобы добиться максимально возможного в своем положении и дождаться, когда участок станет свободным. Считают, что нечто подобное происходит на токовищах у тетерева. Во-вторых, полагают, что самцы-сателлиты применяют альтернативную эволюционную стратегию и их успех размножения равен успеху территориальных самцов. По-видимому, как раз это и наблюдается у турухтанов. Как и в случае автомимикрии, обсуждавшейся раньше, такая стратегия - это по существу форма использования существовавшей до этого стратегии.
Хотя давно уже стало ясно, что могут существовать альтернативные стратегии, лишь сейчас стало возможным дать им удовлетворительное объяснение в понятиях эволюционной теории. Ключ к этой проблеме содержится в концепции «эволюционно стабильной стратегии», развитой Мэйнардом Смитом, Паркером и другими.
2. Эволюционно-стабильные стратегии
Эволюционно-стабильная стратегия - это стратегия, которую нельзя улучшить какой-либо другой возможной альтернативной стратегией при условии, что достаточное число членов популяции приняло ее. Это иной способ утверждения того, что наилучшая стратегия для особи зависит от стратегий, принятых другими членами популяции. Поскольку это же самое относится и ко всем особям в популяции, настоящую ЭСС никакой мутантный ген успешно внедрить не может. Рассмотрим в качестве примера вопрос конфликтов и оценки соперников. Животные, которые используют стратегию избегания ненужных конфликтов, обычно получают эволюционное преимущество. Эту идею можно формализовать следующим образом: предположим, что в популяции представлены два типа стратегий: стратегия ястреба, которая состоит в том, чтобы сражаться до гибели или ранения соперника, даже если есть риск получить ранение самому, и стратегия голубя, которая заключается в угрозах и демонстрации силы наряду с избеганием серьезных конфликтов. Эти две стратегии - крайние примеры того, что может существовать в реальной жизни. Допустим, что мы приписываем результаты в виде возрастания приспособленности следствию конфликта между двумя животными. При этом победитель получает + 50 очков, а проигравший 0. Пусть цена потери времени на демонстрацию силы будет равна —10, а цена ранения —100. У нас будут четыре возможных типа столкновений в популяции, содержащей как «ястребов», так и «голубей», средние результаты их показаны в табл. 1. Можно видеть, что в столкновении ястреба с ястребом каждый в среднем останется в проигрыше, поскольку, хотя и существует 50%-ная вероятность победы в данном столкновении, есть также и 50%-ная вероятность поражения. Средний итог каждого ястреба будет равен —25 как результат нападения со стороны другого. Если ястреб встречает голубя, то ястреб всегда выигрывает 50 очков, тогда как голубь не выигрывает ничего, поскольку, избегая конфликта, он избегает ранения. Когда голубь встречает голубя, каждый из них угрожает другому и выигрывает половину состязания без борьбы. Средний итог составляет в этом случае +15.
Легко понять, что ястреб смог бы внедриться в популяцию, образованную целиком из голубей. Хотя голуби в среднем получают в конфликтах друг с другом + 15, вторгнувшийся ястреб имел бы выигрыш + 50 в каждом столкновении с голубем и таким образом получил бы преимущество. В популяции из одних ястребов средний результат составляет - 25 очков, поэтому голубь смог бы легко захватить ее, так как голубь получает в состязании с ястребом 0, что лучше чем — 25. Итак, мы видим, что ни популяция, состоящая только из голубей, ни популяция, состоящая только из ястребов, не могут устоять против вторжения и ни одна из этих стратегий не является примером ЭСС. Однако возможно, что смешанная популяция из ястребов и голубей могла бы дать устойчивую ситуацию, если бы их соотношение достигало определенной величины. Пусть доля ястребов будет h, а доля голубей -. Средний результат для голубя можно подсчитать из вероятности встречи ястреба или другого голубя и итога каждого типа конфликта:
D = 0h + 15.
Точно так же средний результат для ястреба будет
H = -25h + 50.
Если D равно H, то равновесие будет устойчивым; и тогда h = 7/12, а = = 5/12. Поэтому ЭСС будет наблюдаться
Таблица 1. Соревнование между ястребами и голубями
Результат подсчитывается следующим образом: победителю +50, раненому —100, проигравшему 0, за демонстрацию силы —10
Сетка средних результатов для нападающего показана внизу.
|
Нападающий |
Противник |
|
|
Ястреб |
Голубь |
|
|
Ястреб |
|
|
|
Голубь |
|
|
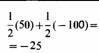


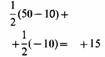
Когда «ястреб» встречается с «ястребом», мы предполагаем, что в половине случаев он побеждает, а в половине случаев получает ранение. «Ястреб» всегда побеждает «голубя». «Голубь» всегда немедленно отступает перед «ястребом». Когда «голубь» встречается с «голубем», мы допускаем, что всегда имеют место демонстрации и один из двух побеждает.
В том случае, когда 7/12 популяции составят ястребы и 5/12 - голуби. Другая возможность состоит в том, что особи ведут себя как ястребы в 7/12 состязаний и как голуби в 5/12, причем стратегия для каждого случая выбирается наугад. Это называется смешанной стратегией. В этом случае равновесие достигается только тогда, когда стратегия голубя используется в 5/12 всех конфликтов, а ястреба - в 7/12. Каждая стратегия благоприятна, если она относительно редка. Тогда ее представительство в популяции возрастает, пока снова не будет достигнута точка равновесия. И ни вторжение ястребов, ни вторжение голубей не может нарушить эту ЭСС.
Еще одна стратегия, возможная в данном случае, - это так называемая стратегия хозяина, в которой каждая особь ведет себя как ястреб, являясь хозяином территории, и как голубь, вторгаясь на чужую территорию.
Допустим, что каждая особь будет хозяином в половине конфликтов и пришельцем в другой половине; тогда результаты будут такими, как показано в табл. 2. Если популяция образована целиком из «хозяев», то средний итог конфликта будет + 25, т. е. больше, чем могли бы получить вторгшиеся голуби или ястребы, которые имели бы соответственно + 7,5 и + 12,5 в конфликте с хозяином. Таким образом, стратегия хозяина - это ЭСС. Очевидно, что эта стабильная стратегия является стратегией положения, т. е. предполагает жесткое сопротивление пришельцу и отступление перед хозяином.
Эти примеры, впервые предложенные Мэйнардом Смитом, сильно упрощают ситуацию, встречающуюся в природе. Тем не менее они представляют значительное продвижение в развитии эволюционной теории; стало ясно, что такой последовательный подход к сложной ситуации в принципе может решить давно поставленные проблемы. Основные принципы ЭСС, возможно, следует разрабатывать применительно к конкретной природной ситуации, но это справедливо для любого достаточно общего закона науки.
Фактически стратегии хозяина, голубя и ястреба можно наблюдать в природе. Например, самцы пятнистой древесницы конкурируют за брачную территорию. Конфликты у них непродолжительны, и владелец территории всегда побеждает. В этом примере стратегия хозяина используется всеми членами популяции. Стратегия голубя часто обнаруживается у животных. обладающих потенциально опасным оружием. Например, у орикса очень острые прямые рога, которыми можно нанести смертельные раны. Их можно использовать в защите против хищников, но в схватках друг с другом рога используются чисто ритуальным образом и самец никогда не бьет ими противника в бок. Точно так же у гремучих змей конфликты носят ритуальный характер для демонстрации силы, когда каждая змея пытается прижать противника к земле. Они не используют против соперников ядовитые зубы. Стратегия ястреба не универсальна в животном мире, но она может проявляться в конфликтах за ценные ресурсы, такие, как возможность спариваться. У видов, у которых самки труднодоступны или про-
Таблица 2. Конфликты между «ястребами», «голубями» и «хозяевами».
Результат подсчитывается, как и в табл. 1: победителю +50, раненому — 100, проигравшему 0, за демонстрацию —10.
Сетка средних результатов для нападающего показана внизу
|
Нападающий |
Противник |
||
|
Ястреб |
Голубь |
Хозяин |
|
|
Ястреб |
-25 |
+ 50 |
+ 12,5 |
|
Голубь |
0 |
+ 15 |
+ 7,5 |
|
Хозяин |
-12.5 |
+ 32.5 |
+ 25 |
Обратите внимание на то, что. когда «хозяин» встречает либо «ястреба», либо «голубя», мы предполагаем, что он побеждает в половине случаев, так как играет роль «ястреба», и проигрывает в половине случаев, т.е. играет роль «голубя». Его результаты, следовательно.-это среднее из двух верхних строчек сетки. Если «хозяин» встречает «хозяина», то в половине случаев он побеждает, а в половине случаев, как пришелец, проигрывает. В этом случае за демонстрации или ранение очки не вычитаются.
Продолжительность жизни очень невелика, можно ожидать конфликтов со смертельным исходом, потому что результатом этого может быть единственный за всю жизнь шанс внести свой вклад в генофонд будущих поколений. Например, самцы осагаонид вступают в смертельные схватки за возможность оплодотворить самок внутри соцветия инжира. У самцов мощные челюсти, и они могут перекусывать других ос пополам. Гамильтон обнаружил одно соцветие, в котором было 15 самок, 42 самца, погибших или погибающих от ран, и 12 самцов без каких бы то ни было повреждений. У овцебыков до 10% взрослых самцов за год могут погибать в результате схваток из-за самок. Серьезные ранения могут получать самцы благородных оленей, однако лишь после довольно продолжительной схватки.
3. Стратегия роющей осы
Самцы большой золотой роющей осы откладывают яйца в подземные норки, куда они перед этим закладывают длиннорогих кузнечиков, служащих пищей для личинок. Джейн Брокман тщательно изучила поведение самок этих ос. Она почти непрерывно проводила записи гнездовой активности 68 индивидуально покрашенных самок на трех различных участках на протяжении шести сезонов размножения. Брокман обнаружила, что самки либо сами роют норку, либо занимают уже готовую. Чтобы вырыть норку, самке требуется в среднем 100 мин. Затем она приносит в эту норку парализованных кузнечиков, откладывает одно яйцо и запечатывает норку. После этого цикл повторяется. Может случиться так, что ее норку займет другая оса, которая также обеспечивает питанием будущую личинку. Обе осы могут снабжать одну и ту же норку, не имея в это время другой вырытой норки. Поскольку обе осы тратят большую часть времени на охоту, встретиться они могут нескоро. Когда же они наконец встретятся, они вступают в схватку, и одна оса обычно изгоняется. В конечном счете только одна оса откладывает яйцо в подготовленную ячейку.
Для самки осы возможны две стратегии. Она может выкопать собственную норку, но при этом существует небольшой риск, что другая оса попытается ее занять, или она может занять уже готовую норку, не тратя сил на ее выкапывание, но подвергаясь риску быть изгнанной хозяйкой. Наилучшая стратегия будет зависеть от того, какая стратегия принята большинством других самок ос в окрестности. С одной стороны, если почти все другие самки устраивают собственные гнезда, то лучше принять стратегию занимания гнезда, так как в окрестностях будет множество норок, используемых малым количеством ос. С другой стороны, если большинство других ос пользуется стратегией вселения, то конкуренция будет весьма сильна, и в этом случае лучше выкопать собственную норку. Следовательно, ЭСС лежит между этими двумя крайностями.
Можно измерить успех этих двух стратегий числом яиц, отложенных в данный период времени. Роющие осы используют смешанную стратегию, поэтому следует сравнивать не успех отдельных особей, а результаты решений «вселяться» или «копать». На основе допущения, что эти альтернативные решения и составляют смешанную ЭСС, можно предположить, что число отложенных за единицу времени яиц будет одинаковым, независимо от решения осы - вселяться в уже готовую норку или копать собственную. Данные проведенных наблюдений позволяют сказать, что успех одинаков при всех возможных последствиях этих двух решений. Подсчет числа отложенных яиц не выявил каких-либо значительных различий между этими двумя стратегиями. Такое заключение подтверждает гипотезу, что «вселение» и «выкапывание» суть компоненты смешанной ЭСС.
Как мы уже знаем, если две самки занимают одну норку, схватка между ними обычно бывает неизбежна. Они становятся «на дыбы», наносят удары раскрытыми жвалами и борются друг с другом. Схватка продолжается от 2 до 16 мин и кончается, когда побежденная оса покидает поле боя. В 18 из 23 наблюдавшихся случаев побежденная оса не возвращалась к гнезду, тогда как в 5 случаях она возвращалась много часов спустя. Наградой победителю служит возможность использования норы, но ее ценность зависит от количества кузнечиков, лежащих там. Норка, в которой находятся четыре кузнечика, готова для откладывания в нее яиц, и за нее стоит сражаться. Норка, в которой нет кузнечиков, все же стоит некоторых усилий, так как победитель сберегает энергию, необходимую для выкапывания новой норки. Исходя из этого, можно было бы ожидать, что каждая оса сражается с такой силой, которая зависит от награды за победу. Не стоит очень настойчиво сражаться за пустую норку, поскольку усилия и риск в сумме превышают усилия и риск, которые потребовались бы для выкапывания новой норки.
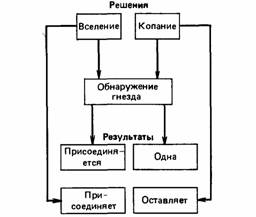
Рис. 2. Смешенная стратегия самки роющей осы.
На рисунке 2 показаны последствия решений копать новую норку или вселиться в уже готовую. Выкопанную норку иногда приходится оставить, или же в ней может обосноваться другая оса. При вселении в уже готовую норку оса может столкнуться с другой осой или же к ней может вселиться другая оса.
Проблема заключается в том, что ситуация одинакова для обеих ос. Если обе осы знают, сколько кузнечиков лежит в норке, то можно ожидать, что они будут сражаться с одинаковой силой. Будет ли такая ситуация эволюционно стабильной? Доукинс и Брокман считают, что не будет. Если норка стоящая, обе осы будут сражаться в течение долгого времени, но и уступят примерно в одно время, причем победитель определится случайным образом. За менее стоящую норку схватка будет короче, но обе ее участницы все же будут нести соответствующие затраты. Такая ситуация может иметь место в игровой стратегии: «В половине случаев эти встречи закончатся тотчас же без схватки; в оставшейся половине они будут продолжаться в течение какого-то времени, пока одна из соперниц не отступит». В популяции, где преобладают «разумные» осы, сражающиеся в соответствии с общей ценностью норки, игрок окажется в выигрыше, поскольку будет побеждать в среднем в половине схваток, но ничем не будет платить за проигранные встречи. Однако чисто игровая стратегия, по-видимому, не будет ЭСС.
Доукинс и Брокман проанализировали обстоятельства, соответствующие наблюдавшимся схваткам между двумя осами. Для каждой из 23 схваток они установили: 1) продолжительность схватки; 2) какая оса копает норку; 3) в течение какого времени каждая оса связана с гнездом; 4) какая оса крупнее; 5) сколько кузнечиков каждая оса закладывает в гнездо; 6) какая оса побеждает. Оказалось, что победитель не обязательно крупнее, или что он выкапывает норку, или что он был последним в норке. Одиннадцать ос сражались более одного раза, однако не было выявлено какого-либо достоверного индивидуального влияния на распределение победителей и проигравших. Доукинс и Брокман обнаружили, что осы, заложившие в гнездо наибольшее число кузнечиков, чаще были победительницами. Такой результат удивителен, поскольку он представляет собой типичный пример так называемого «конкорд-эффекта», т.е. поведение в будущем определяется на основе прошлых вложений, а не перспектив в будущем. Как хорошо известно, решения относительно самолета «Конкорд» были основаны на этих соображениях, однако едва ли можно ожидать, что естественный отбор действует таким же способом.
Продолжительность схватки в высокой степени коррелирует и с числом кузнечиков, запасенных побежденным. Схватка кончается, когда проигравший перестает сопротивляться, поэтому он в конечном счете и определяет продолжительность борьбы. Доукинс и Брокман убедительно показали, что время, когда проигравший прекращает схватку, определяется степенью его вклада в гнездо. Это также пример «конкорд-эффекта». Наилучшей стратегией всегда будет являться стратегия, которая основывается на ожидании будущего результата, но самка роющей осы, по-видимому, этого не знает.
Очевидно, что осы не могут оценивать содержимое норки, но способность к этому, вероятно, могла бы отбираться в эволюции. Они могут подсчитать, сколько кузнечиков уже поймано, однако неясно, каким образом они это делают. Как мы уже видели, наилучшая стратегия для особи - сражаться с усилием, пропорциональным общему числу кузнечиков в гнезде, хотя это, возможно, и не ЭСС. Отнюдь не очевидно, что «стратегия конкорд» обязательно является наиболее вероятной ЭСС. Доукинс и Брокман просто предположили, что это она и есть. Однако они все же установили важный принцип в эволюционной теории: стратегия, которая является наилучшей для особи, не обязательно будет отбираться в эволюции, поскольку она может не быть эволюционно стабильной стратегией.