Развитие сердца на примере амфибий
Министерство образования и науки Украины
Днепропетровский Национальный Университет
Биолого-экологический факультет
Кафедра геоботаники, почвоведения и экологии
Аналитический обзор
Развитие сердца на примере амфибий
План
Строение сердца амфибий
Развитие сердца на примере амфибий
1. Строение сердца амфибий
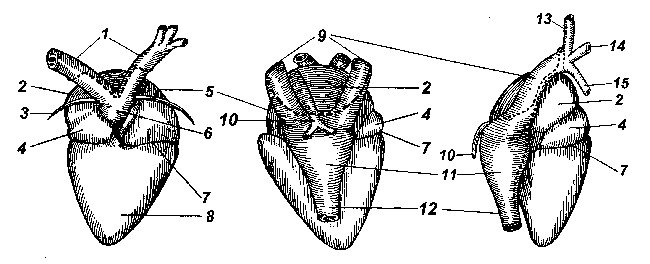
Рис. Общее строение сердца амфибий.
Сердце у амфибий расположено в грудобрюшной полости под грудиной. Его общее строение с дорсальной и вентральной сторон и сбоку (справа) показано на рисунке: 1 - дуги аорты; 2 - правое предсердие; 3 - щетинка, проведенная в отверстие под артериальным стволом; 4 - луковица аорты (артериальный конус); 5 - левое предсердие; 6 - артериальный ствол; 7 - венечная борозда; 8 - желудочек; 9 - передние (краниальные) полые вены; 10 - легочные вены (на виде сбоку только правая вена); 11 - венозный синус; 12 - задняя (каудальная) полая вена; 13 - наружная яремная вена; 14 - безымянная вена; 15 - подключичная вена).
Традиционно считается, что сердце земноводных состоит из трех камер, двух предсердий и одного желудочка. Строго говоря, это не совсем так. К сердцу относятся еще два отдела, явственно обособленные как отдельные камеры у низших позвоночных: рыб, амфибий, некоторых рептилий. Это венозный синус (венозная пазуха) и артериальный конус (луковица аорты).
Венозный синус представляет собой тонкостенную камеру, образованную слиянием полых вен - задней (каудальной) и двух передних (краниальных), левой и правой. Синус располагается с дорсальной стороны сердца, и его можно рассмотреть, если аккуратно оттянуть верхушку сердца вперед, к голове.
Мускулистый артериальный конус расположен вентрально между желудочком и коротким артериальным стволом (он является частью сосудистой системы), от которого отходят левая и правая дуги аорты. Артериальный ствол не прирастает к вентральной поверхности предсердий, под него можно провести тонкий волосок (щетинка "3" на Рис.1).
Мышечные стенки венозного синуса и артериального конуса сокращаются, и в какой-то степени участвуют в перемещении крови.
Однако внешне артериальный конус не отличается от крупного сосуда. Между полыми венами и венозным синусом также нет четкой границы, нельзя определенно указать, где заканчиваются полые вены и начинается венозный синус.
Почему венозный синус и артериальный конус считают частью сердца, а не частью сосудистой системы? Это сделано на основании особенностей эмбрионального развития сердца и сосудов. В ходе эмбриогенеза сердце, включающее венозный синус, предсердие(-я), желудочек(-чки), артериальный кону, возникает из одного зачатка, а сосуды - из другого. У теплокровных животных на определенной стадии развития венозный синус и артериальный конус также отчетливо выражены. Затем венозный синус трансформируется в синусный узел (пейсмекерную зону, пейсмекер), расположенный в стенке правого предсердия, а артериальный конус преобразуется в безмышечное кольцо, расположенное на границе между левым желудочком и аортой. Таким образом, синусный узел теплокровных животных является гомологом венозного синуса низших позвоночных.
Пейсмекер (англ. "pacemaker", водитель ритма) определяет ритм сердечных сокращений. Здесь имеются специализированные мышечные волокна, обладающие автоматией. В пейсмекерных кардиомиоцитах спонтанно, с определенной периодичностью, возникают волны возбуждения, которые затем последовательно распространяются на миокард предсердий, желудочка и артериального конуса. На границах различных отделов сердца (у лягушки между венозным синусом и предсердиями, предсердиями и желудочком, желудочком и артериальным конусом) волна возбуждения проводится с меньшей скоростью, возникает задержка проведения возбуждения, сопровождаемая такой же задержкой волны сокращения.
Венозный синус сердца лягушки сообщается с правым предсердием через широкое овальное отверстие, окруженное мускулистым синоатриальным кольцом. Сокращения синоатриального кольца частично препятствуют обратному току крови из правого предсердия в венозный синус. Других клапанных структур здесь нет.
Легочные вены, несущие аэрированную кровь, перед входом в левое предсердие объединяются в общую легочную вену. Настоящих клапанов здесь также нет.
Передние полые вены образуются слиянием наружных яремных, подключичных и безымянных вен. Короткая задняя полая вена выходит из печени.
Предсердия отделяются от желудочка венечной бороздой. Она разделяет сердце на переднюю часть (предсердия, входящие и выходящие сосуды), и заднюю часть (желудочек).
Снаружи сердце окружено перикардом, который можно представить себе как тонкостенный мешочек, надвинутый на сердце со стороны его верхушки. Внутренний листок перикарда (или эпикард) является наружным слоем сердца. Между эпикардом и наружным листком перикарда, в перикардиальной полости, находится перикардиальная жидкость. Граница прикрепления наружного листка перикарда к стенкам сердца и сосудов показана на Рис.1 пунктирной линией.
В сердце лягушки коронарные сосуды имеются только в стенках артериального конуса. Ткани венозного синуса, предсердий и желудочка снабжаются кислородом за счет перекачиваемой крови.
Миокард предсердий не переходит непосредственно в миокард желудочка. Контакт между ними осуществляется через относительно компактный пучок специализированной мышечной ткани, расположенный в области атриовентрикулярного отверстия, являющегося общим входом для правого и левого предсердия. Здесь имеются хорошо выраженные атриовентрикулярные створчатые клапаны.
Как и у всех более высокоорганизованных классов, в правое предсердие впадают вены большого круга, несущие венозную кровь, в левое— малого с артериальной кровью.
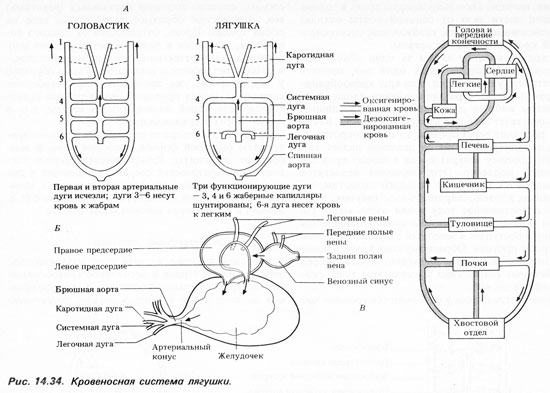
Рис. Схема строения сердца амфибий.
При сокращении предсердий в желудочек, внутренняя стенка которого снабжена большим количеством мышечных перекладин, одновременно попадают обе порции крови. Полного их смешения из-за своеобразного строения стенки желудочка не происходит, поэтому при его сокращении первая порция венозной крови поступает в артериальный конус и с помощью спирального клапана, находящегося там, направляется в кожно-легочные артерии. Кровь из середины желудочка, смешанная, поступает таким же образом в дуги аорты, а оставшееся небольшое количество артериальной крови, последней попадающей в артериальный конус, направляется в сонные артерии.
Кровь, насыщенная в легких кислородом, поступает по легочной вене в левое предсердие, а дезоксигенированная кровь собирается со всего тела в переднюю и заднюю полые вены, по которым она попадает в венозный синус и затем в правое предсердие. В предсердиях венозная и артериальная кровь никогда не смешивается, но оба предсердия одновременно выталкивают кровь в единственный желудочек. Смешиванию крови в желудочке частично препятствуют мышечные гребни, образуемые внутренней поверхностью дорсальной и вентральной стенок желудочка; а когда кровь поступает из желудочка в артериальный конус, то здесь ее смешивание затрудняет спиральный клапан, который не полностью подразделяет артериальный конус на два протока. Дорсальный проток, называемый кожно-легочным стволом, переходит в легочные артерии, от которых ответвляются кожные артерии. А из вентрального протока кровь поступает в общую сонную артерию и в системную дугу аорты.
2. Развитие сердца
Вся кровеносная и вся лимфатическая системы — производные мезодермы.
В ходе гаструляции происходит распространение мезодермального пласта клеток между экто- и энтодермой. На стадии нейрулы разрастание спинного и спинно-бокового участков мезодермальной мантии доходит до головной области зародыша. Разрастание это, однако, неравномерное: на брюшной стороне зародыша, в оральной и глоточной областях, остается в виде треугольника участок, в который мезодерма не проникает

В период, следующий за нейруляцией, происходит постепенное схождение (в середине свободной от мезодермы области) краев разрастающейся мезодермальной мантии (спланхнотомов). Слева и справа, к середине свободного от мезодермы участка, распространяются мезенхимного типа рыхло расположенные клетки, которые являются парным зачатком сердца. Точнее сказать, этот клеточный материал является зачатком эндокардия, т.е. эндотелия полости сердца. Клетки мезенхимы в виде продольной полоски накапливаются, и в результате формативных перемещений клеточного материала формируется тонкостенная трубка, разветвляющаяся на обоих концах: две ветви на переднем конце — это будущие брюшные аорты, а две ветви на заднем конце— это две желточные вены. Как и эндокардиальная трубка, ветви ее — будущие сосуды — состоят также только из одного слоя клеток эндотелия, возникшего вследствие дифференциации мезенхимы. Одновременно с этими процессами края мезодермального пласта клеток доходят друг до друга и срастаются под зачатком сердца (т.е, под эндокардиальной трубкой), но затем зачаток сердца покрывается висцеральным листком мезодермы.
При слиянии мезодермальных листков под и над зачатком сердца образуются перегородки — спинной и брюшной мезокардии (брыжейки сердца, рис. 92, 6). Брюшная перегородка вскоре исчезает, и благодаря этому целомические полости (правой и левой сторон) оказываются не разделенными. Впоследствии рассасывается и спинной мезокардии. Расширяющийся участок целомической полости в районе сердца образует околосердечную полость (рис. 92, 9), которая являясь сначала частью общей полости тела, впоследствии развивается в полость, совершенно обособленную от целома. Эпикардий и миокардий (сердечная мышца) возникают из клеток висцерального листка мезодермы, дифференцирующихся в наружный эпителий и мускульную ткань сердца.

Рис. Развитие сердца зародышей амфибий (по Моллье, из Гертвига, 1906): а, г, д — тритон, б — саламандра, в — лягушка; 1 — энтодерма, 2 — париентальный листок латеральной мезодермы, 3 — висцеральный листок латеральной мезодермы, 4 — зачаток эндокардия, 5 — эндокардиальная трубка, 6 — брюшной мезокардии, 7 — перикардий, 8 — миокардий, 9 — околосердечная полость, 10 — полость сердца
Перикардий является производным париетального листка спланхнотома.
Сначала сердце — это полая, почти прямая, без всяких подразделений трубка. Путем не описываемых здесь изгибаний и поворотов, сжиманий в одних местах и расширений в других происходит изменение формы развивающегося сердца, и оно подразделяется на четыре основные части: венозный синус (sinus venosus), предсердие (atrium), желудочек (ventriculus) и артериальный конус (conus arteriosus).Задолго до полного сформирования сердца, еще прежде чем образуются периферические сосуды, уже начинается ритмическая пульсация сердца. Оно находится, образно говоря, в состоянии готовности играть свою функциональную роль.Развитие сердца амфибий явилось объектом многих экспериментальных исследований; данные этих исследований могут служить иллюстрацией влияния одних частей зародыша на другие.
В связи с не изученными до конца взаимовлияниями клеток мезодермы и прилегающей эктодермы задолго до сформирования сердца со структурой, свойственной его дефинитивному состоянию, уже имеет место специфическая клеточная и тканевая дифференциация презумптивной мезодермы сердца. Такое заключение можно сделать на основании вырезывания презумптивной сердечной мезодермы и культивирования ее вне зародыша. По Гертлеру (1928), такая складывающаяся тканевая система способна развиваться в свойственном ей направлении: дифференцируется характерная для сердца мышечная ткань, начинающая ритмично сокращаться.
При развитии сердца несомненно происходит взаимодействие всех трех зародышевых листков. Мангольду в 1936г. и Балинскому в 1939г. удалось осуществить операции полного удаления энтодермы у зародышей амфибий. Техника операций была такова, что мезодерма, производной которой является сердце, оставалась неповрежденной. Однако у таких безэнтодермных зародышей сердце совершенно не развивалось.Штор в 1924 г. обнаружил, что, достигнув определенной стадии развития, зачаток сердца может некоторое время в условиях эксплантации продолжать развиваться,— значит, независимо от окружающих его частей зародыша развиваются "пульсирующие" мышечные ткани, формируется сердечная трубка.
Подобные явления наблюдаются и в экспериментах на зачатках других органов.
Согласно опытам Экмана (1925), если вырезать не весь парный зачаток сердца, а только правую или левую половины его и культивировать их вне зародыша, каждая половина (как оставшаяся в зародыше, так и удаленная) может дифференцироваться в ритмично сокращающиеся ткани. Нельзя считать удивительным явлением, что развитие сердца в условиях культивирования вне организма половины его зачатка вскоре останавливаются, так как в норме для развития сердца требуется его функционирование, чтобы была циркуляция крови. Можно добиться развития двух сердец у одного эмбриона, если сделать надрез перед тем, как соединятся презумптивные зачатки сердца с правой и с левой сторон. Можно поместить между двумя презумптивными зачатками сердца чужеродную ткань, в результате не произойдет срастания зачатков правой и левой сторон. В этих условиях у одного зародыша могут развиваться два пульсирующих сердца. Считают, что на стадии до слияния двух зачатков сердца они еще не "строго детерминированы".
Используемая литература:
Газарян К.Г., Белоусов Л.В. Биология индивидуального развития животных М.: "Высшая школа", 1983
Грин.Н. Биология 1996 Том 2 с.188
Токин Б.П. Общая эмбриология. Учебник для студентов биологических специальностей университетов. М.: "Высшая школа", 1970
Ноздрачев А.Д., Поляков Е.Л. Анатомия лягушки. (Лабораторные животные). М., Высшая школа", 1994. С. 320.
Ярыгин В.Н., В.И. Васильева, И.Н. Волков, В.В. Синельщикова Биология в двух томах. М.: "Высшая школа", 2003 , Т.2.