Основные свойства цитоплазмы
ОСНОВНЫЕ СВОЙСТВА ЦИТОПЛАЗМЫ
Физико-химические свойства цитоплазмы. Цитоплазма, представляющая собой основную массу протопласта (за вычетом ядра, митохондрии и пластид), имеет сложное строение, детали которого до сих пор еще не выяснены. Она состоит из большого количества высокомолекулярных веществ — биополимеров. Часть последних формирует особые структурные образования — мембраны, придающие цитоплазме значительную структурность. Цитоплазма проявляет свойства сравнительно вязкой жидкости, но одновременно и некоторые свойства твердого тела (эластичность). Подобное сочетание свойств возможно благодаря тому, что молекулы биополимеров способны образовывать временные ассоциации различных размеров. Разрушение этих ассоциаций способствует проявлению жидкостных свойств, их восстановлению — проявлению свойств твердого тела.
Структурную основу всей цитоплазмы, так называемый цитоскелет, составляют особые белки, способные преобразовывать химическую энергию в механическую работу. Это — сократительные белки, подобные тем белкам, из которых построены мышцы животных. Благодаря пустой сети мембран внутри цитоплазмы образуется множество так называемых отделов. Тем самым создаются условия для пространственного разграничения различных биохимических процессов. Эти процессы могут протекать одновременно в разных частях цитоплазмы одной и той же клетки, не мешая друг другу.
Схема строения геля
Молекулы биополимеров — белков и липидов — в мембранах расположены в строго определенной последовательности.
Благодаря такому строению мембрана обладает способностью избирательно пропускать молекулы одних веществ и не пропускать молекулы других веществ, т. е. она обладает избирательной проницаемостью, и играет важную роль в жизнедеятельности.
Цитоплазма имеет две пограничные мембраны. Одна из этих мембран, называемая плазма леммой, отграничивает цитоплазму от вакуоли. Тонопласт — наиболее устойчивая часть цитоплазмы; при гибели содержимого клетки он отмирает последним.
Значительную роль в процессах структурообразования в цитоплазме играют ионы, а также молекулы низкомолекулярных соединений, обладающие полярными группами.

Схематическое изображение различных форм плазмолиза: 1 — выпуклая; 2 — вогнутая; 3, 4 — судорож
Поступление веществ в цитоплазму. Вещество, поступившее в цитоплазму, либо связывается самой цитоплазмой, либо поступает из нее в клеточный сок. Растворы солей или Сахаров высокой концентрации обычно не проникают в цитоплазму, а оттягивают из нее воду. При этом цитоплазма отходит от стенок клетки. Это явление получило название плазмолиза. При отхождении цитоплазмы от стенок клетки она образует вогнутую поверхность (вогнутый плазмолиз), которая затем через 15—30 мин переходит в выпуклую форму (выпуклый плазмолиз).
Своеобразную форму плазмолиза можно вызвать действием раствора роданистого калия (KCNS) на клетку чешуи лука. Анионы CNS проникают в клетку и вызывают набухание мезоплазмы, внутрь вакуоли они не проникают. Мезоплазма при этом становится хорошо заметной в виде колпачков на полюсах плазмолизированного протопласта. Отсюда данный тип плазмолиза получил название колпачкового плазмолиза.
Колпачковый плазмолиз обнаруживает первый тип проницаемости цитоплазмы, когда проникшее вещество связывается самой цитоплазмой и не поступает в вакуоль.
Другой тип проницаемости цитоплазмы связан с проникновением вещества в вакуоль. Далеко не все растворы вызывают долго длящийся плазмолиз. Если плазмолизировать протопласт клетки раствором мочевины или глицерина, то сначала наблюдается плазмолиз. Затем плазмолиз сравнительно быстро заканчивается. Мочевина и глицерин быстро проникают внутрь вакуоли, увеличивают концентрацию клеточного сока, который начинает поглощать воду из окружающего раствора. При этом цитоплазма вновь подходит к стенкам клетки. Это явление получило название деплазмолиза, и такой плазмолиз называется временным.

Колпачковый плазмолиз в растворе роданистого калия
Проникновение вещества в вакуоли клетки можно наблюдать на примере действия красителя метиленового синего (водный раствор 1:5000) на какое-нибудь водное растение, например элодею. Краситель скопляется внутри вакуолей, и очень часто в них даже образуется осадок из красителя, связанного с дубильными веществами вакуолей.
Проницаемость цитоплазмы связана с активной жизнедеятельностью (дыханием) растения. Поступление веществ (солей или, вернее, их катионов и анионов) в цитоплазму идет за счет обмена их на ионы, выходящие из клетки. Это происходит таким образом, что образованные в процессе дыхания Н+ и НСОз (ионы угольной кислоты) выделяются в окружающий раствор, а на их место в цитоплазму поступают ионы калия и натрия вместо ионов водорода, а на место иона НСОз— соответственные анионы.
Не всегда вещества проникают в цитоплазму за счет ее активной жизнедеятельности. Дело в том, что поверхностные слои цитоплазмы богаты липидами, которые не образуют сплошной пленки на поверхности цитоплазмы, а чередуются с молекулами белков или вкраплены в белковый остов поверхностных слоев, т. е. на поверхности цитоплазмы образуется своеобразная мозаика из участков белков и липидов. Растворимые в липидах вещества (спирт, эфир, хлороформ и др.) очень легко проникают в цитоплазму.
Вода и соли, нерастворимые в липидах, проникают в цитоплазму через белковые слои.
Проницаемость цитоплазмы не остается постоянной в течение жизни растения, а меняется с возрастом, а также увеличивается при повышении температуры и интенсивности освещения.
Движение цитоплазмы. Одним из характерных свойств цитоплазмы является ее способность к движению. Движение цитоплазмы и находящихся в ней включений происходит как в постенном слое, так и в тяжах, связывающих ядро с цитоплазмой.
Скорость перемещения цитоплазмы в эпидермисе чешуи лука составляет примерно 5—7 м/с. На скорость движения Цитоплазмы влияют температура, свет и другие факторы. В одном из опытов движение цитоплазмы в клетках водного растения валлиснерии начиналось при температуре 1,25°С, шло с наибольшей интенсивностью при 38,5°С и останавливалось при 45°С.
Движение цитоплазмы играет большую роль в жизнедеятельности растительного организма, способствуя перемещению веществ из одной клетки в другую.
Вязкость — одно из важнейших свойств цитоплазмы. Она очень сильно колеблется в зависимости от вида растения, а также от фаз его развития. У некоторых растений вязкость цитоплазмы немного превышает вязкость воды, а у других достигает вязкости глицерина, превосходящего в этом отношении воду в 87 раз. Вязкость цитоплазмы тесно связана с обменом веществ: чем выше вязкость, тем обычно менее интенсивен обмен. У созревших семян цитоплазма переходит в студенистое состояние — гель. Высокая вязкость цитоплазмы способствует увеличению устойчивости растений к повышенной температуре.
Сравнительное определение вязкости цитоплазмы производят по времени перехода вогнутого плазмолиза в выпуклый.
Насколько тесно связана вязкость цитоплазмы с температурой коагуляции белков, видно на примере озимой ржи. Вязкость цитоплазмы в различных органах ржи неодинакова. В тех органах, где она выше, белки цитоплазмы свертываются при более высокой температуре.

ОРГАНОИДЫ КЛЕТКИ
Помимо цитоплазмы, в световом микроскопе можно наблюдать и другие составные части, получившие название органоидов клетки. К ним относятся ядро, пластиды, митохондрии.
Крупные органоиды (ядро, пластиды) хорошо видны в световом микроскопе, другие органоиды (митохондрии, рыбосомы) и структурные элементы цитоплазмы (аппарат Гольджи, эндоплазматическая сеть) только лишь в электронном микроскопе.
Ядро является обязательной составной частью любой растительной и животной клетки. Оно имеет обычно округлую или слегка вытянутую форму. Абсолютные размеры ядра не превышают 7—8 мкм. Ядро состоит из ядерной плазмы (кариоплазмы), ядрышка, ядерной оболочки, отграничивающей ядро от окружающей цитоплазмы. Кариоплазма содержит твердую часть — хроматин и жидкую — ядерный сок. Хроматин — это сложное образование, в состав которого входят нуклеопротеиды, т. е. соединения белков с нуклеиновыми кислотами. В ядре содержится дезоксирибонуклеиновая кислота, ДНК, а в ядрышке — рибонуклеиновая кислота — РНК.
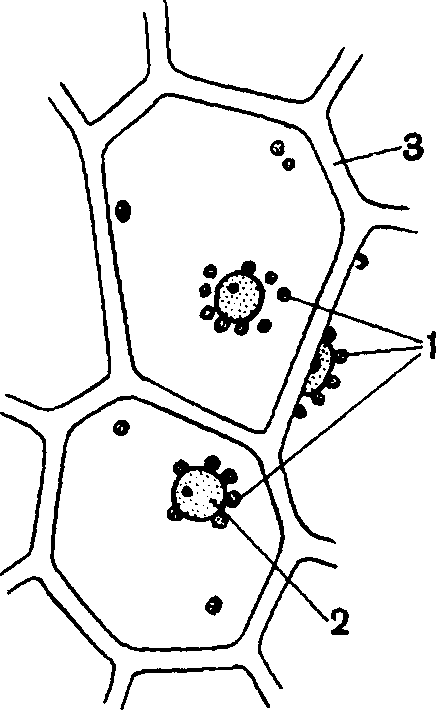
Лейкопласты в эпидермисе листьев традесканции: 1— лейкопласты; 2—ядро; 3— оболочка
Ядро играет огромную роль в жизни клеток. При делении клеток (митозе) из хроматина ядра образуются хромосомы, которые являются носителями наследственности. Число хромосом строго определенно для каждого отдельного вида растений и животных. Ядро имеет большое значение и в неделящейся клетке. О роли ядра можно судить по изучению физиологии безъядерных клеток. В 1890 г. И.И. Герасимов, действуя на делящуюся клетку водоросли спирогиры низкой температурой, или эфиром, получал безъядерные клетки и клетки, содержащие двойное количество ядерного вещества. Безъядерные клетки хотя и продолжали некоторое время жить, но переставали расти, обмен веществ в них шел ненормально. Образовавшийся в процессе фотосинтеза крахмал не претерпевал дальнейших превращений, и клетки им переполнялись.
Отделенная от ядра цитоплазма сравнительно быстро погибает из-за нарушения обмена веществ. Изолированное от цитоплазмы ядро также не может существовать. Жизнеспособными являются только клетки, содержащие цитоплазму и ядро. Пластиды. Пластидами называются особые органоиды в клетке. К ним относят бесцветные лейкопласты, зеленые хлоропласты и оранжевые хромопласты. Все виды пластид могут возникать из бесцветных пропластид. Окраска пластид обусловлена особыми пигментами (красящими веществами): в хлоропластах — зеленым хлорофилле м, а в хромопластах — оранжевым каротином.
Лейкопласты имеются в клубнях и корневищах растений, где они образуют запасной крахмал. Кроме того, они встречаются в эпидермисе листьев некоторых растений, например в листьях традесканции. Роль их в эпидермисе связана с тем, что они содержат ряд ферментов и способствуют ферментативной деятельности клеток.
Известно, что выращенные в темноте растения бывают бледно- желтого цвета.
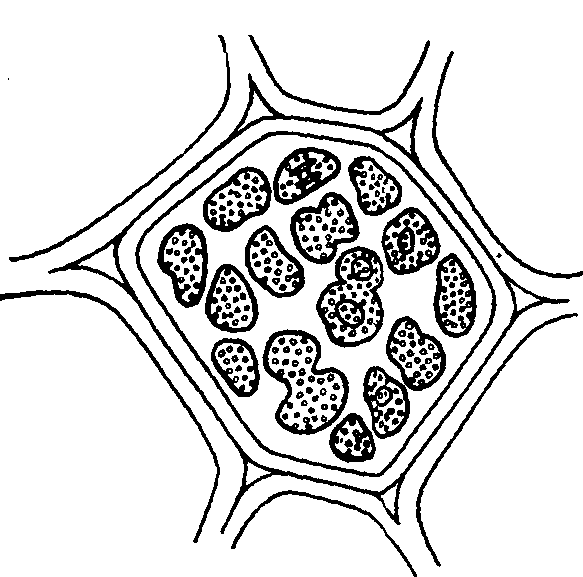
Хлоропласты в листьях лехалениума
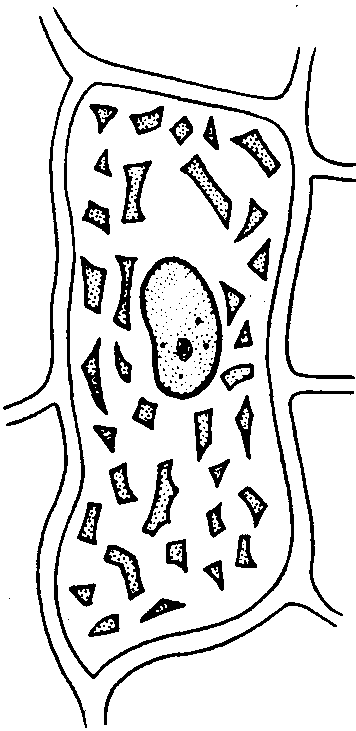
Хромопласты лепестков настурции
Хлоропласты встречаются в лепестках, плодах и некоторых корнях (морковь). Могут возникать из пропластид и из хлоропластов. Плоды многих растений бывают сначала зелеными — содержат хлоропласты (томаты, рябина, шиповник), затем они краснеют, так как у них разрушается — хлорофилл и остается оранжевый пигмент каротин. В хлоропластах также имеется каротин, но он маскируется зеленым пигментом хлорофиллом. Хромопласты часто имеют игольчатую или неправильную форму, так как каротиноиды в них кристаллизуются.
Помимо пластид, в клетках имеются и другие органоиды — митохондрии, размером около 1 мкм, играющие большую роль в дыхании растений.
СУБМИКРОСКОПИЧЕСКОЕ СТРОЕНИЕ КЛЕТКИ
Электронный микроскоп и основные принципы его работы.
Создание и применение светового микроскопа способствовало развитию ряда биологических наук — цитологии, гистологии, микробиологии, успехи которых привели к крупным практическим результатам в области медицины, сельского хозяйства и ряда отраслей промышленности.
Однако уже к концу XIX в. выяснилось, что возможности светового микроскопа ограничены. Частицы мельче 0,2 мкм в световом микроскопе уже неразличимы. Они лежат ниже разрешающей способнорти микроскопа, т.е. способности различать две отдельные точки.
Развитие физики электронных явлений дало возможность использовать для микроскопа электронные излучения. На смену стеклянным линзам пришли "электронные линзы" — электромагнитные поля, способные фокусировать и преломлять электронный пучок.
В сороковых годах XX в. начали изготовляться первые электронные микроскопы. Разрешающая способность лучших из них составляет в настоящее время около 10-8— 2-10-8 см, причем увеличение достигает 1 000 000 раз.
Строение клетки под электронным микроскопом. Изучение ультратонких срезов клеток привело к открытию, что все структурные элементы цитоплазмы клетки и клеточные органоиды имеют мембранное строение. Электронный микроскоп помог увидеть тонкое строение клеток растений и животных. Цитоплазма состоит из матрикса (основы) и заключенных в нее органелл (хлоропласты, митохондрии, аппарат Гольджи и др.), а также мембран. На поверхности цитоплазмы имеются мембраны — плазмалемма а на поверхности вакуоли — тонопласт. Мембраны клетки состоят из липидов и белков. Таким образом, строение мембраны имеет мозаичный характер.
Эндоплазматическая сеть состоит из длинных канальцев, пронизывающих цитоплазму; представляет собой выросты двойной ядерной мембраны. Эндоплазматическая сеть образует в цитоплазме непрерывную систему, ограничивающую полости, по которым перемещаются вещества между ядром и цитоплазмой. Эндоплазматическая сеть увеличивает поверхность цитоплазмы и, как бы изолируя ее на отдельные участки, способствует протеканию разнообразных процессов в [различных частях клетки.
Аппарат Гольджи — структурный элемент цитоплазмы клетки, названный в честь итальянского ученого, который впервые описал это образование. Долгое время считали, что аппарат Гольджи имеется только в животных клетках. В дальнейшем он был обнаружен и в растительных клетках.
Аппарат Гольджи состоит из системы мембран, сгруппированных в стопки. По концам мембран наблюдаются вздутия, которые отшнуровываются от них в виде пузырьков, способных превращаться в вакуоли или цистерны.
Ядро. Изучение ядра в электронном микроскопе показало, 100% то оно окружено оболочкой, состоящей из двух мембран — внутренней и наружной. Предполагают, что наружная мембрана является продолжением мембран, составляющих эндоплазматическую сеть цитоплазмы. В ядерной оболочке имеются поры (до двухсот), через которые происходит обмен между веществами ядра и цитоплазмы. Ядро состоит из округлых гранул, образованных ДНК и РНК в соединении с белками. Кроме того, в ядре имеется одно или несколько ядрышек, содержащих в основном рибонуклеиновую кислоту (РНК).
Хлоропласты. Хлоропласт отделен от цитоплазмы оболочкой. Внутри хлоропласта расположены пачки мембран, образующих пары, соединенные концами. В результате этого образуется замкнутый диск. Пачки дисков, расположенных в определенном порядке образуют граны ^Хлоропластов. У большинства растений мембрана граны переводит в более тонкую мембрану стромы. Строма является основным веществом хлоропласта и заполняет объем, не заснятый мембранной системой.
В строме находятся капельки жира, крахмальные зерна, гранулярное вещество, содержащее ферменты. Считается, что мембрана хлоропласта (толщина около 70-10-8 см) состоит из двух слоев липидов, находящихся между двумя тонкими слоями белка. Хлорофилл образует мономолекулярный слой по всей поверхности диска.
Митохондрии. Электронномикроскопическое изучение митохондрий в клетках показало, что они одеты поверхностной мембраной, под которой находится множество внутренних мембран, расположенных параллельно друг другу. Все мембраны, как мы уже знаем, состоят из нескольких слоев. Толщина слоев и расстояние между ними довольно постоянны. Между мембранами находится внутреннее пространство митохондрий, величина его различна у митохондрий разных клеток.
Митохондрии подвижны и могут перемещаться в клетке за счет собственного движения. Однако по большей части они передвигаются током движущейся цитоплазмы. На свету митохондрии движутся к хлоропластам, а в темноте — к стенкам клетки.
Митохондрии состоят на 30—40% (сухое вещество) из белков, на 25—38% из липидов. В них содержится от 1 до 6% рибонуклеиновой кислоты.
Большую роль в образовании митохондрии играют ионы кальция. При недостатке солей кальция в растении число митохондрий уменьшается.
Мембраны митохондрий содержат постоянный набор ферментов, которые участвуют в процессах окисления и накопления энергии при дыхании. Митохондрии могут осуществлять синтез близких к белкам веществ — пептидов, по-видимому, принимают участие в жировом обмене, а также в поглощении солей и воды. Под влиянием высокой температуры митохондрии набухают и теряют свою структуру.
Рибосомы в отличие от пластид и митохондрий представляют собой не микроскопические, а субмикроскопические органоиды размером от 200-10-8 —280-10-8 см.
Рибосомы состоят из белков (55%), фосфолипидов (4%) и рибонуклеиновой кислоты (40% от сухого вещества). Они содержат 65% всей рибонуклеиновой кислоты клеток.
Рибосомы образуются в ядре, вернее, в ядрышке,' где происходит их сборка из отдельных более мелких частиц. Рибосомы содержатся в цитоплазме и хлоропластах. Особенное хорошо они заметны в клетках, интенсивно синтезирующих белок, так как основная их функция — синтез белка.
Лизосомы. Круглые пузырьки, осуществляющие внутреннее переваривание веществ. Содержат ряд ферментов, отделены мембраной от цитоплазмы.
Пероксисомы. Мелкие пузырьки. В них происходит фотодыхание.
Микротрубочки. Располагаются в наружном слое цитоплазмы. Состоят из белка тубулина. Входят в состав веретена при делении клеток (митозе).
НАСЛЕДСТВЕННАЯ ИНФОРМАЦИЯ И РЕГУЛЯЦИЯ У РАСТЕНИЙ
Основное свойство образовывать один и тот же вид при развитии растения из воспроизводящей клетки, т. е. свойство наследственности, заключено в ядре и связано с ДНК.
Большое значение в регуляции функций организма играет цитоплазматическая связь между клетками. Они соединены цитоплазматическими нитями — плазмодесмами, объединяющими клетки как бы в единое целое. Впервые плазмодесмы были обнаружены проф. Горожанкиным.
Очень важную роль играет гормональная регуляция физиологических функций. В растениях образуются следующие гормоны: ауксины, гиббереллины, цитокинины, абсцизовая кислота и этилен. О значении каждого из этих гормонов будет идти речь ниже.
Световая регуляция осуществляется длиной дня, продолжительность которого влияет на развитие растений. Растения делятся на растения короткого дня, зацветающие лишь при укорочении длины дня осенью, растения длинного дня, зацветающие только при длине дня больше 12 ч, и нейтральные растения, зацветающие как на коротком, так и на длинном дне. Наконец, регулирование светом может осуществляться с помощью специального вещества — фитохрома. Фитохром имеет две формы: активную и неактивную. Активная форма тормозит рост, неактивная форма дает симптомы этиоляции. По своей природе фитохром близок к пигментам сине-зеленых и красных водорослей фикоциану и фикоэритрину. Освещение красным светом с длиной волны 660 нм переводит фитохром в активную форму, а более длинноволновый красный свет приводит фитохром уже в неактивное состояние.
Таковы в очень коротких чертах основы регуляции отдельных функций растений.
Как мы уже знаем, белковые вещества, составляющие основу живых организмов, построены из аминокислот. В организмах имеется свыше 20 аминокислот. Эти 20 аминокислот могут складываться различным образом и образовывать разнообразные белки, которые выполняют ту или иную функцию в организме. Число комбинаций сложения аминокислот почти безгранично, и это определяет специфичность отдельных белков. План построения белка из аминокислот заложен, или, как говорят, "закодирован", в молекуле ДНК, находящейся в ядре.
Нуклеиновые кислоты ДНК и РНК состоят из трех структурных элементов: азотистых оснований, сахара и фосфорной кислоты. Соединяясь, эти вещества образуют нуклеотиды. Нуклеиновые кислоты представляют собой продукты полимеризации (уплотнения) большого числа нуклеотидов.
Полимерами называют вещества, молекулы которых состоят из одинаковых, периодически повторяющихся групп атомов. Например, полиэтилен имеет строение: (—СНг—СНг)>л>—СНг—СНг— ДНК — сложный полимер с молекулярной массой от 4 до 10 млн. Она состоит из двух полинуклеотидных цепей, образованных большим числом соединенных между собой нуклеотидов. В состав ДНК входит сахар дезоксирибоза (С5Н10О4). ДНК очень чувствительна к действию кислот и при гидролизе в молекулярном растворе соляной кислоты при 60°С через несколько минут распадается на нуклеотиды.
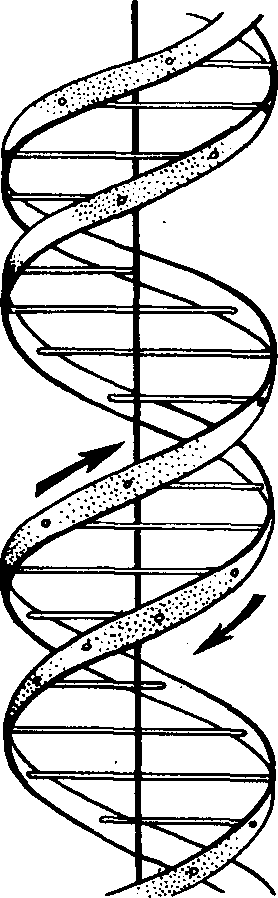
Схема молекулы ДНК
РНК в отличие от ДНК состоит из одной цепи полинуклеотидов и вместо дезоксирибозы содержит d-рибозу (С5Н10О5). РНК распадается на нуклеотиды под влиянием щелочей. В ядре при непосредственном участии ДНК образуется РНК, которая содержит полученные oт ДНК сведения о порядке сложения аминокислот в различные белки. Эта РНК носит название информационной или посредника. На каждую нить информационной РНК садится по нескольку рибосом. Эта цепочка рибосом называется полисомой. В полисомах происходит синтез белка при участии содержащейся в рибосомах рибосомальной РНК. Отдельные рибосомы движутся по нити РНК, считывают заложенную в ней информацию (сведения), полученную в ядре от ДНК и укладывают аминокислоты в полипептидные цепи. Аминокислоты, образовавшиеся в процессе обмена веществ, под водятся к полисомам особой, тоже образовавшейся первоначально в ядре РНК-переносчиком или транспортной РНК названной так потому, что она переносит активированны соответственными ферментами аминокислоты на рибосомы. Таких различных РНК-переносчиков имеется примерно 20 по числу аминокислот, из которых строятся белки.
Таким образом, в синтезе белка в растениях участвуют различные РНК: рибосомальная РНК, информационная РНК передающая порядок укладки аминокислот в полипептидны цепи, и транспортная РНК, которая доставляет активированные соответственными ферментами аминокислоты к полисе мам. Такова схема синтеза белков в растении.
Каждая клетка организма содержит полный набор информации о строении всех белков, которые она может синтезировать. Поэтому из маленького кусочка листа многих растений, например бегонии, может развиться целое растение ил даже из одной клетки (в культуре тканей, см. ниже) может развиться целый организм. С другой стороны, только из ядра или только из цитоплазмы новый организм не образуется, так как весь процесс образования белков происходит только в целостной клетке, состоящей из ядра и цитоплазмы.
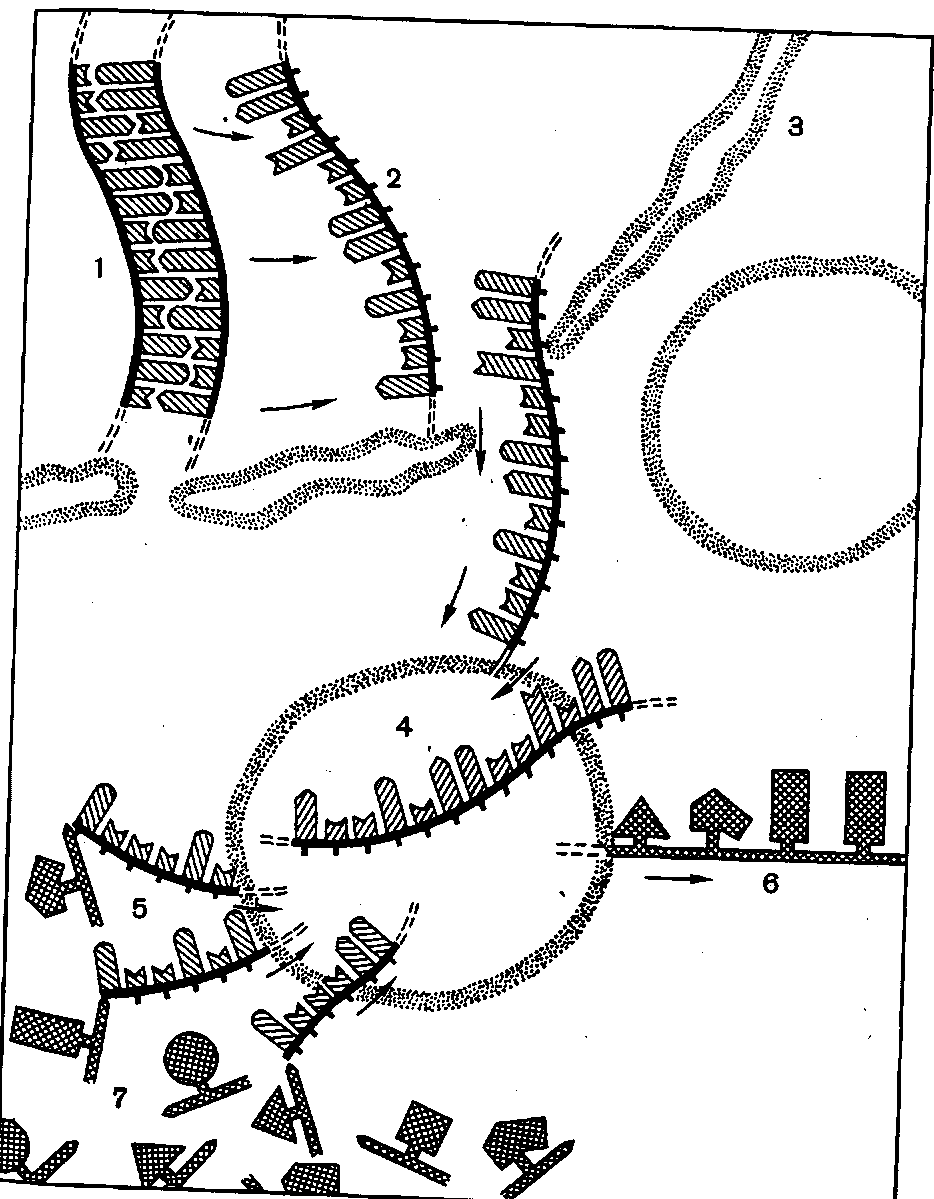
Схема биосинтеза белка в клетке