Синдром гибридного дисгенеза у Drosophila melanogaster
СИНДРОМ ГИБРИДНОГО ДИСГЕНЕЗА У DROSOPHILA MELANOGASTER
Введение
Мобильные генетические элементы (МГЭ) представляют дискретные сегменты ДНК, которые могут перемещаться из одного местоположения в другое внутри хромосом или между ними. На данный момент мобильные генетические элементы обнаружены в геномах практически всех изученных организмов (Хесин, 1984). Геном Drosophila melanogaster содержит около 50-ти различных семейств мобильных генетических элементов, которые вместе составляют 10-15 % ДНК этого вида (Finnegan, Fawsett, 1986; FlyBase, 1999). Число копий элементов отдельных семейств варьирует от нескольких до сотни, и при активации они могут оказывать значительное влияние на функционирование генома (Britten, 1997) и на генетическую изменчивость (Kidwell, Lisch, 1997). Мобильные генетические элементы имеют несколько механизмов перемещения и могут выполнять разные функции (табл. 3), в связи с чем, активация различных семейств мобильных элементов может иметь как отрицательные, так и положительные последствия для генома хозяина (Kidwell, Lisch, 1997).
Синдром гибридного дисгенеза
Некоторые МГЭ дрозофилы способны активироваться в особых межлинейных скрещиваниях и вызывать совокупность генетических нарушений известных как синдром гибридного дисгенеза (Kidwell et al., 1977; Bregliano et al., 1980). Эти нарушения включают повышенную частоту мутаций, хромосомных аберраций и рекомбинаций, температуро-зависимую стерильность (Bregliano et al., 1980). К настоящему времени описано три независимые системы гибридного дисгенеза, в которых проявление перечисленных выше нарушений обусловлено активностью мобильных элементов I, P и hobo (Bregliano et al., 1980). Все три системы имеют сложные механизмы регуляции активности мобильных генетических элементов. Эти механизмы напрямую связаны с процессами транспозиции и репарации, поэтому реагируют на действие факторов, влияющих на эти процессы. Исследование вопроса функционирования систем гибридного дисгенеза в неблагоприятных условиях окружающей среды может иметь большое теоретическое и прикладное значение (Иващенко и др., 1990).
P-M система гибридного дисгенеза была открыта в середине 70-х годов (Kidwell et al., 1977) и на сегодняшний день является наиболее изученной по отношению к H-E и I-R системам. За возникновение этой системы гибридного дисгенеза отвечает мобильный элемент P (Engels, 1989). В соответствии с наличием в геноме P-элементов различают несколько типов линий Drosophila melanogaster (Raymond et al., 1991). P-линии содержат 30-60 копий P-элемента, одна треть из которых состоит из полных P-элементов, а две трети из дефектных (O'Hare, Rubin, 1983; O'Hare et al., 1992). Эти линии имеют P-цитотип. В геноме M-линий отсутствуют P-элементы, и они имеют M-цитотип. Синдром гибридного дисгенеза наблюдается только при скрещивании самок из M-линий (Maternal) с самцами из P-линий (Paternal), однако поскольку P-цитотип наследуется по материнской линии, потомство от обратных скрещиваний между P-самками и M-самцами обычно нормальное. Дополнительно различают также M' и Q линии. M' или псевдо-M линии имеют в геноме множество дефектных P-элементов, однако, характеризуются наличием слабого потенциала репрессии (M-цитотип) (Simmons et al., 1987). Некоторые M'-линии способны индуцировать определенные аспекты гибридного дисгенеза. Q-линии также несут в геноме дефектные элементы и, подобно P-линиям, имеют P-цитотип. Q-линии обладают способностью индуцировать дисгенез в скрещиваниях с истинными M-линиями (Simmions et al., 1985).
В настоящее время P-элемент подробно изучен на молекулярном уровне, что позволяет нам более четко представить его функции в P-M системе гибридного дисгенеза. Как уже было отмечено, в геноме Drosophila melanogaster встречаются структурно и функционально гетерогенные P-элементы (O'Hare, Rubin, 1983). Полноразмерный P-элемент имеет длину 2907 п.н. и характеризуется наличием терминальных инвертированных повторов размером 31 п.н. и субтерминальными инвертированными повторами размером 11 п.н., которые необходимы для его перемещения (O'Hare, Rubin, 1983). Внутренняя часть содержит небольшой инвертированный повтор с неизвестными функциями и ген транспозазы, состоящий из четырех экзонов и трех интронов (Engels, 1989). Ген транспозазы кодирует белок необходимый для перемещения P-элемента, поэтому полноразмерный P-элемент сам контролирует свое перемещение, т. е. является автономным (Rio et al., 1986). Кроме полноразмерных P-элементов, в геноме различных линий Drosophila melanogaster встречаются дефектные копии (O'Hare et al., 1992). К ним относится KP элемент, который имеет делецию в центральном участке, захватывающую 808-2560 нуклеотиды (Black et al., 1987), элементы A12 и D50 (Engels, 1989; Rasmusson et al., 1993). Дефектные P-элементы не способны к синтезу транспозазы, но благодаря сохранности интактных терминальных и субтерминальных последовательностей, они могут перемещаться с использованием транспозазы полноразмерных элементов (Engels, 1989).
На сегодняшний день известно два типа регуляции активности P-элемента (Engels, 1989). Первый тип регуляции ограничивает активность P-элемента только клетками зародышевой линии, второй тип регулирует активность P-элемента в дисгенных скрещиваниях. Ограничение активности P-элемента только клетками зародышевой линии является следствием регулируемого сплайсинга мРНК (Laski et al., 1986). В зародышевых клетках сплайсируются три интрона, что ведет к образованию транспозазы. В соматических тканях третий интрон не удаляется и, вследствие присутствия в этом интроне стоп-кодона, образуется усеченный белок, который действует как репрессор (Robertson, Engels, 1989). Тканеспецифичный сплайсинг является следствием действия соматических факторов, ингибирующих сплайсинг третьего интрона (Siebel et al., 1992).
Механизм регуляции транспозиций P-элемента в дисгенных скрещиваниях еще не понят полностью. На непродолжительный срок (несколько поколений) эта регуляция наследуется по материнской линии, но на более длительный срок определяется хромосомно, самими P-элементами. Такой тип регуляции в клетках зародышевой линии именуется P-цитотипом, ее отсутствие обозначается как M-цитотип. Модель, предложенная для объяснения принципов детерминации и наследования P-цитотипа, основана на альтернативном сплайсинге пре-мРНК P-элемента на уровне 2-3 интрона. Этот альтернативный сплайсинг определяет продукцию транспозазы или репрессора. Сплайсинг зависит от концентрации пре-мРНК P-элемента, будучи менее эффективен, когда концентрация низкая (O'Hare et al., 1992). В P-цитотипе промотор P-элемента репрессирован, что ведет к низкой концентрации пре-мРНК и к синтезу репрессорного белка. Наоборот, в дисгенных условиях P-промотор не репрессирован, что ведет к высокой концентрации пре-мРНК и к синтезу транспозазы. Эта модель была первоначально подтверждена генетическими методами (Lemaitre et al., 1993) и затем данными молекулярного анализа (Roche et al., 1995). Репрессионная способность P-элемента зависит также от структуры и положения в геноме (Ronsseray et al., 1997).
Высокий уровень регуляции перемещений P-элемента предполагает высокую чувствительность P-M системы гибридного дисгенеза к действию ДНК-повреждающих факторов и к нарушениям в процессах репарации. Действительно, это подтверждается многочисленными экспериментальными факторами. Показано, что облучение влияет на эффекты транспозиций P-элемента в условиях гибридного дисгенеза, что повышает выход рецессивных и доминантных летальных мутаций (Margulies et al., 1986, 1987). Наблюдаемый при этом эффект синергичного действия облучения и активности транспозона, вероятнее всего, связан с индукцией этими двумя факторами однотипных повреждений ДНК, а именно, двунитевых разрывов. Способность P-элемента вызывать такие серьезные повреждения ДНК, а также активность на премейотических стадиях развития яйцеклеток, обусловливает повышенный интерес к вопросу о функционировании P-M системы гибридного дисгенеза в условиях нарушения репарации. Особое значение могут иметь мутации в генах mei-9+ и mei-41+, контролирующих одновременно мейотическую рекомбинацию и репарацию (Sekelsky et al., 1998). При исследовании системы транспозиций в условиях гибридного дисгенеза у линий с мутациями генов репарации mei-9+, mei-41+ и mus101+ не наблюдали видимого эффекта на уровень рекомбинации у самцов и инсерционный мутагенез (Slatko et al., 1984). Мутации mei-41 и mus101 имели продленный эффект на нерасхождение хромосом и эмбриональную смертность, усиливая их, присутствие мутации mei-41 значительно снижало появление хромосом с P-элементами. Эти эффекты наблюдали только у мух с M-цитотипом, что демонстрирует их обусловленность синдромом гибридного дисгенеза. На основании этих результатов сделан вывод, что дефекты в процессе пострепликативной репарации (мутация mei-41) усиливают те из проявлений гибридного дисгенеза, которым сопутствуют события клеточной гибели и доминантной летальности (Slatko et al., 1984). Однако, ни пострепликативная репарация (мутация mei-41) ни эксцизионная репарация (мутация mei-9) не влияют на уровень рекомбинации у самцов и частоту инсерций. В то же время показано, что в присутствии мутаций mei-9 и mei-41 резко повышается уровень индуцированных гибридным дисгенезом видимых мутаций, в том числе, в локусе singed (Eeken, Sobels, 1981). Важность путей пострепликативной и эксцизионной репарации для репарации повреждений, индуцируемых при транспозициях P-элемента, подтверждается исследованием уровня стерильности в скрещиваниях с использованием линий mei-9 и mei-41 (Margulies, 1990). Показано, что при скрещивании мух, имеющих нарушение системы репарации, с мухами, имеющими активные P-элементы в геноме, наблюдается высокий уровень термочувствительной стерильности, низкая плодовитость и преждевременное старение клеток зародышевой линии самцов (Margulies, 1990).
Следующая из рассматриваемых систем гибридного дисгенеза связана с активностью hobo-элемента (Yannopoulos et al., 1987). Hobo-элемент перемещается через образование ДНК-посредника и принадлежит к семейству hobo-Ac-Tam3 (hAT) (Calvi et al., 1991). Полный hobo-элемент имеет длину 2959 п.н. (Blackman et al., 1989). Он несет два инвертированных концевых повтора по 12 п.н. и образует дупликацию в сайте инсерции размером 8 п.н. (McGinnis, 1983). Транспозиции hobo-элемента в H-E системе гибридного дисгенеза специфичны для клеток зародышевого пути, хотя может наблюдаться слабая активность hobo в соматических тканях эмбрионов (Calvi, Gelbart, 1994; Handler, Gomez 1995). Подобно P-элементу, активность hobo ограничена зародышевыми клетками из-за отсутствия транспозазы в соматических тканях. Однако, в отличие от P-элемента, тканеспецифическая транспозиция hobo регулируется выработкой транспозазы на уровне транскрипции (Calvi, Gelbart, 1994).
Классификация линий в H-E системе гибридного дисгенеза основана на присутствии или отсутствии полноразмерного hobo-элемента. Используя этот критерий, линии классифицируются как: (1) H-линии (Hobo), когда молекулярными методами определяют наличие полноразмерных hobo-элементов; они также содержат элементы с внутренней делецией; (2) DH-линии (Deleted Hobo), когда определяются только делетированные элементы; (3) E-линии (Empty), которые не имеют ни полных, ни делетированных копий элемента hobo. В дополнение, линии могут быть классифицированы по их способности индуцировать гонадную атрофию (Bazin, Higuet, 1996). Дисгенная стерильность зависит не только от H-, но и от E-линий. Для H-E системы гибридного дисгенеза характерно также отсутствие корреляции между различными дисгенными событиями (Bazin, Higuet, 1996).
Механизмы регуляции транспозиций hobo-элементов несколько отличаются от механизмов регуляции активности P-элементов, однако, схожесть строения и функций этих элементов может предполагать изменение функционирования hobo-элементов в H-E системе гибридного дисгенеза в ответ на действие ионизирующего облучения, как это показано для P-M системы. В пользу предположения о респонсивности hobo-элементов на действие внешних факторов свидетельствуют также данные об изменении характеристик в H-E системе гибридного дисгенеза у некоторых длительно селектируемых по адаптивным признакам линий Drosophila melanogaster (Кайданов и др., 1994). Согласно этим данным, низкоактивные линии характеризуются повышенной способностью индуцировать дисгенную стерильность и пониженной способностью репрессировать гибридный дисгенез. Линии с высокими адаптивными показателями не индуцируют дисгенную стерильность, но существенно репрессируют ее. Возможно, что эти различия определяются разным составом фракций hobo-элемента и разной локализацией его копий в геноме. Выявляется достоверная корреляция между половой активностью самцов соответствующих линий и их репрессионным потенциалом. Низкоактивные линии характеризуются исключительно высокой частотой спонтанного мутирования (высокой частотой возникновения рецессивных сцепленных с полом и аутосомных мутаций, поздних эмбриональных леталей). В основе этого явления лежит механизм перемещения по геному мобильных hobo-элементов. Низкоактивная линия содержит полноразмерные копии hobo-элементов, способных к синтезу транспозазы и транспозициям. У этой линии обнаружены закономерные изменения в числе и локализации в геноме ретротранспозонов, которые связаны с приспособленностью линий. Возможно, что мобильные генетические элементы являются составной частью генотипа селектируемых линий, обеспечивающих стратегию вредных последствий отбора и инбридинга. И хотя дестабилизация hobo-элемента сама по себе не вызывает изменения приспособленности линии, выявляется достоверная корреляция между половой активностью самцов соответствующих линий и их репрессионным потенциалом (Кайданов и др., 1994). Это предполагает возможную роль H-E системы гибридного дисгенеза в формировании генетических механизмов связанных с приспособленностью к внешним условиям и уровнем генетической изменчивости.
I-R система гибридного дисгенеза обусловлена активностью I-элемента (Bucheton et al., 1984), который относится к классу ретропозонов или LINE-подобных элементов (Fawcett et al., 1986; Pelisson et al., 1991). Полноразмерный I-элемент имеет длину 5371 п.н. Перемещение I-элемента происходит через образование РНК-посредника с использованием обратной транспозазы, которая кодируется самим элементом (Chaboissier et al., 1990; Fawcett et al., 1986). По отношению к I-R системе гибридного дисгенеза линии Drosophila melanogaster подразделяются на два типа. I-линии (Inducer) или индукторные и R-линии (Reactive) или реактивные. В геноме I-линий содержится 10-15 копий полноразмерных I-факторов, которые распределены по всем хромосомам (Bucheton et al., 1984). Активация I-элемента происходит в скрещиваниях самцов из I-линий, которые имеют I-цитотип с самками из линий с R-цитотипом, в скрещиваниях I-самок с R-самцами I-элемент не активируется (Bucheton et al., 1984). Дисгенные нарушения наблюдаются только в яичниках у гибридных самок, в то время как у гибридных самцов таких нарушений не наблюдается. Регуляция активности I-фактора в клетках зародышевой линии осуществляется на уровне инициации транскрипции или стабильности РНК (Chaboissier et al., 1990). Частота транспозиций I фактора в дисгенных скрещиваниях регулируется уровнем реактивности R-самок (Udomkit et al., 1996). В соответствии с этим критерием различают линии со слабым, средним или сильным уровнем реактивности. Уровень реактивности определяется клеточным состоянием в зрелом ооците R-самки и наследуется преимущественно по материнской линии. Уровень реактивности связан с механизмами репарации и рекомбинации и усиливается при действии ДНК повреждающих факторов. Так показано, что действие ингибиторами синтеза ДНК и гамма лучами усиливает уровень реактивности сходным образом (Bregliano et al., 1995). В то же время, уровень реактивности коррелирует с частотой кроссинговера и эффективностью репарации (Laurençon et al., 1997). Это позволяет предположить, что уровень реактивности является одним из проявлений единой индуцибельной репарационно-рекомбинационной системы (Bregliano et al., 1995). Биологическая роль которой может быть аналогична SOS-ответу у бактерий, и заключаться в модификации уровня изменчивости в ответ на изменение условий окружающей среды (Bregliano et al., 1995). Предложено называть эту систему VAMOS (от англ. variability modulation system, система
модуляции изменчивости) (Laurençon et al., 1997). Молекулярные механизмы, участвующие в формировании этой системы еще не выяснены, однако наиболее вероятным представляется участие генов, которые одновременно контролируют процессы рекомбинации и репарации (Laurençon, Bregliano, 1995). Из известных на сегодняшний день генов в определении уровня реактивности наиболее вероятно участие генов mei-9+ и mei-41+ (Laurençon, Bregliano, 1995). Дальнейшее исследование роли, которую играет VAMOS в контроле генетической изменчивости при неблагоприятных условиях окружающей среды, может существенно прояснить работу молекулярных механизмов адаптации.
Дисгенные нарушения в рассмотренных системах гибридного дисгенеза в основном обусловлены транспозициями и эксцизиями мобильных элементов в развивающихся зародышевых клетках. Высокая частота хромосомных перестроек и рекомбинации у самцов происходят преимущественно в сайтах инсерции МГЭ (Engels, Preston, 1984; Sved et al., 1990). Повышенный уровень мутаций происходит от инсерционных мутаций и других индуцированных транспозициями МГЭ изменений в геноме.
Явление гонадной атрофии
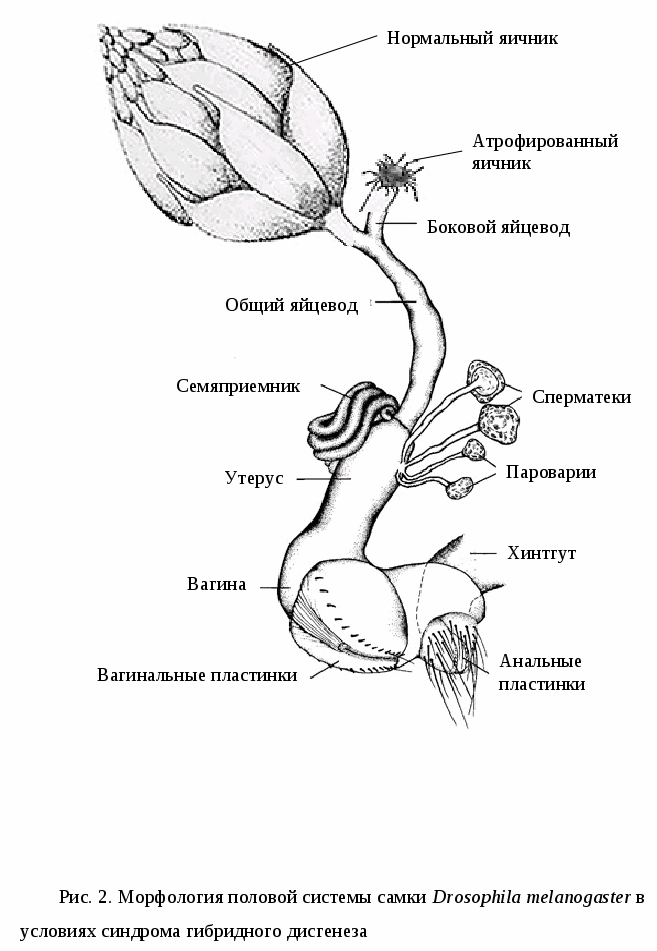
Активация мобильных элементов в системах гибридного дисгенеза вызывает, среди прочих нарушений, особый вид стерильности гибридов, которая обусловлена недоразвитием гонад (Bregliano et al., 1980). Дисгенная стерильность по-разному проявляется в трех системах гибридного дисгенеза. P-M дисгенез приводит к недоразвитию яичников у гибридных самок и самцов (GD-стерильность) (рис. 1) (Kidwell et al., 1977; Schaeffer et al., 1979), в I-R системе, не происходит изменения морфологии гонад, но увеличивается уровень дефектных яиц и частота гибели эмбрионов (SF-стерильность) (Pelisson, 1979). Активация hobo элементов в H-E системе гибридного дисгенеза приводит как к недоразвитию гонад у самок и самцов первого поколения, так и к высокому уровню доминантных леталей среди отложенных яиц.
Стерильность является следствием потери зародышевых клеток на стадиях раннего эмбриогенеза и личинки (Niki, Chigusa, 1986). Для P-M гибридного дисгенеза гибель зародышевых клеток
значительно усиливается при повышении температуры до 29 С (Simmons et al., 1987).
Атрофия гонад одно из характерных и интенсивно изучаемых аспектов P-M и H-E систем гибридного дисгенеза. GD-стерильность является следствием вымирания клеток в примордиальной зародышевой линии возможно из-за хромосомных разрывов, опосредованных активностью P-элемента (Niki, Chigusa, 1986). Однако ни кинетика гибели зародышевых клеток, ни число клеток, которые должны быть элиминированы для появления GD-стерильности не известно. Первые признаки стерильности появляются уже у 5-6 часовых эмбрионов. Максимальный уровень гибели клеток наблюдается на личиночной стадии развития, когда зародышевые клетки испытывают экспоненциальный рост, но некоторые умирающие клетки были обнаружены до деления клеток зародышевой линии (Niki, Chigusa, 1986). Механизм гибели клеток может быть обусловлен разрывами хромосом, имеющими летальный эффект. Следствием этого является отсутствие половых клеток в яичниках и семенниках и общее недоразвитие их у взрослых гибридных самок и самцов. Гибриды могут быть полностью стерильны, если редуцированы обе железы и частично фертильны, если атрофирован один семенник или яичник (рис.2) (Kidwell et al., 1977). Не атрофированные гонады у дисгенных гибридов часто отстают в своем развитии и содержат меньшее число яиц или сперматоцитов по сравнению с недисгенными особями (Ashburner, 1989). Степень выраженности гонадной атрофии сильно зависит от температуры, при которой идет развитие гибридов. В отношении Р-М системы атрофия наиболее значительна при 29 С у самок и при 27 С у самцов, а при 24 С и ниже практически отсутствует (Kidwell et al., 1977; Engels, Preston, 1984). Для Н-Е системы характерна наиболее сильная атрофия при 25 С и наименьшая при 29 С (Stamatis et al., 1989). При более низких температурах АГ наблюдается в меньшей степени. Количество и размерность копий Р- и hobo-элементов так же сильно влияют на способность линий индуцировать гибридный дисгенез, в связи с чем, частота гонадной атрофии может изменяться от нескольких до ста процентов (Yannopoulos et al., 1987; Kidwell et al., 1988; Rasmusson et al., 1993).
Активность мобильных элементов в P-M и H-E системах гибридного дисгенеза обусловливает мутабильность некоторых нестабильных локусов. В P-M системе наибольшую известность получил локус singed-weak (Engels, 1989), в H-E системе локус vgal (Bazin et al., 1993) и сконструированный маркерный элемент h(w+) (Calvi, Gelbart, 1994). Аллель snw обусловлена инсерцией двух дефектных P-элементов и делеция одного или другого из этих элементов в присутствии транспозазы полноразмерных элементов приводит к появлению соответствующих производных аллелей sne и sn(+).Повышенная мутабильность локуса vgal обусловлена активацией дефектных копий hobo в присутствии активных элементов этого семейства. Генетически сконструированный маркерный элемент h(w+) представляет hobo-элемент с встроенным геном mini-white, который определяет оранжевую окраску глаз (Calvi, Gelbart, 1994). Маркерная white линия несет в X-хромосоме два сконструированных элемента h(w+). Это обусловливает оранжевый фенотип цвета глаз, но при эксцизии одного из элементов цвет становится менее выраженным (Calvi, Gelbart, 1994). Линии с нестабильными аллелями широко используются при оценке активности P и hobo-элементов в дисгенных скрещиваниях, и частота мутирования этих аллелей служит дополнительным количественным критерием.
Таким образом, контроль активности мобильных генетических элементов в системах гибридного дисгенеза тесно взаимосвязан с механизмами транспозиций и репарации генетических повреждений. Это обусловливает чувствительность гибридного дисгенеза к действию внешних факторов и его модификацию различным генетическим фоном. Существующие предпосылки позволяют рассматривать синдром гибридного дисгенеза не только как показатель активности некоторых семейств мобильных генетических элементов, но и как целостную генетическую систему, обеспечивающую контроль генетической изменчивости генотипа в неблагоприятных условиях.