Развитие жизни на Земле в протейскую эру (работа 1)
План:
Введение.
А.Предисловие
Б. Масштабы Геологического Времени
В.Основные Подразделения Геологической Истории Земли
Развитие жизни в криптозое.
А. Предисловие
Б. Отложения Криптозоя
В. Резкое Увеличение Богатства Ископаемой Фауны
Заключение.
Введение.
Эволюционное развитие организмов исследуется целым рядом наук, рассматривающих разные аспекты этой фундаментальной проблемы естествознания. Ископаемые остатки животных и растений существовавших на Земле в прошедшие геологические эпохи, изучает палеонтология, которую и следует поставить на первое место среди наук, непосредственно связанных с исследованием эволюции органического мира. Изучая остатки древних форм и сопоставляя их с ныне живущими организмами, палеонтологи реконструируют облик, образ жизни и родственные связи вымерших животных и растений, определяют время их существования и на этой основе воссоздают филогенез — èñòîðèческую преемственность разных групп организмов, их эволюционную историю. Однако в решении этих сложных проблем палеонтология должна опираться на данные и выводы многих других наук, относящихся к кругу биологических, геологических и географических дисциплин (сама палеонтология, изучая ископаемые остатки организмов, находится как бы на стыке биологии и геологии). Для понимания условий жизни древних организмов, определения времени их существования и закономерностей перехода их остатков в ископаемое состояние палеонтология использует данные таких наук, как историческая геология, стратиграфия, палеогеография, палеоклиматология и др. С другой стороны, для анализа строения, физиологии, образа жизни и эволюции вымерших форм необходимо опираться на детальное знание соответствующих сторон организации и биологии ныне существующих организмов. Такое знание дают прежде всего работы в области сравнительной анатомии. Одной из основных задач сравнительной анатомии является установление гомологии органов и структур у разных видов. Под гомологией понимается сходство, основанное на родстве; наличие гомологичных органов доказывает прямые родственные связи обладающих ими организмов (как предков и потомков или как потомков общих предков). Гомологичные органы состоят из сходных элементов, развиваются из сходных эмбриональных зачатков и занимают сходное положение в организме. Развивающаяся ныне функциональная анатомия, а также сравнительная физиология дают возможность подойти к пониманию функционированля органов у вымерших животных. В анализе строения, жизнедеятельности и условий существования вымерших организмов ученые опираются на принцип актуализма, выдвинутый геологом Д. Геттоном и глубоко разработанный одним из крупнейших геологов XIX â. — ×. Лайелем. Ñîãëàñíî ïðèíöèïó àêòóàëèçìà, çàêîíомерности и взаимосвязи, наблюдаемые в явлениях и объектах неорганического и органического мира в дате время, действовали и в прошлом (а отсюда “настоящее есть ключ к познанию прошлого”). Конечно, этот принцип является допущением, но, вероятно, он верен в большинстве случаев (хотя всегда нужно принимать во внимание возможность какого-то своеобразия в протекании тех или иных процессов в прошлом по сравнению с современностью). Палеонтологическая летопись, представленная ископаемыми остатками вымерших организмов, имеет пробелы, иногда очень крупные, обусловленные специфичностью условий захоронения остатков организмов и крайней редкостью совпадения всех необходимых для этого факторов. Для воссоздания филогенеза организмов во всей полноте, для реконструкции многочисленных “недостающих звеньев” па родословном древе (графическом изображении филогенеза) чисто палеонтологические данные и методы оказываются во многих случаях недостаточными. Здесь приходит на помощь так называемый метод тройного параллелизма, введенный в науку известным немецким ученым Э. Геккелем è îñíîâàнный на сопоставлении палеонтологических, сравнительно-анатомических и эмбриологических данных. Геккель исходил из сформулированного им “основного биогенетического закона”, гласящего, что онтогенез (индивидуальное развитие организма) есть сжатое и сокращенное повторение филогенеза. Следовательно, изучение индивидуального развития современных организмов позволяет в какой-то мере судить о ходе эволюционных преобразований их далеких предков, в том числе и не сохранившихся в палеонтологической летописи. Позднее А. Н. Северцов â ñâîåé òåîðèè филэмбриогенезов показал, что соотношение онтогенеза и филогенеза гораздо сложнее, чем считал Э. Геккель. В действительности не филогенез творит индивидуальное развитие (новые эволюционные приобретения удлиняют онтогенез, прибавляя новые стадии), как полагал Геккель, а, наоборот, наследственные изменения хода онтогенеза приводят к эволюционным перестройкам (“филогенез есть эволюция онтогенеза”). Лишь в некоторых частных случаях, когда эволюционная перестрой ка какого-либо органа происходит посредством изменения поздних стадий его индивидуального развития, т. е. новые признаки формируются в конце онтогенеза (такой способ эволюционной перестройки онтогенеза Северцов назвал анаболией), действительно наблюдается такое соотношение между онтогенезом и филогенезом, которое описывается биогенетическим законом Геккеля. Только в этих случаях можно привлекать эмбриологические данные для анализа филогенеза. Сам А. Н. Северцов дал интересные примеры реконструкции гипо тетических “недостающих звеньев” в филогенетическом древе. Изучение онтогенезов современных организмов имеет еще и другое, не менее важное для анализа хода филогенеза значение: оно позволяет выяснить, какие изменения онтогенеза, “творящие эволюцию”, возможны, а какие — íåò, ÷òî äàåò êëþ÷ ê ïîíèìàíèþ êîíêðåòíûõ ýâîëþöèîííûõ ïåðестроек.
Для понимания сущности эволюционного процесса, для причинного анализа хода филогенеза самое первостепенное значение имеют выводы эволюционистики — íàóêè, называемой также теорией эволюции или дарвинизмом, по имени великого создателя теории естественного отбора Ч. Дарвина. Эволюционистика, изучающая сущность, механизмы, общие закономерности и направления эволюционного процесса, является теоретической базой всей современной биологии. По сути дела, эволюция организмов представляет собой форму существования живой материи во времени, и все современные проявления жизни, на любом уровне организации живой материи, могут быть поняты лишь с учетом эволюционной предыстории. Тем в большей мере важны основные положения теории эволюции для изучения филогенеза организмов.
Перечисленные науки отнюдь не исчерпывают перечень научных дисциплин, причастных к изучению и анализу развития жизни на Земле в прошедшие геологические эпохи. Для понимания видовой принадлежности ископаемых остатков и преобразований видов организмов во времени чрезвычайно важны выводы систематики; для анализа смены фаун и флор в геологическом прошлом — äàнные биогеографии.
Особое место занимают вопросы происхождения человека и эволюции его ближайших предков, имеющей некоторые специфические особенности по сравнению с эволюцией других высших животных, благодаря развитию трудовой деятельности и социальности.
Масштабы Геологического Времени
Изучая эволюцию организмов, необходимо иметь представление о ее ходе во времени, о продолжительности тех или иных ее этапов. Историческая последовательность образования осадочных пород, т. е. их относительный возраст, в данном районе устанавливается сравнительно просто: породы, возникшие позднее, отлагались поверх более ранних пластов.
Соответствие относительного возраста пластов осадочных пород в разных регионах можно определить, сопоставляя сохранившиеся в них ископаемые организмы (палеонтологический метод, основы которого были заложены в конце XVIII — íà÷àëå XIX â. ðàáîòàìè àíãëèéñêîãî ãåîëîãà Ó. Ñìèòà). Îáû÷íî ñðåäè èñêîïàåìûõ îðãàíèçìîâ, õàðàêòåðíûõ äëÿ êàæäîé ýïîõè, óäàåòñÿ âûäåëèòü íåñêîëüêî íàèáîëåå îáû÷íûõ, ìíîãî÷èñëåííûõ è øèðîêî ðàñïðîñòðàíåííûõ âèäîâ - òàêèå âèäû ïîëучили название руководящих ископаемых.
Как правило, абсолютный возраст осадочных пород, т. е. промежуток времени, прошедший со времени их образования, непосредственно установить нельзя. Информация для определения абсолютного возраста содержится в изверженных (вулканических) породах, которые возникают из остывающей магмы.
Абсолютный возраст изверженных пород можно определить по содержанию в них радиоактивных элементов и продуктов их распада. Радиоактивный распад начинается в изверженных породах с момента их кристаллизации из расплавов магмы и продолжается с постоянной скоростью до тех пор, пока все запасы радиоактивных элементов не будут исчерпаны.
Поэтому, определив содержание в горной породе того или иного радиоактивного элемента и продуктов его распада и, зная скорость распада, можно достаточно точно (с возможностью ошибки около 5%) âû÷èñëèòü àáñîëþòíûé âîçðàñò äàíной породы. Для осадочных пород приходится принимать приблизительный возраст по отношению к абсолютному возрасту слоев вулканических пород. Длительное и кропотливое изучение относительного и абсолютного возраста горных пород в разных регионах земного шара, потре-бовавшее напряженной работы нескольких поколений геологов и палеонтологов, позволило наметить основные вехи геологической истории Земли. Границы между этими подразделениями соответствуют разного рода изменениям геологического и биологического (палеонтологического) характера. Это могут быть изменения режима осадконакопления в водоемах, приводящие к формированию иных типов осадочных пород, усиление вулканизма и горообразовательные процессы, вторжение моря (морская трансгрессия) благодаря опусканию значительных участков континентальной коры или повышению уровня океана, существенные изменения фауны и флоры. Поскольку подобные события происходили в истории Земли нерегулярно, продолжительность различных эпох, периодов и эр различна. Обращает на себя внимание огромная длительность древнейших геологических эр (археозойской и протерозойской), которые к тому же не разделены на меньшие временные промежутки (во всяком случае, нет еще общепринятого подразделения). Это обусловлено в первую очередь самим фактором времени — äðåâíîñòüþ îòëîæåíèé àðõåозоя и протерозоя, подвергшихся за свою длительную историю значительному метаморфизму и разрушению, стершим существовавшие когда-то вехи развития Земли и жизни. Отложения архейской и протерозойской эр содержат чрезвычайно мало ископаемых остатков организмов; по этому признаку археозой и протерозой объединяют под названием “криптозой” (этап скрытой жизни) противопоставляя объединению трех последующих эр — “ôàíåðîçîé” (ýòàï ÿâíîé, íàáëþäàåìîé æèçíè). Âîçðàñò Çåìëè îïðåäåëÿåòñÿ ðàçëè÷íûìè ó÷åíûìè ïî-ðàçíîìó, íî ìîæно указать на приближенную цифру 5 ìëðä. ëåò.
Развитие жизни в криптозое.
Эры, относящиеся к криптозою, — археозойская и протерозойская — вместе продолжались более 3,4 ìëðä. ëåò; ò. î. криптозой составляет не менее 7/8 âñåй геологической истории. Однако в отложениях криптозоя сохранилось чрезвычайно мало ископаемых остатков организмов, поэтому представления ученых о первых этапах развития жизни в течение этих огромных промежутков времени в значительной степени гипотетичны.
Отложения Криптозоя
Древнейшие остатки организмов были найдены в осадочных толщах Родезии, имеющих возраст 2,9—3,2 ìëðä. ëåò. Òàì îáíàðóæåны следы жизнедеятельности водорослей (вероятно, сине-зеленых), что убедительно свидетельствует, что около 3 ìëðä. ëåò íàçàä íà Çåìëå óæå ñóùåñòâîâàëè ôîòîñèíòåçèðóþùèå îðãàíèçìû — âîäîðîñëè. Î÷евидно, появление жизни на Земле должно было произойти значительно раньше,— ìîæåò áûòü, 3,5— 4 ìëðä. ëåò íàçàä. Íàèáîëåå èçвестна среднепротерозойская флора (нитчатые формы длиной до нескольких сотен микрометров и толщиной 0,6—16 ìêì, èìåþùèå ðàçëè÷íîå ñòðîåíèå, îäíîêëåòî÷íûå ìèêðîîðãàíèçìû (Ðèñ.1), äèàìåòðîì 1—16 ìêì, òàêæå ðàçëè÷íîãî ñòðîåíèÿ), îñòàòêè êîòîðîé áûëè îáíàðóæåíû â Êàíàäå —â êðåìíèñòûõ ñëàíöàõ íà ñåâåðíîì áåрегу озера Верхнего. Возраст этих отложений составляет около 1,9 ìëðä. ëåò.
Рис. 1
Пример найденных ископаемых останков простейших

В осадочных породах, образовавшихся в промежутке времени между 2 è 1 ìëðä. ëåò íàçàä, ÷àñòî âñòðå÷àþòñÿ ñòðîìàòîëèòû, ÷òî ãоворит о широком распространении и активной фотосинтезирующей и рифостроительной деятельности сине-зеленых водорослей в этот период.
Следующий важнейший рубеж в эволюции жизни документируется рядом находок ископаемых остатков в отложениях, имеющих возраст 0,9—1,3 ìëðä. ëåò, ñðåäè êîòîðûõ íàéäåíû ïðåêðàсной сохранности остатки одноклеточных организмов размером 8—12 ìêì, â êîторых удалось различить внутриклеточную структуру, похожую на ядро; îáíàðóæåíû òàêæå ñòàäèè äåëåíèÿ îäíîãî èç âèäîâ ýòèõ îäíîêëеточных организмов, íàïîминающие стадии митоза — ñïîñîáà äåëåíèÿ ýóêàðèîòè÷еских (т. е. имеющих ядро) клеток.
Если интерпретация описанных ископаемых остатков правильна, это означает, что около 1,6—1,35 ìëðä. ëåò íàçàä ýâîëþöèÿ îðãàíèçмов прошла важнейший рубеж — áûë äîñòèãíóò óðîâåíü îðãàíèçàöèè ýукариот.
Первые следы жизнедеятельности червеобразных многоклеточных животных известны из позднерифейских отложений. В вендское время (650—570 ìëí. ëåò íàçàä) ñóùåñòâîâàëè óже разнообразные животные, вероятно, принадлежавшие к различным типам. Íåмногочисленные отпечатки мягкотелых вендских животных известны из разных районов земного шара. Ряд интересных находок был сделан в позднепротерозойских отложениях на территории бывшего СССР.
Наиболее известна богатая позднепротерозойская ископаемая фауна, обнаруженная Р. Сприггом в 1947 ã. â Öåíòðàëüíîé Àâñòðàëèè. Èññëåäîâàâøèé ýòó óíèêàëüíóþ ôàóíó Ì. Глесснер считает, что она включает примерно три десятка видов очень разнообразных многоклеточных животных, относящихся к разным типам (Рис. 2).
Рис. 2. Реконструкция фауны Эдиакары:
1—10—кишечно-полостные; 11—14—плоские и кольчатые черви; 15—16—членистоногие; 17—иглокожие; 18 — шарообразные студенистые организмы.

Большинство форм принадлежит, вероятно, к кишечно-полостным. Это медузоподобные организмы, вероятно “парившие” в толще воды, и прикрепленные к морскому дну полипоидные формы, одиночные или колониальные, напоминающие современных альционарий, или морские перья. Замечательно, что все они, как и другие животные эдиакарской фауны, лишены твердого скелета.
Кроме кишечно-полостных, в кварцитах Паунд, вмещающих эдиакарскую фауну, найдены останки червеобразных животных, причисляемых к плоским и кольчатым червям. Некоторые виды организмов интерпретируются как возможные предки членистоногих. Наконец, имеется целый ряд ископаемых остатков неизвестной таксономической принадлежности. Это указывает на огромное распространение фауны многоклеточных мягкотелых животных в вендское время,
Поскольку вендская фауна столь разнообразна и включает довольно высокоорганизованных животных, очевидно, что до ее возникновения эволюция продолжалась уже достаточно долго. Вероятно, многоклеточные животные появились значительно раньше — ãäå-òî â ïðомежутке 700—900 ìëí. ëåò íазад.
Резкое Увеличение Богатства Ископаемой Фауны
Граница между протерозойской и палеозойской эрами (т. е. между криптозоем и фанерозоем) отмечается поразительным изменением в составе и богатстве ископаемой фауны. Внезапно (другого слова здесь, пожалуй, и не подберешь) после толщ верхнего протерозоя, почти лишенных следов жизни, в осадочных породах кембрия (первого периода палеозойской эры), начиная с самых нижних горизонтов, появляется огромное разнообразие и обилие остатков ископаемых организмов. Среди них остатки губок, плеченогих, моллюсков, представителей вымершего типа археоциат, членистоногих и других групп. К концу кембрия появляются почти все известные типы многоклеточных животных. Этот внезапный “взрыв формообразования” на границе протерозоя и палеозоя — îäíî èç ñàìûõ çàãàäî÷ных, до сих пор полностью не разгаданных, событий в истории жизни на Земле. Благодаря этому начало кембрийского периода является столь заметной вехой, что нередко все предшествующее время в геологической истории (т. е. весь криптозой) именуют "докембрием.
Вероятно, обособление всех основных типов животных произошло в верхнем протерозое, в промежутке времени 600—800 ìëí. ëåò назад. Примитивные представители всех групп многоклеточных животных были небольшими лишенными скелета организмами. Продолжавшееся накопление кислорода в атмосфере и увеличение мощности озонового экрана к концу протерозоя позволили животным, как указано выше, увеличить размеры тела и приобрести скелет. Организмы получили возможность широко расселиться на малых глубинах различных водоемов, что повело к значительному повышению разнообразия форм жизни.
Список литературы.
Бернал Д. Возникновение жизни, М„ 1969.
Борисяк А. А. Из истории палеонтологии (идея эволюции). М.—Л., 1966.
Брукс М. Климаты прошлого. Ì„ 1952.
Варсанофьева В. А. Развитие жизни на Земле. Ì„ 1988.
Голенкин М. И. Победители в борьбе за существование, М., 1959.
Ливанов Н. А. Пути эволюции животного мира. Ì„ 1955.
Ì. Иорданский Н. Н. Развитие жизни на Земле. Ì. 1989.
Рис. 3. Эвриптериды:
а — Eurypterus. á — Mixoptcrus. Îáå ôîðìû èçîáðàæåíû ñî ñïèííîé (ñëåâà) è áрюшной (справа) стороны.
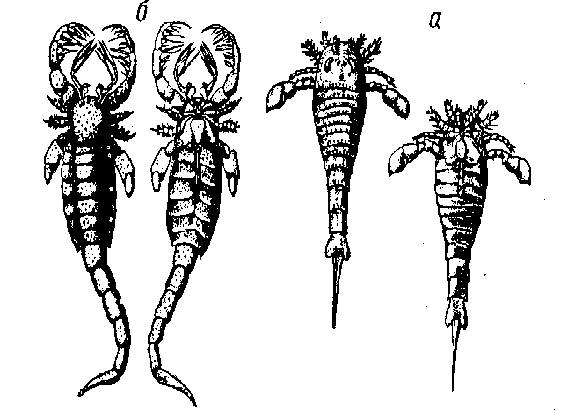
Рис. 4. Ïðèìèòèâíûå áåñ÷åðåïíûå:
1—õîðäà; 2—íåðâíàÿ òðóáêà; 3—ðîòîâîå îòâåðñòèå; 4—ãëîòêà, 5—ýíäîñòèëü; á—íàäæàáåðíàÿ áîðîçäà; 7— æàáåðíûå ùåëè; 8—ïå÷åночный выпоет; 9—êèøå÷íèê; 10 — àнальное отверстие; 11 — миомеры; 12 — áðþøíàÿ àîðòà; 13 — спинная аорта; 14 — æàáåðíûå àðòåðèè.
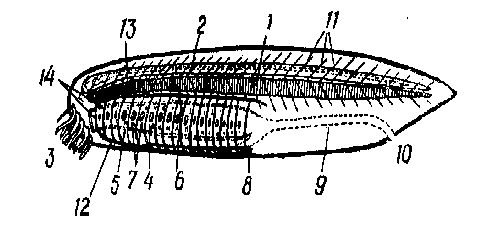
Рис. 5. Ïàíöèðные рыбы (плакодермы): а—ïàíöèðü Dinichthya; á—Pterichthya.

Рис. 6. Акантодия Climatius.

Рис. 7. Палеодиктиоптера (Palaeodiolyoptera).

Рис. 8. Типичный представитель котилозавров

Рис. 9. Ïàðейазавр

Рис. 10. Îñíîâíûå òèïû ÷åðåïà амниот:
а—àíàïñèäíûé; в—ñèíàïñèäíûé; в—äèàïñïäíûé; г—÷åðåïàõè; д—ÿùåðèöû; е— ïòèöû; 1—âèñî÷íûå îêíà; 2—âèñî÷íûе вырезки; 3—çàäíåãëàçíè÷íàÿ кость; 4— ÷åøóé÷àòàÿ êîñòü; 5 — ñкуловая кость; 6 — квадратноскуловая кость: 7 — êâàäðàòíàÿ êîñòü. Стрелками указаны направления филогенетических преобразований.
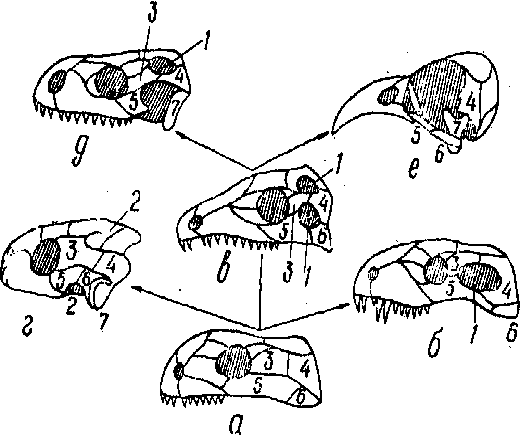
Рис.
11. Ýäàôîçàâð
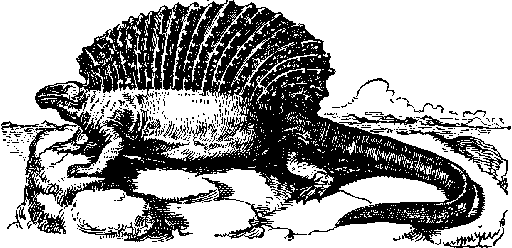
Рис.
12. Èíîñòðàíöåâèÿ

Рис. 13.
Цинодонт
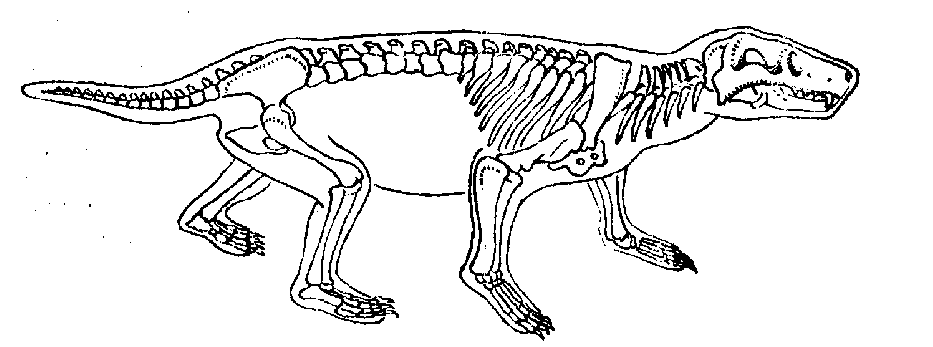
Рис. 14.

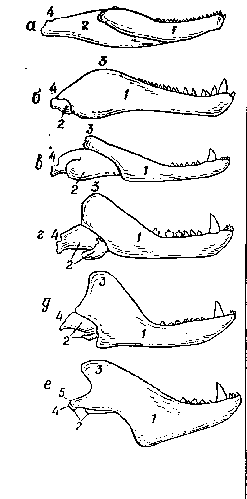 Ðÿä
ïîñëåäîâàòåëüíûõ ïðåîáðàçîâàíèé íèæíåé
÷åëþñòè
Ðÿä
ïîñëåäîâàòåëüíûõ ïðåîáðàçîâàíèé íèæíåé
÷åëþñòè
Рис.15
Эритротерий

Рис. 16
Псевдозухии
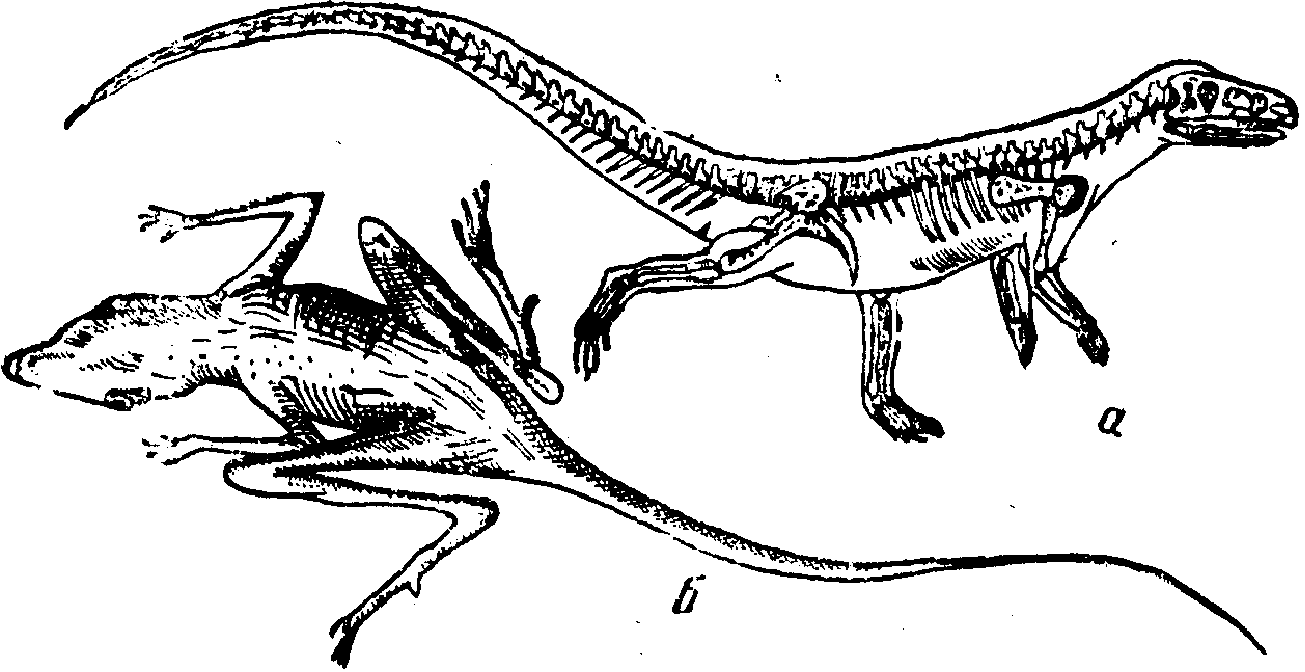
Рис. 17.
Плезиозавр

Рис. 18. Рамфоринх

Рис.
19.
Госперорнис

Рис.
20.
Уинтатерий

Рис. 21, Бронтотерий.
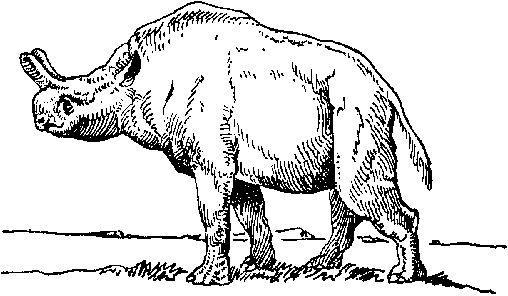
Рис. 22. Индрикотерий.
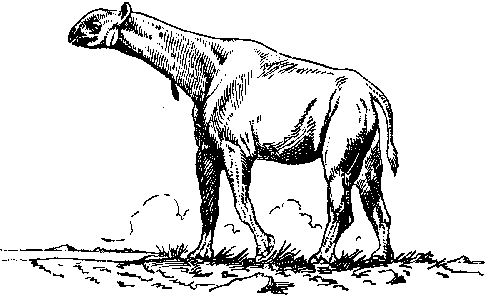
Рис.
23.
Диатрима

Рис. 24. Гиппарион.
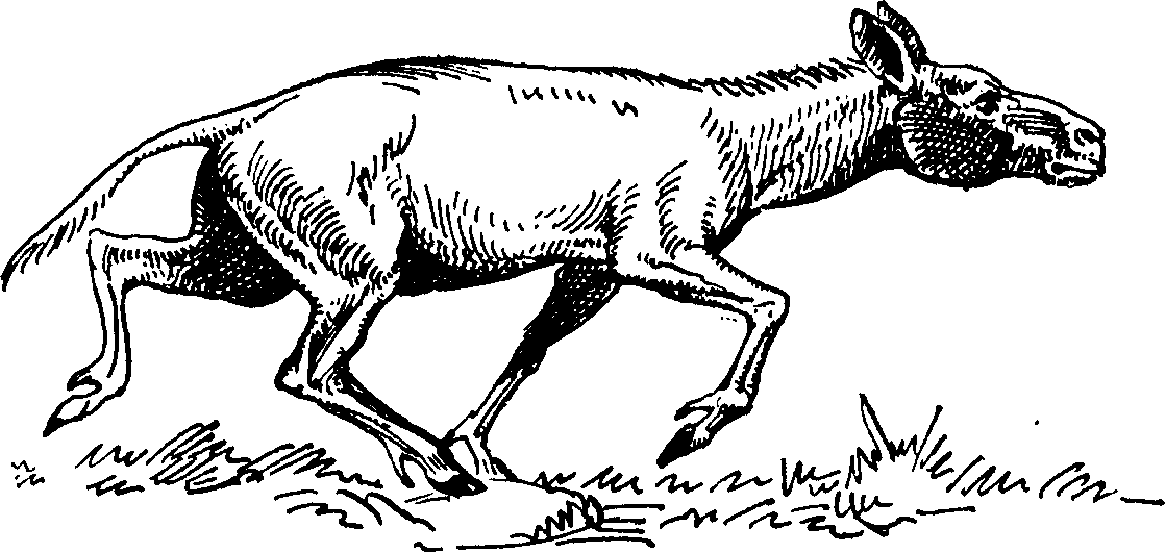
Рис. 25. Большерогий олень.
