Преадаптация
Новый подход в понимании преадаптации
Введение
Лабораторией функциональной ботаники КГУ исследуется вопрос о преадаптации растений [1, 2, 3]. Под термином “преадаптация” мы понимаем всеобщее, фундаментальное свойство живых систем, определяющее их способность к опережающему отражению. В лаборатории разработана классификация преадаптаций, в основу которой положен принцип взаимодействия особи и факторов среды в экологической нише, собраны многочисленные примеры преадаптации растений [4]. Так, очевидно, что преадаптивное значение имеют приобретение растениями неспецифической устойчивости под воздействием стресс-фактора, закаливание, многие фотопериодические реакции, осцилляции и многие другие явления.
Обзор литературы
Термин “преадаптация” введен в биологическую науку французским исследователем Леонардо Кено в 1901 году для обозначения явления предварительного приспособления. Кено понимал преадаптацию как "... безразличные или полуполезные особенности, имеющиеся у вида и способные превращаться в явные приспособления при изменении образа жизни..." Экспериментальные доказательства объективности существования преадаптации были получены в многочисленных работах рада авторов (Сердж, Райн, 1964; Кроу, 1957; Беннет, 1960; Северцов А.Н., 1922; Филюков А.И., 1971). Вместе с тем антидарвиновская трактовка этого явления (преадаптизм), выдвинутая Кено и его последователями, привела к тому, что феномену не уделялось должное внимание, как среди зарубежных, так и среди российских исследователей.
Цель
Целью работы являлось исследование динамики соотношения роста растений, формирования и функционирования фотосинтетического аппарата в онтогенезе георгины культурной в качестве одного из примеров преадаптации.
Методика
В настоящей работе для изучения преадаптивных реакций был избран рост, поскольку он интегрирует все физиологические процессы растения [5].
Опыты проводились в 1995 - 1997 гг. в Ботаническом саду КГУ в условиях мелкоделяночного полевого эксперимента. Для исследований использовали следующие сорта георгины: Залп “Авроры”, Ангажемент, Пионерский галстук, Угунсун Нанс, Фатима, Памяти Гагарина, Вечный огонь, Снежный узор, Ага, Пиегальви ага, Белый кактус, Озирис, Преферанс - отличающиеся темпами роста, характером формирования и габитусом куста, сроками цветения.
Высоту растений измеряли сантиметровой лентой по главному побегу. Площадь листьев определяли весовым методом [6]. Содержание хлорофилла определяли спектрофотометрическим методом в ацетоновом экстракте [7]. Мезоструктуру листьев изучали на прижизненных срезах с помощью светового микроскопа и окулярного микрометра. Объем палисадных клеток рассчитывали с учетом поправочных коэффициентов Ю.Л.Цельникер [8]. Чистую продуктивность фотосинтеза определяли по сухой массе растений [6]. Биологическую активность фитогормонов: цитокининов, гиббереллинов и абсцизовой кислоты - определяли комплексным методом [9]. Содержание фитогормонов в тканях рассчитывали по калибровочным кривым. Замеры и анализы осуществляли с промежутком 5-10 дней в трехкратной биологической и пятикратной аналитической повторностях.
Результаты и обсуждение
Поскольку задачей настоящего исследования было сопоставление онтогенетической динамики изучаемых процессов, мы полагаем корректным представление данных в виде среднесуточных изменений анализируемых параметров. Характер изменений изученных процессов в онтогенезе оказался одинаковым у всех сортов. Однако скорость этих изменений у разных сортов заметно варьировала в зависимости от скороцветности. Ниже приведены данные по трем сортам, различающимся сроками зацветания: ранний сорт Ангажемент, среднеранний - Пионерский галстук и средний - Озирис.
Внешним проявлением продукционных процессов является рост. В этой связи нам представляется целесообразным начать рассмотрение вопроса с онтогенетической динамики различных параметров роста георгины. Данные по среднесуточному приросту растений в высоту представлены в графике рис. 1. На графике видно, что темпы роста в высоту изменяются по одновершинной кривой с максимумом в конце фазы бокового ветвления. Однако высота не может служить единственным критерием роста.
Существенными показателями являются также увеличение массы растения и площади листьев. На графиках рис. 2 и 3 отражен среднесуточный прирост сырой и сухой массы георгины. Как следует из графиков, изменение темпов увеличения массы также описывается одновершинной кривой. Вместе с тем, максимум среднесуточного прироста сырой и сухой массы приходится на конец фазы бутонизации и отстоит от максимального темпа роста растений в высоту на 15-20 дней.
Биологическое значение этого феномена заключается, вероятно, в том, что усиление роста в высоту сопровождается активным формированием листового аппарата и таким образом закладываются структурные предпосылки для будущего усиления фотосинтетической активности при формировании органов семенного и вегетативного размножения. Это предположение подтверждается данными о темпах роста листовой поверхности (рис. 4). Максимальные приросты площади листьев наблюдаются в фазу бокового ветвления, совпадая с максимальными темпами роста стеблей.
В основе биологической продуктивности лежит процесс фотосинтеза. В этой связи следующим этапом исследования было изучение структурных и функциональных параметров фотосинтетической деятельности растений. Для изучения динамики формирования фотосинтетического аппарата мы определяли содержание хлорофилла в листьях и характеристики мезоструктуры листа: мощность слоя палисадной паренхимы, количество хлоропластов в клетках палисадной и губчатой паренхимы. На графике рис. 5 видно, что содержание хлорофилла в листьях растет в первые дни онтогенеза, затем в фазе всходов снижается; а потом опять растет в фазы бокового ветвления и начала бутонизации и постепенно снижается в последующие фазы.
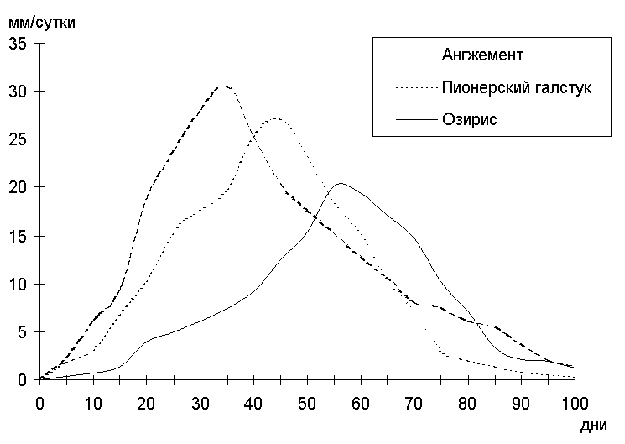
Рис. 1. Среднесуточный прирост в высоту

Рис. 2. Среднесуточный прирост сырой массы

Рис. 3. Среднесуточный прирост сухой массы

Рис. 4. Среднесуточный прирост площади листьев
Аналогично изменяются в онтогенезе георгины параметры мезоструктуры листа (графики рис. 6, 7, 8). Вместе с тем следует отметить, что максимальные значения показателей развития фотосинтетического аппарата в абсолютном выражении наблюдаются в начале онтогенеза.
Иную направленность имеет динамика функционирования фотосинтетического аппарата. Так, начинающийся с первых дней онтогенеза рост чистой продуктивности фотосинтеза (рис. 9) продолжается до конца фазы всходов, затем в фазу бокового ветвления и в начале бутонизации происходит снижение, а в конце фазы бутонизации параметр вновь заметно повышается. В фазы цветения и начала пожелтения нижних листьев чистая продуктивность фотосинтеза резко снижается.

Рис. 5. Среднесуточное изменение содержания хлорофилла в листьях

Рис. 6. Среднесуточное изменение мощности слоя палисадной паренхимы

Рис. 7. Среднесуточное изменение количества хлоропластов в клетках палисадной паренхимы

Рис. 8. Среднесуточное изменение хлоропластов в клетках губчатой паренхимы
Наличие двух максимумов на графиках изменения структурных и функциональных показателей фотосинтетической деятельности, вероятно, связано с необходимостью обеспечения продуктами фотосинтеза интенсивного роста стеблей, листьев и корней в фазу бокового ветвления и процессов формирования органов семенного и вегетативного размножения в фазу цветения.
Поскольку существенным моментом регуляции роста и фотосинтеза является гормональный баланс, нами была изучена динамика содержания эндогенных фитогормонов в листья растений (графики рис. 10-12).
Содержание гиббереллинов в листьях увеличивается вплоть до фазы бутонизации, затем падает. Содержание абсцизовой кислоты и ее производных уменьшается от фазы всходов к фазе бокового ветвления и затем растет вплоть до конца онтогенеза. Активность цитокининов снижается от всходов до бокового ветвления, растет к фазе цветения и падает в фазу начала пожелтения нижних листьев.

Рис. 9. Чистая продуктивность фотосинтеза

Рис. 10. Содержание гиббереллинов в листьях
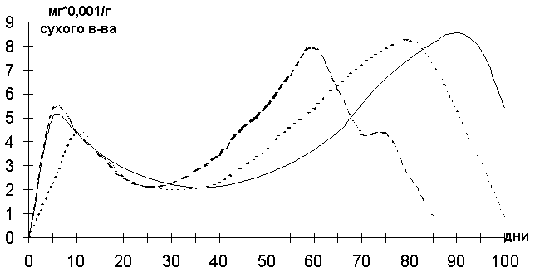
Рис. 11. Содержание цитокининов в листьях

Рис. 12. Содержание абсцизовой кислоты в листьях
Cопоставление полученных данных дает следующие результаты. При сравнении графиков рис. 5, 6, 7, 8 и графика рис. 9 видно, что интенсификация формирования структуры фотосинтетического аппарата опережает в онтогенезе усиление его функциональной активности. Действительно, в первые дни показатели мезоструктуры листа самые высокие, очень резко в начале фазы всходов растет содержание хлорофилла в листьях, а чистая продуктивность фотосинтеза в этот период низка и только начинает расти. Далее в фазе всходов содержание хлорофилла и показатели мезоструктуры снижаются, в то время как чистая продуктивность растет. Максимум как по темпам прироста, так и по абсолютным значениям для показателей развития структуры фотосинтетического аппарата приходится на начало фазы всходов, а для его функциональной активности - на конец этой фазы. Промежуток времени между указанными экстремумами составляет у разных сортов от 10 до 20 дней. В фазе бокового ветвления структурные показатели вновь растут, достигая максимума в начале фазы бутонизации, вместе с тем функциональная активность фотосинтетического аппарата в фазу бокового ветвления и в начале бутонизации снижается, а растет и достигает максимума только в конце фазы бутонизации растений, вновь отставая от максимума структурных показателей на 10-15 дней. В период цветения и начала пожелтения нижних листьев как структурные, так и функциональные показатели снижаются. Таким образом, графики, отражающие изменения структурных и функциональных параметров фотосинтетической деятельности, оказываются сдвинутыми по фазе. Формирование структуры предваряет усиление фотосинтетической функции.
Сравнение динамики ростовых процессов и фотосинтетической деятельности показывает, что в онтогенезе повышение фотосинтетической активности предшествует активизации роста растений. Так, максимальный уровень структурированности фотосинтетического аппарата имеет место в начале фазы всходов, вслед за ним в конце этой фазы достигает максимума фотосинтетическая активность, и только в фазу бокового ветвления наблюдается пик линейного роста стеблей и листьев. Второй максимум темпов формирования элементов фотосинтетического аппарата приходится на начало фазы бутонизации, максимум чистой продуктивности фотосинтеза - на конец этой фазы, а интенсивный рост органов семенного и вегетативного размножения происходит лишь в период цветения.
Оценивая биологическое значение описанных выше закономерностей, мы полагаем возможным интерпретировать их в качестве преадаптации. Четко выраженная временная последовательность проявления экстремумов структурных и функциональных параметров фотосинтетической деятельности позволяет предположить, что заблаговременное повышение уровня структурированности фотосинтетического аппарата подготавливает его к периоду интенсивного функционирования. В противном случае накопление элементов фотосинтетического аппарата, не приводящее к усилению его функции непосредственно в период накопления, теряет биологический смысл. В свою очередь предваряющее интенсивный рост усиление фотосинтетической активности, вероятно, подготавливает растение к предстоящим в период роста метаболическим перестройкам.
Эксперименты показали, что соотношение роста растений, формирования и функционирования фотосинтетического аппарата не зависит от сортовой принадлежности, индивидуальных особенностей, не изменяется в зависимости от погодных условий вегетационного периода. Это позволяет заключить, что обнаруженные закономерности являются генетически детерминированными. В соответствии с разработанной нами классификацией [4] описанная в настоящей работе преадаптация может быть отнесена к алгоритмирующим экспрессивным радиациям. Алгоритмом она является в силу того, что определяет временную последовательность физиологических процессов, и так как не нуждается во внешнем (по отношению к ней) сигнале, может считаться экспрессивной, а в связи с тем, что представляет собой генетическую программу, сформировавшуюся в онтогенезе, то может быть названа радиацией.
Проявившаяся в опытах динамика роста и фотосинтетической деятельности тесно связана с изменениями гормонального баланса. Интенсивный рост происходит на фоне повышенного соотношения гиббереллины/абсцизовая кислота. Снижение этого соотношения сопровождается ослаблением ростовых процессов. Повышению уровня содержания эндогенных цитокининов соответствует усиление дифференциации и функциональной активности фотосинтетического аппарата.
Выводы
Осуществленный в настоящей работе анализ онтогенетической динамики ростовых процессов, формирования и функционирования фотосинтетического аппарата позволяет сделать следующие выводы.
1. Усиление формирования фотосинтетического аппарата предшествует в онтогенезе повышению его функциональной активности.
2. Повышение фотосинтетической активности предваряет интенсификацию линейного роста органов и накопление массы растений.
3. Динамика фотосинтетической и ростовой активности сопровождается соответствующими изменениями гормонального баланса.
4. Полученные данные могут быть интерпретированы как пример преадаптации, являющейся алгоритмирующей экспрессивной радиацией.