Амилолитические препараты
АМИЛОЛИТИЧЕСКИЕ ПРЕПАРАТЫ
Амилолитические препараты широко выпускаются в нашей стране и за рубежом. В основном это крупнотоннажное производство. Амилазы находят применение почти во всех областях, где перерабатывается крахмалсо-держащее сырье. Амилазы используют для осахаривания зернового и картофельного крахмала. Самым большим потребителем амилолитических ферментов является спиртовая и пивоваренная промышленности, где в настоящее время солод (проращенное зерно) успешно заменяется амилолитическими ферментными препаратами.
Источники получения амилаз
Амилазы очень широко распространены в природе. Они синтезируются многими микроорганизмами (бактерии, грибы, актиномицеты, дрожжи), животными и растениями. До развития ферментной промышленности главным промышленным источником получения амилаз в европейских странах было проросшее зерно (солод). Для медицинских целей амилазы получали из животного сырья. В настоящее время главным источником амилаз являются микроорганизмы, особенно бактерии, грибы и реже дрожжи.
Механизм действия и свойства амилаз
Субстратами для действия амилаз являются крахмал, состоящий из амилозы и амилопектина, продукты частичного гидролиза крахмала и гликоген.
Крахмал - растительный полисахарид с очень сложным строением. Это двухкомпонентное соединение, состоящее из 13-30% амилозы и 70-85% амилопектина. Оба компонента неоднородны, их молекулярная масса (М. м.) колеблется в широких пределах и зависит от природы крахмала. Амилоза - это неветвящийся полимер, в котором остатки глюкозы соединены -1, 4-гликозидной связью; степень полимеризации около 2000. В «аномальных» амилозах с одной-двумя -1, 6-связями полимеризация может возрасти до 6000 (рис.1). Амилоза практически не обладает восстанавливающей способностью, так как в каждой молекуле амилозы имеется только одна свободная альдегидная группа.
Молекула амилозы представляет собой растянутую спираль, шаг которой составляет 10,6 А и в каждый виток входит 3 остатка глюкозы. Максималь-
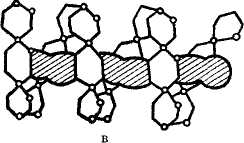
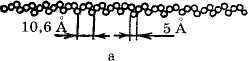
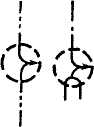
Рис 1. Строение амилозы:
а — амилоза без аномальных отклонений; б - схема возможных ветвлений амилозы; в — спираль амилозы в растворе с заключенными в ее полость молекулами йода.
Концевые звенья с
альдегидными группами 1 разветвление
- 2 разветвления 4 разветвления 8 развет
16 концевых звеньев
64 концевых
звена
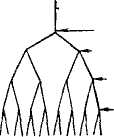
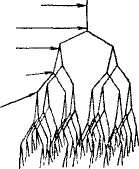
Рис. 2.2. Амилопектин, схема дихотомического деления амилопектина (по К. Мейеру):
а - в плоскости; б - в пространстве.
ная длина молекулы амилозы достигает 7000 А. В растворе спираль сжимается за счет увеличения витка, в котором уже участвует 6 остатков глюкозы. При вхождении молекул йода в спираль амилозы возникает характерный синий цвет. Строго говорить о величине молекулы амилозы нельзя, т. е. даже из одного образца крахмала извлекается амилоза, с величиной молекулы от 500 до 2000 остатков глюкозы. Амилопектин имеет большую молекулярную массу, чем амилоза, и более сложное строение. Это ветвящийся полисахарид. Предполагается, амилопектин ветвится дихотомически, т. е. число концевых звеньев всегда на единицу больше числа звеньев, дающих ветвление, а сумма этих чисел дает общее число звеньев по всей цепи (см. рис. 2.2).
Механизм действия. К группе амилотических ферментов относятся - и -амилазы, глюкоамилаза, пуллуланаза, изоамилаза и некоторые другие ферменты. Амилазы бывают двух типов: эндо- и экзоамилазы. Четко выраженной эндоамилазой является а-амилаза, способная к разрыву внутримолекулярных связей в высокополимерных цепях субстрата.
Глюкоамилаза и -амилаза являются экзоамилазами, т. е. ферментами, атакующими субстрат с нередуцирующего конца.
При изучении механизма действия амилаз имеются определенные сложности, и прежде всего они заключаются в том, что субстрат - крахмал неоднороден и имеет различные характеристики по степени полимеризации гли-козидной цепи и количеству ветвлений.
Реакции, катализируемые амилазами, имеют две стадии: короткую -предстационарную и длительную - стационарную. Во время первой стадии эндоамилаза быстро уменьшает молекулярную массу субстрата, образуя смесь линейных и разветвленных олигосахаридов. Второй этап реакции продолжается, пока продукты гидролиза не перестанут окрашиваться йодом; он протекает значительно медленнее и зависит от индивидуальных свойств фермента и его природы. Поэтому конечные продукты гидролиза а-амилазами могут быть различными. Первая стадия воздействия фермента на субстрат хотя и носит неупорядоченный характер, имеет для всех видов -амилаз схожий механизм.
Существует две гипотезы о механизме действия экзоамилаз на субстрат. Первая гипотеза предполагает, что, воздействуя на субстрат по одноцепочечному или «молниеобразному» механизму, экзоамилаза образует фермент-субстратный комплекс с захватом нередуцирующего конца цепи.
Дальнейшее продвижение фермента по этой цепи происходит до полного ее гидролиза. По второй гипотезе (- и глюкоамилаза действуют на субстрат путем механизма множественной атаки, т. е. фермент образует комплекс с молекулой субстрата, затем через несколько этапов этот комплекс распадается и фермент связывается с новой молекулой субстрата. Иными словами, при множественной атаке происходит нечто среднее между неупорядоченным механизмом и одноцепочечной, «молниеобразной» атакой. Для полного гидролиза по этому механизму одна
молекула субстрата должна образовывать много раз фермент-субстратные комплексы. При этом возможен гидролиз нескольких связей в одном каталитическом акте.
Механизм воздействия амилаз на субстрат может быть рассмотрен с нескольких позиций:
вид разрываемой связи (-1,4 или -1,6);
тип воздействия на субстрат (эндо- или экзо-);
влияние на скорость гидролиза степени полимеризации субстрата;
4) возможность гидролиза олигосахаридов;
способность фермента к множественной атаке субстрата.
Наличие признаков амилаз, отраженных в 3 и 4 позициях, при действии на линейные субстраты может свидетельствовать о существовании у этих ферментов подцентровой структуры. Вероятно, активный центр амилазы может состоять из нескольких подцентров, каждый из которых может вступать в контакты с глюкозным остатком. Энергия взаимодействия (А;), выраженная в единицах свободной энергии (кДж/моль), определяет подцентровое сродство фермента к субстрату. Это сродство индивидуально и может быть как положительным, так и отрицательным. Вероятность существования подцентровых структур амилаз помогает установить строение активного центра амилаз, дает более четкое объяснение субстратной специфичности, но не дает объяснений механизма гидролиза разветвленных субстратов.
-Амилаза. а-Амилаза (а-1,4-глюкан-4-глюканогидролаза, КФ 3.2.1.1.) является эндоамилазой, вызывающей гидролитическое расщепление -1,4-гли-козидных связей внутри высокополимеризованного субстрата. Фермент назван -амилазой потому, что он высвобождает глюкозу в -мутамерной форме.
а-Амилаза - водорастворимый белок, обладающий свойствами глобулина и имеющий М. м. 45 000-60 000. Своего рода исключением является -амилаза В. macerans, которая имеет М. м. 130 000. Есть указания, что некоторые термостабильные -амилазы имеют М. м. 14 000-15 000, но в их молекулах содержится в 2-3 раза больше атомов кальция.
Все а-амилазы относятся к металлоэнзимам, содержание в них Са колеблется от 1 до 30 г атом на 1 г моль фермента. Полное удаление кальция
приводит к инактивации фермента. Повторное введение кальция в среду может частично восстановить его активность. -Амилаза В. sub>tilis с помощью иона цинка способна образовывать димерную форму, чего лишены другие -амилазы. Все -амилазы устойчивы к воздействию протеаз. Они богаты тирозином и триптофаном. Глютаминовая и аспарагиновая кислоты составляют 25% массы белка. Наличие этих кислот в а-амилазе связывают с их оса-харивающей способностью. Так, разжижающие -амилазы не имеют сульфгидрильных групп, а осахаривающие содержат один остаток цистеина. Сравнительно мало или совсем отсутствуют в а-амилазах содержащие серу аминокислоты. Некоторые -амилазы грибного происхождения имеют углеводный фрагмент, в состав которого могут входить манноза, ксилоза, гексозоамин, но функции его не установлены.
В зависимости от вида микроорганизма свойства ос-амилаз могут сильно отличаться не только по механизму воздействия на субстрат и конечным продуктам, но и по оптимальным условиям для проявления максимальной активности.
Действуя на целое крахмальное зерно, -амилаза атакует его, разрыхляя поверхность и образуя каналы и бороздки, т.е. как бы раскалывает зерно на части. Клейстеризованный крахмал гидролизуется ею с образованием на окрашиваемые йодом продукты - в основном состоящие из низкомолекулярных декстринов. Процесс гидролиза крахмала многостадийный. В результате воздействия -амилазы на первых стадиях процесса в гидролизате накапливаются декстрины, затем появляются неокрашивающиеся йодом тетра- и тримальтоза, которые очень медленно гидролизуются -амилазой до ди- и моносахаридов.
Все -амилазы проявляют наименьшее сродство к гидролизу концевых связей в субстрате. Некоторые же а-амилазы, особенно грибного происхождения, на второй стадии процесса гидролизуют субстрат более глубоко с образованием небольшого количества мальтозы и глюкозы. Схему гидролиза под действием а-амилазы можно записать следующим образом:
а-Амилаза
Крахмал,----------------> а-Декстрины + Мальтоза + Глюкоза
гликоген (много) (мало) (мало)
-Амилаза. -Амилаза (-1,4-глюкан мальтогидролаза, КФ 3.2.1.2) - активный белок, обладающий свойствами альбумина. Каталитический центр фермента содержит сульфгидрильные и карбоксильные группы и имидозольный цикл остатков гистидина. -Амилаза -экзофермент концевого действия, проявляющий сродство к предпоследней -1,4-связи с нередуцирующего конца линейного участка амилозы и амилопектина.
В отличие от -амилазы -амилаза практически не гидролизует нативный крахмал, тогда как клейстеризованный крахмал гидролизуется ею с образованием мальтозы -конфигурации, поэтому данная амилаза по аналогии с -амилазой называется -амилазой. Если гидролизу подвергается амилоза, то гидролиз идет полностью до мальтозы. Незначительное количество декстринов может образовываться при гидролизе «аномальных» амилоз, так как гидролиз -амилазой идет только по линейной цепи до -1,6-связей. Если субстратом для -амилазы служит амилопектин, то гидролиз идет в значительно меньшей степени. -Амилаза отщепляет фрагмент с нередуцирующего конца участка от внешних линейных ветвей, имеющих по 20-26 глюкозных остатков, с образованием 10—12 молекул мальтозы. Гидролиз приостанавливается на предпоследней -1,4-связи, граничащей с -1,6-связью. В гидролизате накапливается 54-58% мальтозы, остальное составляют высокомолекулярные декстрины, содержащие значительное количество а-1,6-связей - так называемые -декстрины. Действие -амилазы на крахмал можно записать в виде следующей схемы:
-Амилаза
Крахмал,----------------> Мальтоза + р-Декстрин
гликоген (54-58%) (42-46%)
-Амилазы проявляют большую стабильность в отсутствие ионов Са2+. Молекулярная масса -амилазы растений достаточно высока, она составляет от 50 000 до 200 000. Фермент может состоять из одной или четырех
субъединиц до 50 000 каждая. Фермент содержит SH-группы и чувствителен к действию тяжелых металлов. Считается, что (-ами-лаза обладает высокой способностью к множественной атаке субстрата. Для амилозы средней молекулярной массы в одном присоединении фермента к субстрату возможно отщепление до четырех остатков мальтозы. При увеличении молекулярной массы субстрата возможно и большее количество мест атаки.
Глюкоамилаза. Глюкоамилаза (а-1,4-глюкан глюкогидролаза, КФ 3.2.1.3.) широко распространена в природе. Она синтезируется многими микроорганизмами и образуется в животных тканях, особенно в печени, почках, плаценте кишечника и т. д. Фермент в литературе известен под различными названиями: амилоглюкозидаза, -амилаза, лизосомальная -глюкозидаза, кислая мальтаза, матулаза и экзо-1,4--глюкозидаза. Глюкоамилаза катализирует последовательное отщепление концевых остатков -D-глюкозы с нередуцирующих концов субстрата. Это фермент с экзогенным механизмом воздействия на субстрат. Многие глюкоамилазы обладают способностью так же быстро, как и -1,4-связь, гидролизовать -1,6-глюкозидные связи. Но это происходит только в том случае, когда за -1,6-связью следует -1,4-связь, поэтому декстран ими не гидролизуется. Отличительной особенностью глюкоамилаз является способность в десятки раз быстрее гидролизовать высокополимеризованный субстрат, чем олиго-и дисахариды.
В литературе высказывается мнение, что механизм атаки субстрата глюкоамилазой может быть двух типов: либо одно-цепочечный, либо множественной атаки, и что активный центр имеет под-центровую структуру.
Почти все глюкоамилазы являются гликопротеидами, содержащими от 5 до 35% углеводов, которые состоят из олиго-, ди- и моносахаридов. Углеводный компонент может быть целостным фрагментом или же разбитым на индивидуальные соединения, которые прикрепляются к белку через треонин и серин. Например, у глюкоамилазы A. niger их 20. Большинство известных глюкоамилаз имеет оптимум рН при 4,5-5,2, реже - при 5,7-6,0, в основном для дрожжевых глюкоамилаз.
РН-стабильность микробных глюкоамилаз лежит в широком диапазоне -от 2,5 до 9. Термостабильность глюкоамилаз лежит в интервале от 30 до 45°С и редко повышается до 55-60°С. Глюкоамилазы различного происхождения заметно отличаются по молекулярной массе, которая, по данным различных авторов, имеет значения от 48 000 до 210 000. Следует заметить, что далеко не все микробные глюкоамилазы способы полностью гидролизовать крахмал до глюкозы. Еще в 60-х годах И.Фукумото предложил все микробные глюкоамилазы разделить на два типа:
полностью гидролизующие крахмал до глюкозы и
гидролизующие крахмал до глюкозы на 80-85%.
В то время предполагалось, что степень гидролиза зависит только от свойств глюкоамилаз и их происхождения. Позже было показано, что при росте культуры параллельно накапливаются и другие амилолитические ферменты, обладающие не только гидролитическим, но и трансферазным действием. Это гликозилтрансфераза и -амилаза. Даже в случае, если система открытая и продукт гидролиза (глюкоза) постоянно удаляется из системы, процесс может дойти до полного гидролиза крахмала до глюкозы. Если же система закрытая и концентрация субстрата велика, то при достижении определенной концентрации глюкозы в реакционной среде в результате переноса глюкозильных остатков на глюкозу, ди- и олигосахариды начинают накапливаться изомальтоза, паноза, нигероза, изомальтотриоза и другие сахара, которые имеют горький вкус. В результате процесс не может дойти до полного превращения крахмала в глюкозу и возникает ошибочное представление, что глюкоамилаза не полностью гидролизует крахмал. Сама же глюкоамилаза может проявлять небольшую трансферазную активность, но только при концентрации глюкозы свыше 60-70%. Поэтому ранее принятое деление глюкоамилаз на два типа следует считать необоснованным.
ТЕХНОЛОГИЧЕСКИЕ СХЕМЫ И РЕЖИМ ПОЛУЧЕНИЯ СПИРТА
Технология этилового спирта из крахмалистого сырья основана на ферментативном гидролизе зернового или картофельного крахмала и сбраживании образующихся сахаров дрожжевыми микроорганизмами, т.е. является биохимической технологией.
Процесс получения спирта из зерно-картофельного сырья включает следующие стадии: очистка и подготовка сырья, водно-тепловая обработка его, осахаривание разваренной массы и охлаждение сусла, приготовление засевной культуры дрожжей, сбраживание сусла, перегонка бражки и ректификация спирта.
Производство спирта из крахмалистого сырья осуществляется по периодической, полунепрерывной и непрерывной схемам. По периодической схеме с использованием разварников Генце получали этиловый спирт с конца XIX в. В настоящее время эта схема осталась только на некоторых спиртовых заводах малой мощности (до 800 дал). Полунепрерывная схема производства, основанная на использовании периодически действующих аппаратов, впервые была внедрена на заводах СССР в 1917—1950 гг. По этой схеме до сих пор работают около 15—20% спиртовых заводов страны.
ПРИНЦИПИАЛЬНАЯ ТЕХНОЛОГИЧЕСКАЯ СХЕМА ПОЛУЧЕНИЯ ЭТИЛОВОГО СПИРТА ИЗ КРАХМАЛИСТОГО СЫРЬЯ
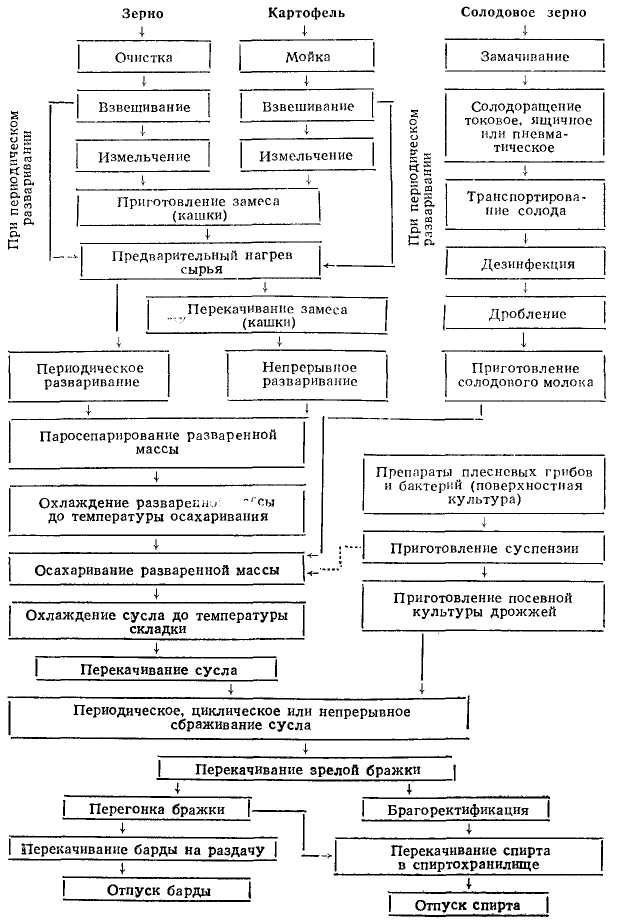
ОСАХАРИВАЮЩИЕ МАТЕРИАЛЫ
Крахмал, растворенный при разваривании зерна и картофеля, гидролизуют (осахаривают) амилолитическими ферментами зернового солода или культур микроорганизмов, преимущественно микроскопических (плесневых) грибов и бактерий.
Амилолитические ферменты содержатся во многих высших растениях, но наиболее богато ими пророщенное в определенных условиях зерно растений семейства мятликовых (злаков), называемое солодом. Способность солода осахаривать крахмал известна с древнейших времен, и с тех пор солод используется при получении спирта. Также давно известно свойство микроскопических грибов осахаривать крахмал. С их помощью восточные народы приготовляли различные охмеляющие напитки.
Культуры микроскопических грибов или ферментные пре параты применяют в спиртовой промышленности большинства зарубежных стран, причем в основном в виде концентрированных сиропообразных препаратов или сухого порошка, реже — в виде культуральной жидкости.
Культуры микроскопических грибов имеют ряд преимуществ по сравнению с солодом. Их выращивают на пшеничных отрубях или в составе питательной среды используют обычную кукурузную муку, тогда как для приготовления солода расходуется 14...20 % кондиционного (96 % проростаемости) зерна в расчете на массу крахмала сырья.
При солодоращении теряется 16...18 % крахмала, часть крахмала солода в процессе производства спирта остается неосахаренной и, следовательно, не сбраживается. Кроме того, с солодом вносятся в сусло посторонние микроорганизмы, вследствие чего в большей мере протекают и другие виды брожения, отрицательно отражающиеся на выходе спирта. В случае применения смеси солодов из различных злаков с целью полного осахаривания крахмала работа солодовен усложняется.
Культуры микроскопических грибов содержат комплекс амилолитических ферментов, отличающихся от ферментов солода и позволяющих глубже и полнее гидролизовать крахмал. В микроскопических грибах активнее целлюлозолитические ферменты, расщепляющие гемицеллюлозы до сахаров, часть которых сбраживается дрожжами, при этом повышается выход спирта.
С помощью культур микроскопических грибов можно увеличить концентрацию ферментов и таким образом сократить продолжительность осахаривания и последующего дображивания сусла в 2...3 раза.
Микроскопические грибы быстро размножаются, для выращивания поверхностной культуры достаточно около 1,5 сут, проращивание же зерна для получения солода длится 9... 10 сут. Глубинные культуры выращивают в стерильных условиях, что обеспечивает «чистоту» процесса спиртового брожения.
Действие солода и культур микроскопических грибов не ограничивается осахариванием крахмала, они еще способствуют накоплению в сусле достаточного количества органического азота для питания дрожжей и частичному растворению клеточных стенок эндосперма сырья. В осуществлении этих процессов, а также в выращивании солода и микроскопических грибов участвуют многочисленные ферменты, поэтому необходимо знание их химической природы, строения и механизма действия.
ПОЛУЧЕНИЕ СОЛОДА
Технология солода складывается из следующих основных процессов: замачивание и проращивание зерна; сушка солода и удаление ростков. На отечественных спиртовых заводах для осахаривания используют сырцовый — несушеный солод (неправильно называемый «зеленым»). Этот солод не может долго храниться, поэтому на каждом спиртовом заводе его готовят в количествах, необходимых для текущей работы.
ЗАМАЧИВАНИЕ ЗЕРНА
Основная цель замачивания — увлажнить зерно; дополнительная — отмыть от остатков пыли, удалить легкие зерновые и незерновые примеси и подавить микроорганизмы. Замачивание ведут с применением воздуха и воды, чередуя насыщение зерна водой и аэрацию.
Во время замачивания протекают физико-химические и биохимические процессы, приводящие к глубоким изменениям в зерне.
ФИЗИКО-ХИМИЧЕСКИЕ ПРОЦЕССЫ ПРИ ЗАМАЧИВАНИИ ЗЕРНА
Зерно прорастает нормально при влажности 40...46 %. При меньшей влажности ростки быстро увядают, накапливается мало ферментов, эндосперм плохо и неравномерно растворяется. Переувлажненное зерно долго не начинает прорастать, а затем быстро трогается в рост с большим выделением теплоты. Замачивание обычно заканчивается по достижении влажности 38...40 %, но обильно орошают зерно водой во время проращивания.
Вода поступает в зерно через плодовую и семенную оболочки, обладающие полупроницаемостью, поэтому в процессе замачивания главную роль играют ультрафильтрация и осмодиффузия. Цветочная пленка (мякинная оболочка) в начале замачивания непроницаема, и вода впитывается по тонким капиллярам — трахеидам зародышевой части, не покрытой этой оболочкой. Сорбируясь крахмалом, белками и другими высокополимерами и растворяя минеральные вещества, через полупроницаемые стенки клеток зародыша и эндосперма вода проникает внутрь зерна.
Со временем вследствие вымывания инкрустирующих веществ становится проницаемой и мякинная оболочка.
В нормально замоченном зерне ячменная влага распределена неравномерно. Наибольшая влажность (около 47 %) в основании — в зоне расположения зародыша; в самом зародыше влажность еще выше (68...75 %). В середине зерна влажность 38 %, в кончике 39 %.
Скорость замачивания зависит от структуры, размера зерна и температуры. Пленчатое, высокобелковое и крупное зерно обычно увлажняются медленнее, чем голое, низкобелковое и мелкое, хотя нередки исключения из этого правила. Зерно, выращенное в сухом жарком климате, впитывает влагу хуже зерна, полученного во влажном умеренном теплом климате. С повышением температуры скорость замачивания возрастает
Зерно большинства культур замачивают при 18...20 °С (смеси зерна и воды). При более высокой температуре необходимы частая смена воды, энергичное аэрирование, тщательное подавление микрофлоры, что усложняет управление процессом. Исключение составляет просо, которое насыщается влагой труднее, поэтому температуру замачивания его поддерживают в пределах 25...30 °С.
В воде с высокой жесткостью (14... 15 мгэкв/л) и большой щелочностью замачивание замедляется. Такое же действие оказывают хлориды; сульфаты, наоборот, ускоряют замачивание.
Во время замачивания зерно набухает, увеличиваясь в объеме, например, на 40...45 % (ячмень). Из твердого и хрупкого оно становится мягким и эластичным.
БИОХИМИЧЕСКИЕ ПРОЦЕССЫ ПРИ ЗАМАЧИВАНИИ ЗЕРНА
В результате увеличения влажности зерна при замачивании резко усиливается жизнедеятельность и в первую очередь дыхание зерна, сопровождающееся потребностью в кислороде. Вместе с тем запас кислорода в воде очень быстро уменьшается, напри-
мер при замачивании ячменя на 60...80 мин он исчезает, поэтому обеспечение зерна кислородом затруднено. Проникновению кислорода в зерно через зародыш (в начале замачивания) препятствует щиток, а через оболочки впоследствии — большое количество воды в тканях. Диффузия кислорода в воде примерно в 10 000 раз медленнее, чем в газе; кроме того, растворимость его в воде в 40 раз меньше, чем диоксида углерода. Недостаток кислорода в процессе замачивания подтверждается и значением дыхательного коэффициента, который выше единицы (около 1,07), а через 8 ч от начала замачивания равен 1,38, т. е. наблюдается уже анаэробное дыхание.
При кислородном голодании образуется этиловый спирт, вредно влияющий на жизнеспособность зародыша. В таких условиях частично нарушается структура тканей и зерно легко переувлажняется. Во время последующего проращивания требуются длительная перестройка типа дыхания, сжигание спирта и других метаболитов, на образование которых были затрачены углеводы. Отсюда следует, что с самого начала замачивания должны быть созданы условия для нормального дыхания зерна.
При замачивании зерна одновременно с усилением дыхания происходит глубокая перестройка всего ферментного комплекса.
ПРОРАЩИВАНИЕ ЗЕРНА
Цель солодоращения — накопление ферментов, растворение межклеточных пластинок и стенок клеток эндосперма, что необходимо для снабжения развивающегося зародыша питательными веществами и перехода ферментов в сусло.
Зерно проращивают в таких условиях, чтобы расход крахмала
на дыхание и образование новых вегетативных органов был минимальным, при возможно меньшем обсеменении микроорганизмами, особенно кислотообразующими.
МОРФОЛОГИЧЕСКИЕ И ЦИТОЛИТИЧЕСКИЕ ИЗМЕНЕНИЯ ЗЕРНА
При проращивании в зерне происходят процессы распада и синтеза. В эндосперме гидролизуются резервные вещества — крахмал, белки, а также пектиновые вещества, гемицеллюлозы, целлюлоза; образующиеся растворимые продукты поступают через щиток в зародыш. В результате процессов синтеза из зародыша вырастают стебелек и корешки.
БИОХИМИЧЕСКИЕ ИЗМЕНЕНИЯ ЗЕРНА
Ферменты синтезируются в вегетативных частях растения — листьях и стеблях — и уже из них мигрируют в созревающее зерно. По исследованиям А. Н. Баха и А. И. Опарина, активность ферментов в зерне вначале увеличивается, в период молочной спелости достигает максимума, в период восковой спелости снижается и, наконец, во вполне созревшем зерне еле проявляется. При проращивании зерна активность ферментов вновь увеличивается, достигая даже значительно большей величины, чем в период молочной спелости.
Понижение активности амилаз при созревании зерна объясняется связыванием их с белками. В таком, зимогенном, состоянии амилазы нерастворимы и потому неактивны. Активность их восстанавливается после воздействия протеаз, освобождающих амилазы из зимогена.
В созревающем зерне ячменя, ржи, пшеницы и овса присутствуют - и -амилазы, но в зрелом зерне обнаруживаются лишь следы -амилазы. Заметное содержание свободной -амилазы найдено в зерне ржи и овса. В пророщенном зерне всех этих культур содержание р-амилазы не превышает таковое при действии папаина на исходное зерно, т. е. р-амилаза накапливается в солоде в результате освобождения из зимогена. Полагают, что ос-амилаза накапливается таким же путем и частично синтезируется.
Динамика изменения ферментативной активности ячменя при его проращивании (по Г. И. Фертману и А. Н. Лазаревой) показана на рис. 3. Видно, что величины АС и ДС достигают максимального значения на 10-е сутки, по другим данным, у ржи — на 7...8-е сутки. Декстринолитическая способность возрастает v овса на 10... 12-е, у
проса — на 5...6-е сутки. Исходя из этого, устанавливают и продолжительность солодоращения.
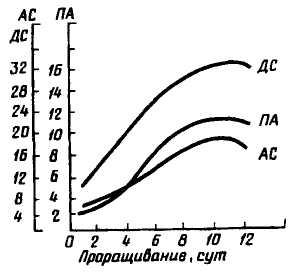
Рис. 3. График изменения ферментативной активности в прорастающем ячмене
В солоде амилолитические ферменты распределены неравномерно: в ячменном приблизительно 70 % амилаз локализовано в нижней части эндосперма, прилегающей к щитку, в верхней части их около 25, в щитке 4, в стебельке и корешках 1 %.
Цитолитические ферменты в процессе проращивания зерна также активируются, и активность их возрастает до определенного времени, у ячменя — до 5...7 сут. Наибольшей активности цитазы соответствует переход твердого состояния мучнистого тела в рыхлое, когда эндосперм легко может быть растерт между пальцами.
Главная роль в растворении клеточных стенок принадлежит гемицеллюлазам и пектиназам. Основной составной частью ге-мицеллюлоз ячменя является -глюкан, гидролиз которого катализируется эндо--глюканазой. Меньшая роль принадлежит ара-бинозидазе и ксиланазе.
В зерне злаков из протеиназ содержится фермент типа папаина, из пептидаз — аминопептидазы, карбоксипептидазы и дипептидазы.
В результате действия протеиназ из зимогена освобождаются амилазы, под действием пептидаз накапливаются аминокислоты, главным образом аспарагиновая и глутаминовая. Активность протеаз возрастает в процессе солодоращения соответственно увеличению активности амилаз
Протеиназы ячменя, как и папаин, в зависимости от природы гидролизуемого белка могут проявлять свою активность при рН 3,8; 6,3 и 8,6, в соответствии с чем их подразделяют на кислые, нейтральные и щелочные. При солодоращении наибольшую каталитическую активность проявляют кислые протеиназы, активаторами которых служат сульфгидрильные соединения, содержащие группу —Н, цистин и восстановленный глютатион. В первые сутки проращивания зерна количество глютатиона значительно увеличивается, причем в зародыше более энергично, чем в эндосперме. В последующие сутки накопление глютатиона в зародыше происходит медленнее, но все время в зародыше его больше, чем в эндосперме.
Активность кислой протеиназы в продолжение солодоращения возрастает приблизительно в 40 раз. Пептидазная активность проявляется также сильно, но позже протеиназной.
К концу проращивания в зерне накапливаются довольно активные липаза и фосфатаза (фитаза, нуклеотидаза). Активность фосфатазы тем выше, чем ниже температура солодоращения.
По данным А. Н. Баха и А. И. Опарина, в зерне активность ферментов дыхания — оксидазы, пероксидазы и каталазы — выше активности гидролитических ферментов. В результате проращивания повышается активность обеих групп ферментов, но соотношение их активности резко изменяется в обратную сторону и тем сильнее, чем ниже температура. Поэтому в процессе солодоращения накапливается значительное количество гидролитических ферментов при сравнительно небольших тратах крахмала на дыхание.
ИЗМЕНЕНИЕ ХИМИЧЕСКОГО СОСТАВА ЗЕРНА
Несмотря на то что солодоращение протекает при сравнительно низких температурах, сильно отличающихся от оптимальных для действия ферментов, за время проращивания зерна происходят существенные изменения его химического состава.
Наибольшие преобразования претерпевает крахмал — основной резервный углевод зерна. Приблизительно 20 % от всего его количества гидролизуется: из них 8...9 % расходуется на дыхание, 3...4 % на построение стебля и корней и 8... 10 % остается в виде сахара, придающего солоду сладкий вкус.
Свободные сахара состоят главным образом из сахарозы, ин-вертного сахара и мальтозы. При температуре проращивания 15...16 °С образуются преимущественно сахароза и продукты ее гидролиза, при температуре 20...23 °С — мальтоза.
В нерастворенных гранулах крахмала при рассмотрении под микроскопом хорошо видны канальцы и повреждения поверхности, что является результатом действия амилаз. Размер гранул несколько уменьшается, повышается содержание амилозы, внешние цепи амилопектина укорачиваются. Температура клейс-теризации возрастает приблизительно на 4 С, а вязкость клейстера, наоборот, понижается.
Количество целлюлозы в зерне и солоде почти одинаково. Это объясняется тем, что наряду с растворением клеточных стенок эндосперма примерно такое же количество клетчатки образуется в новых вегетативных органах. Количество пентозанов в солоде на 2...3 % больше, чем в исходном зерне. Большая часть пентозанов растворима в воде и состоит не только из высокомолекулярных продуктов гидролиза, но и из моносахаридов (ксилозы, арабинозы).
Белковые вещества также претерпевают значительные изменения. При существующих способах производства спирта азотистое питание для дрожжей накапливается в основном процессе солодоращения. Некоторое количество растворимого азота образуется и при осахаривании разваренной массы, но оно относительно невелико.
Изменения состава белковых веществ ячменя и солода характеризуют данные, приведенные в табл. 1. Наиболее сильному гидролизу подвергается гордеин, несколько меньшему — глютелин; количество альбумина и глобулина почти не изменяется. Одновременно в 4...5 раз увеличивается содержание аминокислот. Об их составе некоторое представление может дать анализ ячменя и пивоваренного солода.
Содержание общего азота в зерне и солоде, %
|
Белковые вещества |
Ячмень |
Солод |
|
Гордеин Глютелин Глобулин Альбумин Протеазы Аминокислоты |
36 30 10 12 5 7 |
17 21 11 10 9 32 |
Общее содержание азота на протяжении всего периода соло-доращения остается практически таким же, содержание аминного азота резко возрастает на 6...8-е сутки, а затем темпы роста замедляются. Белки исходного ячменя гидролизуются примерно на 55 %, из которых около 23 % сосредоточивается в проростках в виде качественно иных белков.
В процессе солодоращения содержание жиров уменьшается на 10...30 %. От фосфорорганических соединений отщепляются фосфаты. Образуются продукты неполного окисления углеводов — лимонная, щавелевая, молочная и другие органические кислоты, которые вместе с аминокислотами повышают общую кислотность, например с 1,5...2,5 мл 1 н. раствора NaOH на 100 г ячменя до 4,4...7,5 мл на 100 г солода. Однако значения рН в вытяжках из ячменя и солода вследствие буферных свойств мало различаются, но буферная емкость вытяжки из солода на 20...40 % больше.
При солодоращении освобождается инозит и возрастает содержание других витаминов — тиамина и рибофлавина, имеющих важное значение для жизнедеятельности и бродильной энергии дрожжей. Образуются эфиры и другие соединения, придающие солоду специфический запах: свежих огурцов — ячменному, стручков акации, медовый или яблочный — просяному и т. д.
ИНТЕНСИФИКАЦИЯ ПРОЦЕССА СОЛОДОРАЩЕНИЯ
Для интенсификации солодоращения с сокращением срока проращивания и повышением ферментативной активности в спиртовой промышленности применяют биостимуляторы — гиб-берелловую кислоту и комплексный ферментный препарат МЭК-1.
Гибберелловую кислоту используют в виде водного раствора при поливе зеленого солода в процессе выращивания, а также при замачивании зерна перед солодоращением. При поливе расход гибберелловой кислоты (в пересчете на чистое ее количество) — 600 мг/т (ячмень, рожь и овес) и 400 мг/т (просо). Если
эту кислоту добавляют на стадии замачивания зерна, то расход ее увеличивается и составляет 660...800 мг/т.
При использовании гибберелловой кислоты продолжительность выращивания ячменного и овсяного солода 8...9 сут. Ферментативная активность зеленого солода, обработанного гибберелловой кислотой, повышается не менее чем на 15 % по сравнению с необработанным солодом.
Для интенсификации солодоращения нормального зерна, а также при получении солода из нестандартного зерна в целях повышения прорастаемости и увеличения ферментативной активности в качестве биостимулятора кроме гибберелловой кислоты применяют комплексный ферментный препарат МЭК-1. Под его действием интенсифицируются процессы растворения мучнистой части зерна, образования амилолитических ферментов при проращивании, в результате чего продолжительность солодоращения сокращается с 10...12 до 6...7 сут.
В комплексный ферментный препарат МЭК-1 входят Амилосубтилин Г1Ох и Амилоразин П1Ох.
Препарат МЭК-1 (мультиэнзимная композиция, тип I) представляет собой порошок светло-бежевого цвета влажностью 12...13 %. Амилолитическая активность препарата (АС) 2650 ед/г, протеолитическая (ПС) — 30 ед/г. Расход МЭК-1 составляет 50 г стандартного препарата на 1 т солодового зерна при замачивании. Препарат применяют в виде рабочего водного раствора (50... 100 г препарата растворяют в 10 л воды). Рабочий раствор используют не только для замачивания, но и для полива при проращивании из расчета 200 л на I т солодового ячменя.
Повышение ферментативной активности солода с помощью биостимуляторов при уменьшении общей продолжительности проращивания способствует сокращению удельных потерь крахмала.
ПРИГОТОВЛЕНИЕ СОЛОДОВОГО МОЛОКА
Готовый солод, направляемый в производство, учитывают по массе или объему. Лучший способ его подачи — гидравлический: он удобен, солод хорошо промывается водой и очищается от микроорганизмов. Недостатки гидроподачи — большой расход воды (около 1000 % массы солода) и потеря части отломанных корешков со сточной водой.
Солод вместе с водой насосом перекачивается на барабанное разделительное сито, транспортная вода проходит через уловитель корешков, а солод направляется в дробилку и далее в бак для приготовления солодового молока. Чтобы улучшить доступ ферментов к разваренной массе сырья, солод измельчают на молотковых, дисковых или вальцовых дробилках. Более тщательное измельчение достигается на дисковых дробилках.
Рабочий орган дисковой дробилки — два вертикальных стальных диска, из которых один установлен неподвижно, другой вращается с частотой 750 об/мин. Зубья дисков поставлены так, что зуб одного диска входит в выемку между зубьями второго, образуя бороздки, по которым солод проходит от центра к периферии дисков. Расстояние между зубьями дисков регулируется с помощью специального приспособления, расположенного между поддерживающими вал подшипниками и состоящего из коробки и двух регулирующих маховичков. В некоторых конструкциях дисковых дробилок для лучшего измельчения солода предусмотрена многократная циркуляция его между дисками дробилки и баком. В таких циркуляционных дробилках корпус улиткообразный, а зубья привариваются тангенциально.
Для дезинфекции солода при гидротранспортировании в транспортную воду добавляют хлорную известь или хлорамин ХБ технический. Раствор хлорной извести готовят из расчета 400 мг активного хлора на 1 л воды. Например, хлорной извести марки Б II сорта, содержащей не менее 32 % активного хлора, берут 125 г на 100 л воды. В воде с указанной концентрацией активного хлора солод выдерживают 25...30 мин.
При подаче ленточным транспортером или ковшовым элеватором солод обрабатывают в течение 25...30 мин в сборнике с мешалкой водным раствором хлорной извести, который затек спускают в канализацию. При отсутствии хлорной извести используют формалин из расчета 2,8 л 30 %-ного формальдегида на 1 м3 воды.
Тонкоизмельченный солод дополнительно обрабатывают формалином в сборнике с мешалкой. В этой емкости солод смешивают с водой в соотношении 1:(2...2,5), после чего в солодовое молоко (на 1 м3) приливают 25...28 мл 37 %-ного раствора формалина с таким расчетом, чтобы концентрация его в сусле составляла 0,025 %. Раствор выдерживают в течение 25...30 мин, разбавляют чистой водой и перекачивают в расходный сборник. Общий расход воды 4...5 л на 1 кг солода.
Для осахаривания зерно-картофельного крахмала применяют смесь ячменного (50 %), просяного (25 %) и овсяного (25 %) солодов, причем общее содержание просяного и овсяного соло-дов должно быть не менее 30 %. Можно использовать смесь из двух солодов: ячменного и овсяного или просяного. Ячменный солод можно заменить ржаным полностью или частично, а просяной — солодом из чумизы. Запрещается применять солод из одной культуры, например ячменя, при производстве спирта из зерна той же культуры.
При осахаривании разваренной массы смесью солодов, состоящей, например, из 70 % ячменного и 30 % просяного солодов, с нерастворенным крахмалом в бражке теряется до 20...40 % крахмала солода, так как крахмала ячменного солода растворяется
60...64 %, просяного — 24 %. При проращивании на солод нестандартного зерна, имеющего пониженную прорастаемость, эти потери могут быть значительными только за счет непроросших зерен.
ПОЛУЧЕНИЕ МИКРОБНЫХ ФЕРМЕНТНЫХ ПРЕПАРАТОВ
Микроорганизмы способны синтезировать разнообразные ферменты. В зависимости от состава питательной среды и условий культивирования они легко переключаются с синтеза одного фермента на синтез другого. У микроорганизмов сравнительно короткий цикл развития (10... 100 ч), вследствие чего можно получать сотни «урожаев» в год.
Продуцентами ферментов могут быть бактерии, грибы, дрожжи и актиномицеты. Для промышленного получения ферментных препаратов используют как природные штаммы микроорганизмов, выделенные из естественных сред, так и мутантные,
отселекционированные в результате воздействия на природные штаммы физических и химических мутагенов.
Микроорганизмы синтезируют одновременно комплекс ферментов, но некоторые из них, особенно мутантные штаммы, продуцируют один фермент в значительных количествах. Для лучшего использования крахмалсодержащего сырья в спиртовом производстве осахаривающие материалы должны содержать не только амилолитические ферменты, но и ферменты, гидролизующие другие углеводы сырья — целлюлозу и гемицеллюлозы. Для обеспечения дрожжей азотистым питанием имеют значение и протеолитические ферменты.
Несмотря на то, что для успешного осахаривания нужен комплекс ферментов, отбор микроорганизмов-продуцентов до сих пор проводился главным образом по высокой активности амилолитических ферментов — -амилазы и глюкоамилазы.
МИКРООРГАНИЗМЫ - ПРОДУЦЕНТЫ ФЕРМЕНТОВ
Наиболее часто в качестве продуцентов амилолитических ферментов в спиртовом производстве используют микроскопические грибы, реже — дрожжеподобные организмы и споровые бактерии.
МИКРОСКОПИЧЕСКИЕ ГРИБЫ
Для получения амилаз широко применяют микроскопические грибы рода Aspergillus, видов: niger, oryzae, usamii, awamori, bata-tae, рода Rhizopus, видов: delemar, tonkinensis, niveus, japonicum и др., а также отдельные штаммы Neurospora grassa и Mucor.
Микроскопические грибы очень широко распространены в природе; основное место их обитания — почва. Несмотря на наличие многих родов и видов микроскопических грибов, все они характеризуются нитевидным строением тела и специфическим строением плодоносящих органов. Тело гриба состоит из длинных переплетенных нитей сероватого или белого цвета, называемых гифами. Они распространяются по поверхности питательного субстрата, образуя мицелий, и частично врастают в него. Некоторые гифы, поднимающиеся над поверхностью в виде легкого пушка, имеют более сложное строение и представляют собой органы плодоношения, называемые конидие- или спорангиеносцами. У мукоровых грибов на конце спорангиеносца находится шаровидное вздутие, окруженное оболочкой, внутри которого образуются споры. У аспергиллов конец конидиеносца имеет булавовидное утолщение, от которого отходят удлиненные клетки, называемые стеригмами; от стеригм отшнуровываются более мелкие круглые клетки — конидии.
Отделившиеся конидии или споры, попадая в благоприятные условия, начинают прорастать, затем гифы ветвятся, образуя ми-
целий; при истощении питательных веществ в среде гриб переходит в стадию споро- или конидиеобразования. Споры и конидии микроскопических грибов содержат пигменты, что и придает зрелым культурам характерную окраску.
Аспергиллы — типичные аэрофилы, поэтому они могут развиваться только на поверхности твердой или жидкой среды или в жидкой, достаточно аэрируемой среде. Оптимальная температура для большинства аспергиллов 25...30 "С, для некоторых — до 35 °С. Большинство грибов при поверхностном культивировании могут переносить кратковременное повышение температуры до 40 °С и даже 45 °С без заметной потери активности ферментов. Оптимальная влажность среды для них около 65 %.
Для питания аспергиллов необходимы углеводы, азотистые и минеральные вещества. В качестве источника углевода, кроме моносахаридов, многих олиго- и полисахаридов, могут служить спирты и органические кислоты, однако для накопления амилазы в среде обязательно должны присутствовать крахмал, декстрины или мальтоза. В средах, содержащих другие сахара, в том числе глюкозу, грибы не образуют амилазы. Источником азота могут быть белки и их гидролизаты, аммонийные соли и нитраты.
Среда должна содержать соединения, в состав которых входят сера, фосфор, калий, магний и микроэлементы. Большинство микроскопических грибов усваивают серу из сульфатов, а фосфор — из фосфатов. Аспергиллы не нуждаются в готовых вита-
минах и факторах роста, так как способны сами синтезировать их из более простых соединений, имеющихся в среде. Препараты ферментов из микроскопических грибов включают, как правило, широкий набор ферментов, поэтому могут полностью заменять зерновой солод.
На спиртовых заводах стали широко применять высокоактивный по глюкоамилазе штамм Asp. awamori 466 и ВУД-Т2, выращиваемые на концентрированном кукурузном сусле (18 % сухих веществ). Готовая культура имеет активность до 250 ед. ГлА на 1 мл, других ферментов образует мало.
ДРОЖЖЕПОДОБНЫЕ ОРГАНИЗМЫ
Амилолитические ферменты синтезируют также некоторые дрожжи и дрожжеподобные грибы родов Saccharomyces, Candida, Endomycopsis и Endomyces.
В спиртовом производстве нашли применение End. bispora и End. species 20-9, выращиваемые глубинным способом и продуцирующие главным образом активную глюкоамилазу; ос-амилаз-ная активность проявляется слабо. Высокоактивный End. bispora имеет разветвленный мицелий, образует бластоспоры; гифы — септированные, зернистые; на твердых агаризованных средах образуют колонии с воздушным серовато-белым мицелием, на жидких питательных средах — гифы и некоторое количество бластоспор.
Дрожжеподобные грибы в спиртовом производстве самостоятельно не применяют, так как они не содержат других ферментов, необходимых для нормального осахаривания сусла из крах-малсодержащего сырья. Обычно их используют в смеси с ферментными препаратами из микроскопических грибов или бактерий.
БАКТЕРИИ
Активные амилазы способны синтезировать многие бактерии: Вас. sub>tilis, Вас. diastaticus, Вас. mesentericus, Вас. macerans и Вас. polymycus и др.
Бактерии — продуценты амилолитических ферментов представляют собой палочки длиной 1,2...1,3 мкм и диаметром 0,6...0,8 мкм. Палочки соединяются по две, три, иногда образуют цепочки. Цикл развития бактерий короче, чем микроскопических и дрожжеподобных грибов. Например, культуру Вас. diastaticus выращивают в глубинных условиях при температуре 60 °С в течение 10... 12 ч.
Бактерию Вас. sub>tilis-82, применяемую в настоящее время на спиртовых заводах как продуцент -амилазы в смеси с препара-
тами глюкоамилазы, выращивают в течение 48...60 ч при температуре 30...35 'С.
Особенность бактерий — их способность образовывать высокоактивную термостойкую -амилазу, необходимую для разжижения и декстринизации крахмального клейстера на стадии под-варивания замесов и осахаривания сусла.
ПРОИЗВОДСТВЕННЫЕ СПОСОБЫ КУЛЬТИВИРОВАНИЯ МИКРООРГАНИЗМОВ - ПРОДУЦЕНТОВ ФЕРМЕНТОВ
Существует два способа культивирования микроорганизмов — продуцентов ферментов: поверхностный и глубинный.
Первый способ, применяемый для культивирования микроскопических грибов, характеризуется развитием мицелия на поверхности твердого или жидкого субстрата. На жидком субстрате образуется пленка мицелия, продуцирующего не только амилолитические ферменты, но и органические кислоты, инактиви-рующие их, поэтому используют твердые субстраты с развитой поверхностью — пшеничные отруби, дробину барды, картофельную мезгу и др.
Максимальная активность ферментов достигается при культивировании грибов на пшеничных отрубях. Дробина барды бедна питательными веществами, и активность ферментов в культурах грибов, выращенных на ней, в 4...5 раз ниже, чем на отрубях. Зрелая культура грибов вследствие обволакивания частиц отрубей мицелием имеет вид плотной войлокообразной массы.
При поверхностном культивировании пшеничные отруби должны быть увлажнены и простерилизованы. В стерильных условиях готовят посевную культуру, но выращивают грибы в нестерильных условиях в кюветах, устанавливаемых в негерметичных растильных камерах. Теплоту, выделяющуюся в процессе роста грибов, удаляют продуванием через растильную камеру стерильного кондиционированного воздуха.
Поверхностный способ выращивания микроскопических грибов имеет ряд преимуществ. Так как во время роста гриба отруби не перемешиваются, посторонние микроорганизмы не распространяются по всей их массе и вызывают лишь незначительное местное инфицирование, которое, как правило, не влияет на активность ферментов. Это, однако, не исключает необходимости тщательной стерилизации воздуха, среды и оборудования. Культуру на отрубях высушивают до содержания влаги 10... 11 %. В таком виде она может храниться продолжительное время без значительной потери активности. Это позволяет организовать централизованное снабжение спиртовых заводов сухой культурой микроскопических грибов, что является одним из преимуществ поверхностного способа выращивания.
Недостаток поверхностного способа — необходимость устанавливать множество кювет, работу с которыми трудно механизировать. Себестоимость культуры гриба-продуцента высока,
причем в основном из-за затраты большого количества ручного труда. Механизация процесса выращивания возможна путем создания непрерывнодействующих установок или бескюветных аппаратов с вертикальным толстым слоем питательной среды и интенсивным продуванием воздуха через этот слой.
Глубинную культуру микроорганизмов выращивают на жидкой питательной среде при энергичной аэрации в герметически закрытых аппаратах и в стерильных условиях. Процесс полностью механизирован. Стерильность глубинной культуры микроорганизма — продуцента ферментов положительно отражается на результатах сбраживания сусла дрожжами.
ПОВЕРХНОСТНОЕ КУЛЬТИВИРОВАНИЕ МИКРОСКОПИЧЕСКИХ
ГРИБОВ
Питательные среды. При поверхностном культивировании твердой средой обычно служат пшеничные отруби, содержащие в достаточных количествах все вещества, необходимые для питания грибов. Лучшие результаты достигаются при содержании не менее 16 % крахмала в отрубях или в другой питательной среде.
Получение посевного материала. Для засева производственной питательной среды может быть использован посевной материал трех видов: споры, мицелий и спороносящая культура, выращенная на твердой питательной среде.
Споровый материал готовят поверхностным культивированием гриба на твердой или жидкой питательной среде в специальных аппаратах с кюветами. В первом случае гриб выращивают в пробирке на косом агаре до обильного образования спор, затем спорами засевают колбы со стерильными пшеничными отрубями, наконец, спороносящую культуру передают в специальный аппарат. По окончании спорообразования в этом аппарате отбирают споры посредством специального вибросепаратора при тщательной аспирации. Споры фасуют в полиэтиленовые мешочки, стеклянные или дюралюминиевые банки, в которых они могут сохраняться при температуре от 8 до 24 °С около 1,5 лет.
Во втором случае из спор, собранных в пробирке с косым агаром, готовят водную суспензию и ею засевают жидкую питательную среду, которую направляют в растильный аппарат. Из-под выросшей спороносящей пленки удаляют жидкую среду, пленку подсушивают, снимают с кювет, измельчают и упаковывают. Готовый споровый материал не содержит примеси среды и обладает высокой всхожестью (90...95 %), которая сохраняется свыше 3 лет.
Споры (конидии), полученные любым методом, обладают водоотталкивающей способностью и почти не смачиваются водой. Это свойство приводит к неравномерности засева среды и, следовательно, к неодинаковой скорости роста культуры. Смачивае-
мость спор можно улучшить, если добавить в их водную суспензию 25...50 мг поверхностно-активного вещества, например ал-килбензолсульфата на 1 г спорового материала. При этом всхожесть спор остается без изменения.
Для сокращения длительности культивирования микроскопических грибов в производственных условиях можно применять предварительное проращивание спор в течение 7 ч в жидкой питательной среде до появления ростовых трубочек. При этом продолжительность лаг-фазы сокращается на 8...9 ч и увеличивается оборачиваемость растильных камер.
При использовании спорового материала упрощается технологический процесс, появляется возможность более полно механизировать его, а также сократить площадь цеха чистой культуры. Централизованное производство спорового материала для группы предприятий, работающих с данным штаммом гриба, выгодно создать в одном специализированном цехе чистой культуры. Положительный опыт подобной организации имеется в производстве лимонной кислоты биотехнологическим способом.
Исследователями б. ВНИИПрБ было предложено засевать производственную питательную среду мицелием, полученным глубинным культивированием гриба или бактерий в колбах на качалке. При высокой производительности предприятия целесообразно выращивать посевной мицелий в небольших ферментерах, как это было предложено А. П. Левчиком и осуществлялось на Мичуринском экспериментальном ферментно-спиртовом заводе.
На спиртовых заводах получило распространение приготовление посевного материала в виде поверхностной культуры гриба на пшеничных отрубях, которое складывается из следующих двух операций:
выращивание спороносной культуры гриба в пробирках на сусло-агаре;
засев спорами пшеничных отрубей в колбах и выращивание культуры гриба.
Культуру получают из лаборатории чистых культур ВНИИПрБ. Исходная — музейная чистая культура хранится в холодильнике при температуре от 2 до 4 °С. Один раз в месяц пересевают споры музейной культуры из пробирки в пробирку с сусло-агаром с соблюдением правил проведения микробиологических работ. При этом первая засеянная пробирка является музейной, а остальные две-три пробирки используют для дальнейшего размножения. Засеянные пробирки помещают в термостат при температуре 30 С и выдерживают в нем в течение 4...6 сут.
При нормальном росте поверхность сусло-агара равномерно покрывается спороносящим мицелием. Культура гриба с плохим спороношением, медленно растущим мицелием, пушком вторичного роста в производство не допускается. Пробирки с вырос-
шей культурой вынимают из термостата и хранят в холодильнике. Перед посевом в колбы проверяют чистоту культуры высевом в чашки Петри и микроскопированием.
Чистая культура на сусло-агаре служит исходным материалом для размножения грибов на пшеничных отрубях в колбах. Отруби смешивают с водопроводной водой в соотношении 1:0,7 и переносят в несколько колб по 10...20 г в каждую. Колбы закрывают ватными пробками и стерилизуют в автоклаве при избыточном давлении 0,15 МПа в течение 1 ч. После охлаждения до комнатной температуры в колбы вносят суспензию спор гриба, содержащую 150...200 тыс. спор в 1 мл. Для этого в пробирку с культурой гриба на сусло-агаре прибавляют 10 мл стерильной водопроводной воды и при помощи стерильной пипетки над огнем соскабливают конидии гриба. Полученную суспензию используют для посева (2...2,5 % массы засеваемой среды или 0,4...0,5 мл на 10 г отрубей).
Выращивание ведут в термостате при температуре 30 °С в течение 4...5 сут. Одну из колб предназначают для дальнейшего размножения чистой культуры, остальные колбы хранят в холодильнике до последующего цикла. В колбу, используемую для размножения, вливают 50...60 мл стерильной водопроводной воды и над пламенем стерильной пипеткой тщательно перемешивают.
Расход чистой культуры при пересеве из колбы в колбу или банку составляет 5... 10 % массы засеваемой среды.
Размножение посевного материала. Пшеничные отруби подготавливают так же, как и при получении чистой культуры. Отруби, стерилизованные в бюксах под избыточным давлением 0,15 МПа в течение 1,5 ч, переносят на специальный стол, предварительно вымытый и продезинфицированный. Охлаждение отрубей до температуры 40...42 "С проводят перемешиванием с соблюдением правил микробиологической чистоты. Затем их засевают спорами из колбы с чистой культурой (расход культуры 0,5... 1 % массы отрубей). Суспензию спор готовят из расчета 1 часть культуры на 5 частей воды. Засеянные отруби распределяют по кюветам, предварительно простерилизованным при 120 °С в течение 2 ч, слоем толщиной 2...2,5 см. Кюветы сверху закрывают крышкой, под которую подкладывают стерильную бумагу; на отверстие по центру крышки накладывают стерилизованную ватную подушку, после чего кюветы помещают в термостат на 18...24 ч при температуре 30...32 "С. В дальнейшем кюветы устанавливают на стеллажи, расположенные в самой растильной камере, и ведут выращивание при 24...28 °С в течение 3...4 сут. Выросшую культуру затем помещают в комнату подсушки и в воздушно-сухом состоянии передают на хранение в кладовую, температура в которой не должна превышать +8 °С, но не ниже 0°С.
Посевной материал, приготовленный описанным способом, представляет собой спороносную культуру гриба, содержащую не менее 0,7 млрд спор в 1 г, из которых 86...90 % способны к прорастанию. Посевной материал не должен содержать посторонней микрофлоры, подвергаться замораживанию, оттаиванию и увлажнению.
Производственное культивирование. Технологическая схема получения культуры микроскопических грибов в растильных камерах на кюветах приведена на рис, 4. Этот способ связан с большими затратами ручного труда, однако пока применяется на заводах.
Пшеничные отруби ковшовым элеватором / подают на автоматические весы 2, отсюда они поступают в загрузочный бункер 3, а из него — в стерилизатор 5. В этом аппарате отруби увлажняются водой, подаваемой через форсунки из бака 4 для стерильной воды. На 1 кг отрубей добавляют 0,2 л воды и 7...8 мл соляной кислоты относительной плотностью 1,19 или 2,7...3,0 мл серной кислоты плотностью 1,84. Отруби стерилизуют острым паром при температуре 103... 105 "С (давление 0,07 МПа) в течение 1...1,5 ч, периодически включая мешалку. По окончании стерилизации влажность отрубей доводят до 58...60 %, охлаждают их до 40...42 °С и затем вносят культуру гриба, полученную в отделении чистых культур.
Расход посевного подсушенного материала составляет 0,5...0,6 % массы отрубей. Посевной материал вводят в виде водной суспензии, приготовляемой из культуры гриба в 5... 10-кратном количестве кипяченой воды.
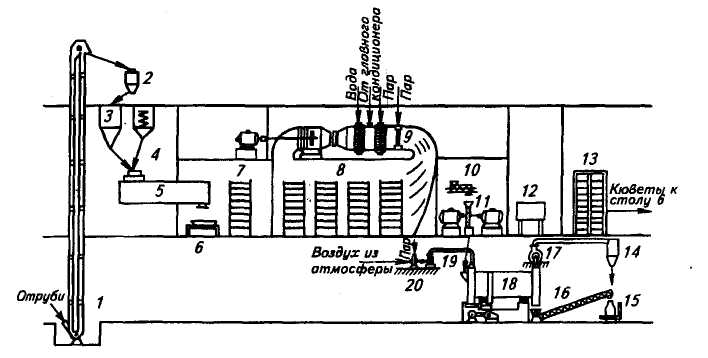
Рис. 4. Технологическая схема получения культуры микроскопических грибов в
камерах на кюветах
Засеянную среду тщательно перемешивают в течение 15...20 мин, выгружают на раскладочный стол 6 и распределяют по кюветам. Наполненные кюветы ставят в этажерки 7, перевозят их на тележках в растильную камеру 8, в которой поддерживают температуру 30...32 °С в течение 12... 16 ч (до начала интенсивного роста). В дальнейшем из кондиционера 9 в камеру подают вентилятором кондиционированный воздух температурой 26...28 °С и относительной влажностью 96...100 %. К концу выращивания культуры температуру снижают до 24...26 °С. Общая продолжительность выращивания 36...42 ч. В тех случаях, когда использование готовой культуры для осахаривания разваренной массы задерживается более чем на 2 ч, ее высушивают до влажности 18...20 %.
При необходимости транспортировки на дальние расстояния культуру гриба дробят на шнеке-дробилке 10 и дезинтеграторе 11 до получения частиц размером не более 10 мм и высушивают в сушилке 18 теплым воздухом до содержания влаги не более 10... 12 %. Воздух нагревают в калорифере 20 и подают в сушилку вентилятором 19. Отработавший воздух удаляют вентилятором 17 и выбрасывают в атмосферу через циклон 14. Высушенную культуру шнеком 16 подают на весы 75 для фасования. Сюда же подаются уловленные в циклоне частички культуры, унесенные воздухом. Освободившиеся кюветы проходят через мойку 12, обеспложиваются в стерилизаторе 13 и возвращаются под загрузку.
Готовую культуру гриба упаковывают в бумажные крафт-мешки. Каждый мешок снабжают этикеткой или биркой с указанием завода-изготовителя, наименования продукта, даты выпуска, номера партии, активности ферментов и массы. Хранят культуру в сухом помещении на деревянных стеллажах.
Предложены технологические схемы производства поверхностных культур плесневых грибов с использованием механизированных растильных установок: с вертикальными каналами (установка была смонтирована на Мичуринском экспериментальном заводе и может быть использована на предприятиях средней мощности — до 2 т культуры гриба в сутки); непрерывнодейст-вующей конвейерно-пластинчатой, предназначенной для специализированных ферментных предприятий мощностью более 2 т культуры в сутки.
ГЛУБИННОЕ КУЛЬТИВИРОВАНИЕ МИКРООРГАНИЗМОВ
При глубинном культивировании микроорганизмы развиваются во всем объеме жидкой питательной среды. Так как подавляющее большинство продуцентов ферментов — строгие аэробы, среду интенсивно аэрируют. В микроорганизмах протекают два связанных процесса — синтез биомассы и синтез ферментов.
Питательные среды. Для глубинного культивирования используют жидкие среды, содержащие твердые компоненты. При работе с комплексными средами, основанными на естественном сырье с добавлением отрубей, ростков, кукурузного жмыха, глю-тена, свекловичного жома, спиртовой барды, следят за тем, чтобы не было крупных комочков, так как они затрудняют стерилизацию и могут привести к закупорке коммуникаций. Поэтому перед смешением твердых компонентов необходимо проводить их просеивание или грубую фильтрацию (например, зерно-картофельной барды).
Жидкую часть питательной среды (воду или фильтрат барды) обогащают питательными солями, гидролизатами белков, аминокислотами, источниками витаминов, различными углеводами. Содержание сухих веществ в жидких средах может колебаться от 1,5 до 20 % в зависимости от продуцента и принятого режима культивирования.
В научно-исследовательских организациях постоянно проводится работа по улучшению эффективности производства ферментных препаратов, в основном по повышению активности, зрелой культуры. Это достигается не только селекцией штаммов микроорганизмов, но и совершенствованием условий культивирования, в том числе и изменением состава питательной среды, поэтому приведенные среды могут подвергаться значительным изменениям при корректировании аэрации и других условий культивирования.
Получение и размножение посевного материала. Для засева производственной питательной среды при глубинном культивировании посевной материал готовят глубинным или поверхностным способом. Вид посевного материала зависит от продуцента:
для грибов это вегетативная мицелиальная масса или споронося-щая поверхностная, для бактерий — молодая растущая культура на начальной стадии спорообразования.
Посевной материал получают постадийным увеличением массы культуры продуцента. При небольшой производительности цеха это сводится к одной или двум операциям, а для заводов большой производительности представляет собой многоступенчатый процесс.
В связи с вводом штаммов микроскопического гриба Asp. awamori 466 или ВУД Т-2 усовершенствована и значительно упрощена схема приготовления спорового взамен вегетативного посевного материала. В простейшем виде она выглядит следующим образом. Пробирку с исходной культурой на агаризованной питательной среде пересевают водной суспензией в колбы со стерильными отрубями, которые выдерживают в термостате до полного созревания культуры. Из колб посевной материал в виде водной суспензии спор пересевается непосредственно в производственный ферментер, минуя инокулятор. В зависимости от объема ферментера количество посевных колб увеличивают в 2..4 раза. Спорового посевного материала вполне хватает для того, чтобы вырастить достаточно активную культуру при обычной продолжительности процесса. При этом получаются большой выигрыш в трудоемкости операций и более надежная защита от инфекции, так как в 2 раза сокращаются число операций и продолжительность выращивания посевного материала.
Культивирование на всех стадиях должно проводиться при оптимальной температуре, аэрации и строго определенное время. Если возникают непредвиденные задержки в использовании, то
посевной материал охлаждают до 8... 10 °С и хранят не дольше 4 ч, иначе качество его может резко ухудшиться.
Посевной материал как на отдельных стадиях, так и готовый подвергают тщательному микробиологическому контролю. Он не должен быть инфицирован посторонней микрофлорой, в нем должно содержаться определенное количество спор или клеток на единицу массы, генетически заложенные в нем свойства продуцировать ферменты должны стойко сохраняться.
Производственное культивирование. Известны следующие способы глубинного культивирования: периодический, непрерывно-циклический и непрерывно-проточный.
Периодический способ характеризуется несменяемостью питательной среды в ферментере, состав которой в процессе развития постепенно изменяется. Процесс протекает по-стадийно: в ферментер набирают питательную среду и задают посевной материал; после размножения микроорганизмов и накопления продуктов их обмена зрелую культуру выгружают, а все оборудование, коммуникации и запорные устройства промывают, а затем стерилизуют паром.
При непрерывноциклическом способе микроорганизмы, расположенные на неподвижной насадке в ферментере, омываются средой, протекающей в замкнутом контуре, до полного потребления ими питательных веществ. После этого зрелую культуру выгружают, начиная с последнего, аппараты промывают, стерилизуют и цикл повторяют с последнего головного ферментера. Богатая питательными веществами среда в ходе такой циклической ферментации постепенно истощается; по времени пребывания среды в зоне реакции этот процесс более продолжителен, чем периодический.
Непрерывно-проточный способ культивирования микроорганизмов более совершенен. Суть его заключается в том, что микробная популяция развивается в проточной питательной среде. Способ имеет две разновидности: гомогенно-непрерывный и градиентно-непрерывный. В первом случае выращивание ведут в одном ферментере; при тщательном перемешивании среды и аэрации обеспечивается одинаковое состояние культуры во всем объеме жидкости. В ферментер при этом непрерывно поступает свежая среда, а из него также непрерывно вытекает избыток зрелой культуральной жидкости.
Градиентно-непрерывное культивирование осуществляют в батарее ферментеров, соединенных переточными трубами. Засеянная среда с большим содержанием углеводов и других ингредиентов непрерывно перетекает из одного ферментера в другой и также непрерывно вытекает в виде готовой культуры.
Непрерывным культивированием в проточных средах можно выращивать микроорганизмы в условиях, оптимальных для их стадии развития. При этом такие важные факторы, как концент-
рация питательных веществ, количество продуктов обмена, рН, содержание растворенного кислорода, резко изменяющиеся при периодическом способе культивирования, поддерживаются постоянными на заданном уровне или изменяются по разработанной программе.
Способы непрерывного культивирования в производстве ферментных препаратов для спиртового производства еще находятся в стадии производственных испытаний. Поэтому, к сожалению, в настоящее время на заводах до сих пор применяют периодическое культивирование микроорганизмов.
КОНЦЕНТРИРОВАНИЕ ФЕРМЕНТНЫХ РАСТВОРОВ
Глубинные культуры микроскопических грибов в жидком виде не могут сохраняться длительный срок без потери активности, перевозить их на большие расстояния вследствие значительного содержания в них воды (до 85...90 %) также экономически не всегда оправдано. Поэтому при организации снабжения культурой спиртовых заводов целесообразно ее предварительно концентрировать.
Концентрирование культур можно осуществлять на вакуум-выпарных установках с получением сиропов при температурах, обеспечивающих сохранение ферментов, или на распылительных сушилках с получением порошкообразного ферментного препарата. Однако эти способы сопровождаются значительными потерями активности (до 50 %) и поэтому не нашли распространения на спиртовых заводах. Начиная с семидесятых годов во ВНИИПрБТ был разработан и внедрен на Мичуринском экспе-
риментальном заводе способ концентрирования глубинных культур ультрафильтрацией.
Способ основан на проведении процесса фильтрации через ряд полупроницаемых мембран, в результате чего ферменты и другие высокомолекулярные вещества задерживаются или выводятся из аппарата в виде ультраконцентрата (концентрированного сиропа). Вода и часть низкомолекулярных веществ (пермеат) проходят через мембраны и также удаляются.
Потери активности при ультрафильтрации составляют до 15 % в пересчете на первоначальную активность исходной осветленной культуральной жидкости.
При использовании концентрированных препаратов на станции осахаривания применяют обычное оборудование для дозирования ферментных осахаривающих материалов.
ПОДГОТОВКА КУЛЬТУР МИКРООРГАНИЗМОВ
К ПРИМЕНЕНИЮ ДЛЯ ОСАХАРИВАНИЯ РАЗВАРЕННОЙ МАССЫ
При замене солода глубинной культурой микроскопических грибов в ферментер по окончании ферментации при тщательном перемешивании добавляют 40%-ный раствор формалина из расчета 2 л на 1 м3 культуральной жидкости. После этого культуру перекачивают насосом в расходные баки, из которых она через дозатор непрерывно поступает в осахариватель.
При замене солода сухой поверхностной культурой микроскопических грибов или смесью культур их взвешивают, смачивают водой в соотношении 1:1 и тщательно измельчают на дробилках, применяемых для дробления солода. Измельченную массу направляют в сборники и смешивают с теплой водой (28...30 °С) из расчета 3...4 л на 1 кг исходной культуры. Для стерилизации приливают 20...25 мл 40%-ного раствора формалина на 1 дал суспензии, тщательно перемешивают 15...20 мин, после чего выдерживают в течение 20...30 мин и передают в расходные баки.
Если применяют смесь поверхностной культуры микроскопических грибов и солода, например Asp. awamori или Asp. oryzae и
ячменного или ржаного солода, то их Дробят вместе и приготавливают водную суспензию так же, как солодовое молоко.
Расход поверхностной культуры, удовлетворяющей требованиям стандарта, определяемый по/массе крахмала зерна и картофеля, включая крахмал, в культуре препарата, составляет 5 %, в том числе Глюкаваморина П — 4 %, Амилоризина П — 1 %; при использовании смеси солода и поверхностной культуры: на крахмал картофеля и зерна — 4 % солода и 2 % Глюкаваморина П или 4 % солода и 3 % Амилоризина.
Расход глубинной культуры микроорганизмов, а также концентрированных препаратов на осахаривание рассчитывают по их активности.
Ниже приведен пример с культурой Asp. awamori 466. Эта культура имеет незначительное количество а-амилазы, поэтому ее необходимо применять в смеси с другими источниками а-амилазы. Например, при полной замене солода Глюкаваморином Гх и Амилосубтилином Гх расход ферментов рассчитывают, исходя из норм расхода ферментов в единицах активности на 1 г перерабатываемого крахмала сырья. Причем этот расход изменяется в зависимости от принятой продолжительности брожения.
Так, при 72-часовом брожении расход -амилазы должен составлять 1,5...2 ед. АС, глюкоамилазы — 6 ед. на 1 г крахмала. При 48-часовом брожении расход -амилазы остается неизменным, а глюкоамилазы увеличивается до 15 ед. ГлА на 1 г крахмала. При этом достигаются равные показатели выбраживания.
Аналогично рассчитывают расход и других препаратов микроорганизмов — источников -амилазы. Препарат -амилазы рекомендуется подавать в две точки технологической схемы: 0,5 ед АС — на разжижение подвариваемой массы в предразварник и 1 ед. АС на 1 г крахмала — в осахариватель вместе с глюкоамилазой.
Если препарат -амилазы содержит и другие амилолитические ферменты, в частности глюкоамилазу, то следует их учитывать и соответственно сокращать расход Глюкаваморина.
Так как активность Глюкаваморина Гх, ВУД Т-2 может быть очень высокой (100...250 ед/мл), то для лучшего дозирования его рекомендуется разбавлять водой из расчета 4...8 м3 на 1 м3 препарата.
Препарат Глюкаваморин можно применять и в смеси с солодом. В этом случае в зависимости от расхода солода рассчитывают и расход препарата в единицах ГлА на 1 г крахмала. Так, при 72-часовом брожении и расходе зерна на солод 5 % массы перерабатываемого крахмала необходимо 4 ед. ГлА на 1 г крахмала.
1