Зависимость интенсивности дыхания растительных продуктов от температуры (работа 1)
МОСКОВСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ ПРИКЛАДНОЙ БИОТЕХНОЛОГИИ
КАФЕДРА ХОЛОДИЛЬНОЙ ТЕХНИКИ И ТЕХНОЛОГИИ
ТЕМА КУРСОВОЙ РАБОТЫ:
ЗАВИСИМОСТЬ ИНТЕНСИВНОСТИ ДЫХАНИЯ РАСТИТЕЛЬНЫХ ПРОДУКТОВ ОТ ТЕМПЕРАТУРЫ
ВЫПОЛНИЛ: Воробьев.А.Н
ФАКУЛЬТЕТ:ХТиТ
КУРС: 3
ГРУППА: 6
ПРИНЯЛА: Волдова.Т.А
МОСКВА 2002
СОДЕРЖАНИЕ:
1.Введение.
2.Основные процессы и изменения, происходящие в продуктах растительного происхождения.
3.Процессы и изменения связанные с дыханием.
4.Хранение плодов в регулируемой газовой системе.
5.Охлаждение и хранение плодов и овощей в охлажденном состоянии.
6.Замораживание и хранение продуктов растительного происхождения в замороженном состоянии.
7.Режимы хранения.
8.Быстрое замораживание.
9.Замораживание клеток.
10.Количество вымерзающей воды.
11.Рекристаллизация.
12.Замораживание продуктов бестканевой структуры.
13.Водосодержание, теплоемкость и теплота замерзания некоторых продуктов.
14.Преимущества и недостатки.
15.Список используемой литературы.
1)Введение.
В наше сложное время, с больной кризисной экономикой сохранение пищевых продуктов с наименьшими затратами и с большей эффективностью является одной из главных задач.
Структура питания нуждается в улучшении технологии хранения пищевых продуктов, но улучшить структуру питания нельзя с помощью хладовой обработки, структуру питания можно лишь поддерживать с помощью хладовой обработки, что позволяет поднять качество продуктов растительного происхождения при хранении, и что в свою очередь позволяет увеличить сроки хранения продуктов, а так же стабильность хранения. При холодильной обработке и хранении в пищевых продуктах происходят сложные процессы и явления, приводящие к различным изменениям их структуры. Зависимостью интенсивности дыхания растительных продуктов от температуры, и исследованием влияния охлаждения (замораживания) на пищевые продукты я и решил заняться в данной курсовой работе.
2)Основные процессы и изменения, происходящие в продуктах растительного происхождения.
Для понимания и совершенствования технологических процессов холодильного консервирования возникает настоятельная необходимость изучения изменений, происходящих в плодах и овощах при хранении, в зависимости от вида продукта, действия фитогормонов, устойчивости к фитопатогенным микроорганизмам и физиологическим заболеваниям.
3) ПРОЦЕССЫ И ИЗМЕНЕНИЯ, СВЯЗАННЫЕ С ДЫХАНИЕМ
Условия холодильной обработки и хранения плодов и овощей должны быть таковы, чтобы понизить до минимума интенсивность дыхания, не нарушив при этом естественного течения метаболических процессов, поскольку только продукты растительного происхождения с нормальным дыханием обладают необходимой лежкоспособностью.
Дыхание является основной формой диссимиляции — расщепления органических веществ. Это окислительный процесс, при котором потребляется кислород и выделяется углекислый газ. Обмен углекислого газа и кислорода происходит путем диффузии через устьица и частично через кутикулу.
Главными дыхательными субстратами являются углеводы, жиры и белки. Около '/з количества углеводов в ткани расходуется при дыхании. Жиры и белки используются в меньшей степени, и их участие наиболее характерно для семян.
Расщеплению макромолекулярных субстратов предшествует их гидролиз: поли- и дисахаридов до моносахаридов, жиров до глицерина и жирных кислот, белков до аминокислот.
В результате реакции гликолиза из моносахаридов образуется пировиноградная кислота, затем вследствие окислительного декарбоксилирования пировиноградной кислоты — ацетил-КоА, который является также продуктом последовательной цепи превращений жирных кислот и аминокислот. В цикле Кребса ацетил-КоА подвергается реакциям декарбоксилирования и дегидрирования до полного расщепления. Образующиеся в цикле Кребса изолимонная, -кетоглутаровая и яблочная кислоты являются непосредственными продуктами окисления.
Процессы окисления весьма сложны и осуществляются через многоступенчатую систему специфических ферментативных реакций. Окисление может происходить непосредственным присоединением кислорода к дыхательному субстрату (А+0 → АО ); отщеплением водорода от субстрата (АН +Х → А+ХН ), при этом окисление субстрата АН и образование продукта окисления А происходят в результате восстановления вещества X; удалением электрона от заряженного иона (например, Fe —е → Fe ). В продуктах растительного происхождения происходят окислительные реакции всех трех типов, но процессы второго и третьего типов преобладают, так как они протекают без непосредственного участия кислорода.
В основе современных представлений о механизме окислительных процессов в биологических объектах лежит теория Баха—Палладина, согласно которой дыхание клетки возможно лишь при наличии активированных форм кислорода и водорода, обладающих высокой реакционной способностью.
Активация и передача водорода дыхательного субстрата происходят при участии ферментов дегидрогеназ, характеризующихся высокой специфичностью по отношению к окисляемому субстрату. По природе простетических групп дегидрогеназы делятся на пиридиновые и флавиновые.
Коферментами пиридиновых дегидрогеназ являются НАД-никотинамидадениндинуклеотид и НАДФ-никотинамидаденин-динуклеотидфосфат, выполняющие функции переносчиков водорода. В основе действия пиридиновых дегидрогеназ лежит способность к обратимому гидрированию и дегидрированию пиридинового ядра, входящего в состав коферментов в виде амида никотиновой кислоты—витамина РР. Пиридиновые дегидрогеназы широко распространены в клетках продуктов растительного происхождения и являются в буквальном смысле универсальными окислительными системами.
Флавиновые дегидрогеназы имеют коферменты, представляющие собой нуклеотидные производные рибофлавина — витамина В . Флавиновые ферменты весьма разнообразны, наиболее многочисленная их группа представлена дегидрогеназами, являющимися промежуточными переносчиками водорода в цепи окисления. Дегидрогеназы окисляют восстановленные пиридиновые основания и передают получаемые от них электроны по цепи в направлении к кислороду. Коферментами флавиновых дегидрогеназ, или, как их иногда называют, флавопротеидов (ФП), являются флавинмононуклеотид (ФМН) и флавинаде-ниндинуклеотид (ФАД).
Последними в цепи окисления переносчиками водорода обычно выступают соединения, известные под общим названием коэнзимы Q. Этим наименованием обозначают ряд веществ, являющихся нейтральными липидами и представляющих собой соединение бензохинона с углеводородной боковой цепью, которая состоит из различного числа изопреновых групп. Известны коэнзимы Q с 10, 9, 8 и 7 изопреновыми группами.
Будучи способными обратимо окисляться и восстанавливаться, коэнзимы Q являются связующим звеном в цепи переноса электронов от флавопротеидов к цитохромам.
Активация кислорода в цепи окисления происходит под действием многочисленных ферментов (оксидаз), большинство из которых содержит в качестве простетических групп железопорфириновый комплекс, являясь Fe-протеидами.
В цепи окисления из оксидаз основную роль играют цитохромы, состоящие из специфического белка и железопорфирина. Цитохромы способны обратимо окисляться и восстанавливаться благодаря ионам железа, входящего в их состав. Восстановление окисленных цитохромов происходит вследствие присоединения электронов KoQ либо флавопротеидов. Окисление цитохромов проходит через цепь, состоящую обычно из четырех различных цитохромов. Последним в цепи является цитохром Аз, называемый цитохромоксидазой, окисление которого происходит непосредственно кислородом воздуха.
Как известно, способность какого-либо соединения присоединять или отдавать электроны определяется в основном соотношением величин окислительных потенциалов данного соединения и соединения, с которым оно взаимодействует. Причем реакция проходит тем энергичнее, чем меньше различие между потенциалами взаимодействующих веществ.
К окислительному потенциалу дыхательного субстрата наиболее близки потенциалы, свойственные пиридиннуклеотидам, а к потенциалу кислорода, имеющего наибольшую величину,— потенциал цитохромоксидазы.
Исходя из этого, считают, что первый этап окисления субстратов осуществляется с участием пиридиновых дегидрогеназ, второй — флавиновых дегидрогеназ, третий — коэнзима Q. Причем на этих этапах происходит перенос двух атомов водорода. Далее имеет место перенос непосредственно электронов через систему цитохромов на кислород.
Процесс окисления субстрата в дыхательной цепи сопровождается фосфорилированием АДФ и запасанием энергии в форме АТФ. В цепи окисления есть три участка фосфорилирования: участок окисления флавопротеидов, участок перехода электронов от цитохрома В к цитохрому С и участок перехода электронов от цитохрома А к цитохрому Аз (рис. 1).
Каждая из трех карбоновых кислот, образующихся в цикле Кребса и являющихся основным субстратом в дыхательной цепи, образует три молекулы АТФ.
Аккумуляция и выделение энергии, которая по мере надобности расходуется клеткой, являются основным назначением дыхания. Фосфорилирование неразрывно связано с окислением в дыхательной цепи, поэтому обычно говорят о процессе окисдительного фосфорилирования. Впервые предположение о наличии сопряжения между окислением и
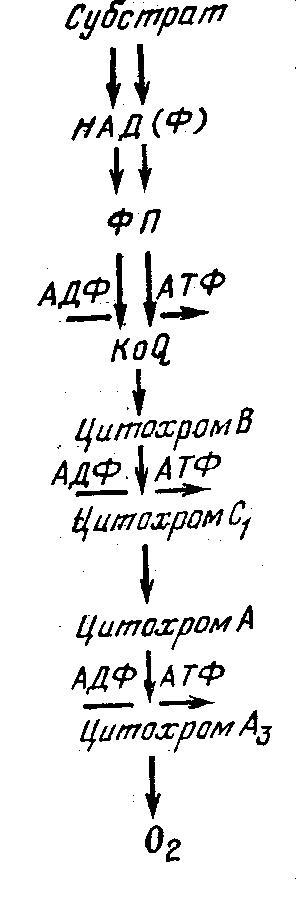
Рис. 1. Схема окисления в дыхательной цепи
фосфорилированием и экспериментальное доказательство этого были даны академиком В. А. Энгельгартом.
Как правило, процессы дыхания происходят в митохондриях клеток. Каждая стадия дыхания осуществляется мультиферментной системой, закрепленной на внутренней митохондриальной мембране. Около 1/4 белков внутренней мембраны являются ферментами, участвующими в переносе электронов и окислительном фосфорилировании. К ним относятся флавопротеиды, цитохромы, ферменты, участвующие в образовании АТФ.
Флавопротеиды и цитохромы располагаются в мембране в виде отдельных групп, каждая из которых содержит все необходимые ферменты и функционирует самостоятельно. Такие группы, называемые дыхательными ансамблями, равномерно распределены по всей плоскости мембраны. Они весьма чувствительны к действию различных внешних воздействий. Под влиянием низких температур, ядов, ионизирууэщей радиации нарушается целостность митохондриальных мембран, и в этом случае сопряжение дыхания утрачивается, или происходит разобщение дыхания. Процессы фосфорилирования более чувствительны. Поэтому при разобщении дыхания окислительные процессы часто не нарушаются, но при этом энергия окисления не превращается в энергию АТФ, а рассеивается в форме тепла.
Разобщение дыхания представляет собой необратимый процесс, являющийся признаком гибели клетки. Однако растительные ткани вследствие наличия у них специальных приспособительных механизмов способны переносить неблагоприятные внешние условия.
Дыхательные процессы, протекающие в клетках, обладают рядом особенностей, обусловленных тем, что продукты растительного происхождения лишены стабильной внутренней среды, например парциального давления кислорода и углекислого газа, температуры, и изменяются под действием непостоянных
и крайне изменчивых факторов внешней среды.
Б. А. Рубин считает, что для структуры аппарата растений характерны три принципиальные особенности:
1. Принцип множественности (мультипринцип) при построении окислительных систем, обусловленный тем, что в клетке имеется, как правило, не один, а несколько ферментов, выполняющих одну и ту же функцию.
2. Полифункциональность каталитических систем, или наличие ферментов, обладающих не одним, а несколькими свойствами.
3. Рассредоточенность (делокализованность) аппарата.
Вследствие этих особенностей процессы окисления биологических объектов могут осуществляться различными путями, или, как говорят, для растительных тканей характерно наличие альтернативных механизмов окислительного обмена, что играет исключительную роль как фактор адаптации, обусловливающий способность тканей сохраняться и нормально функционировать в условиях изменчивой внешней среды, а также при действии различных неблагоприятных факторов.
Схема окисления дыхательного субстрата в клетках разных тканей различна и зависит от их физиологического состояния и условий существования. Так, в отдельные периоды жизненного цикла плодов, например при созревании, клетки нуждаются в дополнительном притоке энергии. При этом в качестве дополнительного дыхательного субстрата используется янтарная кислота, окисление которой происходит более коротким путем, минуя НАД, непосредственно через ФП на КоQ. При этом сукцинатдегидрогеназа, относящаяся к флавиновым ферментам, строго специфична по отношению к янтарной кислоте. Обладая способностью акцептировать водород непосредственно от субстрата и передавать его на КоQ, сукцинатдегидрогеназа образует боковую транспортную цепь окисления.
Кроме того, в клетках имеются альтернативные окислительные механизмы, образующие сопряженные цепи с флавопротеиновыми ферментами и цитохромами. Среди них главную роль играют оксидазы, содержащие в молекуле ионы железа или меди.
В группу Fe-протеидов наряду с флавопротеиновыми ферментами и цитохромами входят каталаза и пероксидаза. В прохождении дополнительного, альтернативного, пути окисления особенно велика роль пероксидаз, разлагающих перекись водорода с освобождением активного атомарного кислорода. Пероксидазы, встречающиеся в клетках продуктов растительного происхождения, весьма многочисленны и окисляют различные вещества, в том числе практически все фенолы, ароматические амины и аскорбиновую кислоту. Их активность возрастает при изменении условий внешней среды. Так, в паренхимной ткани плодов при пониженных температурах хранения или недостаточной обеспеченности кислородом происходит возрастание активности пероксидазы.
К группе Сu-протеидов относятся различные полифеноло-ксидазы и аскорбиноксидаза. В основе их действия лежит обратимое окисление одновалентной меди в двухвалентную.
Полифенолоксидазы представляют собой ферменты, окисляющие в присутствии молекулярного кислорода различные фенолы и их производные с образованием соответствующих хинонов. Полифенолоксидазы катализируют прямое окисление субстратов атмосферным кислородом и участвуют в различных окислительных процессах, в том числе в окислении аминокислот, полипептидов, белков и аскорбиновой кислоты. Активность полифенолоксидаз возрастает при механическом повреждении плодов, воздействии низких температур, кристаллизации воды при замораживании и т. д.
Потемнение мякоти многих плодов (яблок, груш, абрикосов и др.), а также клубней картофеля, наблюдаемое при разрезании, очистке и механическом повреждении, происходит вследствие возрастания активности полифенолоксидаз и нарушения координации реакций окисления и восстановления, в результате чего в ткани накапливаются стойкоокрашенные продукты.
Аскорбиноксидаза обладает строгой специфичностью по отношению к субстрату и окисляет l-аскорбиновую кислоту, превращая ее в дегидроформу. Являясь переносчиком водорода, аскорбиновая кислота тесно связана со всей системой ферментов, участвующих в дыхательных процессах.
Полифенолоксидазы и аскорбиноксидаза принимают участие в окислительных процессах на заключительном этапе дыхания, заменяя цитохромы.
На рис. 2 показаны возможные альтернативные пути процесса окисления, которые в. растительных клетках происходят различным образом в зависимости от видовых особенностей ткани, физиологического состояния данного органа, условий внешней среды и других факторов.
Различная активность оксидаз в условиях измененной газовой среды представлена на рис. 3.
Наибольшее сродство к кислороду имеет цитохромоксидаза, ее действие проявляется при содержании кислорода в атмосфере всего 1 %. Глубинные слои паренхимной ткани плодов и овощей, для которых характерно пониженное содержание кислорода, более богаты цитохромоксидазой, нежели поверхностные.

Рис. 2. Альтернативные пути окисления
Ферменты, участвующие в альтернативных
процессах окисления, локализованы в
различных структурных элементах
клетки. Пероксидазы, полифенолоксидазы,
аскорбиноксидаза сосредоточены
главным образом в цитоплазме; вся
система ферментов, обеспечивающая
окислительное фосфорилирование,
обнаружена в ядрах и хлоропластах.
Благодаря такой делокализации ферментов
дыхательной цепи практически все
структурные составляющие клетки
могут быть источниками того или иного
количества энергии и использоваться
ею в зависимости от условий внешней
среды.
О характере и интенсивности дыхания судят по количеству выделяемого клетками углекислого газа либо по величине дыхательного коэффициента ДК:
ДК=Vсо,/Vо„ где Vco, Vo - объемы выделившегося углекислого газа и поглощенного кислорода.
Величина дыхательного коэффициента определяется прежде всего химической природой субстрата. При окислении углеводов ДК=1 в соответствии с суммарным уравнением окисления глюкозы
С Н О + 60 -> 6СО + 6Н О + Q.
Число выделившихся грамм-молекул углекислого газа равно числу поглощенных грамм-молекул кислорода, а согласно закону Авогадро грамм-молекулы всех газов занимают одинаковый объем.
При дыхании число выделившихся молекул углекислого газа равно числу атомов углерода в молекуле субстрата. Поэтому число использованных молекул кислорода на атом углерода в субстрате возрастает с увеличением числа Н-атомов и снижается с увеличением содержания O-атомов в молекуле субстрата.
Если субстратами дыхания являются высшие карбоновые кислоты или аминокислоты, относительно богатые водородом и бедные кислородом, то ДK<1. Для жиров ДК~0,7, для белков ДК~0,8. В качестве примера приведем окисление стеариновой кислоты:
С Н О + 26O -»18СО + 18Н О; ДК = 18/26 = 0,69.
Если дыхание происходит с использованием низших ди- и трикарбоновых кислот (яблочной, лимонной, винной и щавелевой), богатых кислородом, то ДК>1.
Так, при окислении яблочной кислоты
С Н О + ЗО -» 4СО + ЗН О; ДК. = 4/3 = 1,33.
Интенсивность дыхания и соответственно величина дыхательного коэффициента молодых развивающихся тканей, например меристематических, тканей прорастающих семян выше, чем зрелых плодов. У тканей одного и того же плода или овоща интенсивность дыхания периферических тканей, как правило, выше.
Ниже приведена интенсивность дыхания [в мг СO2/(г*сут)] различных тканей при температуре 15—20 °С.
12,4
69,3
10,6
2,45 11,3 13,8
3,8
Лимон
целые плоды
кожура
мякоть
Картофель
целые клубни
кожица
покоящиеся глазки
мякоть
При понижении температуры интенсивность дыхания плодов снижается и соответственно дыхательный коэффициент уменьшается. Так, по данным Ф. В. Церевитинова, дыхательный коэффициент для яблок сорта Канада составляет 1,14 при 30 °С и 0,88 при 0 °С. У большинства продуктов растительного происхождения дыхание прекращается при температуре около —10 °С. У растений, переносящих в естественных условиях суровые зимы, дыхание отмечается при —20°С и более низких температурах.
Дыхание зависит от газового состава окружающей атмосферы. При понижении концентрации кислорода интенсивность дыхания снижается, при увеличении возрастает. Интенсивность дыхания снижается вследствие увеличения концентрации углекислого газа.
Когда содержание кислорода в окружающей атмосфере уменьшается ниже определенного предела или при полном отсутствии кислорода, клетки продуктов растительного происхождения переходят к так называемому интрамолекулярному или бескислородному дыханию, представляющему собой в химическом отношении брожение.
Брожение наряду с дыханием является основной формой диссимиляции веществ в организме. При дыхании суммарный выход энергии велик, а конечными продуктами являются неорганические соединения углекислого газа и воды. При брожении выход энергии значительно меньше, органический материал не распадается до конца и накапливаются относительно богатые энергией органические вещества.
Для большинства тканей продуктов растительного происхождения характерна способность к спиртовому брожению, для некоторых — к молочнокислому.
Начальные стадии дыхания и брожения одинаковы, но в анаэробных условиях процесс диссимиляции углеводов прекращается на стадии гликолиза при молочнокислом брожении либо на стадии образования ацетальдегида и этанола при спиртовом.
Переход к интрамолекулярному дыханию у растений может быть частичным, что определяется не только составом газовой среды, но и физиологическим состоянием организма. Так, бескислородное дыхание отмечается в период созревания плодов. При полном отсутствии кислорода переход исключительно к интрамолекулярному дыханию приводит к гибели организма, так как накапливающийся этанол губительно действует на живую клетку.
4)ХРАНЕНИЕ ПЛОДОВ В РЕГУЛИРУЕМОЙ ГАЗОВОЙ СРЕДЕ
При хранении плодов практически единственной формой их взаимодействия с окружающей средой является дыхательный газообмен. Поэтому их жизнедеятельность в роцессе хранения в значительной степени определяется составом окружающей атмосферы. Изменяя определенным образом газовую среду, например повышая содержание углекислого газа и уменьшая содержание кислорода, можно снизить интенсивность обменных процессов в хранимых плодах, для того чтобы отделить состояние климактерия и старения плода. На подавлении жизнедеятельности путем создания определенного состава атмосферы основано хранение плодов в регулируемой газовой среде.
Для паренхимной ткани плодов характерно наличие обширных межклеточных пространств, заполненных газом. Газовый состав внутренней атмосферы плодов устанавливается в результате динамического равновесия между интенсивностью потребления кислорода и выделением углекислого газа, с одной стороны, и скоростью диффузии этих газов — с другой. Обмен с окружающей средой происходит через кутикулу, которая затрудняет циркуляцию газов, вследствие чего внутри плода создается своеобразная газовая атмосфера с повышенным содержанием углекислого газа и пониженной концентрацией кислорода. Характерно, что состав атмосферы в межклетниках неодинаков: в расположенных ближе к поверхности межклетниках состав атмосферы более близок к составу окружающей среды. Газовый состав в межклетниках у разных плодов различен, изменяется с их возрастом и зависит от температуры и наружной атмосферы.
Кутикула плодов разных сортов различается по толщине и составу, в результате чего поступление внутрь кислорода и его концентрация в межклетниках и клеточном соке также варьируются. В связи с этим одни плоды будут испытывать недостаток кислорода или избыток углекислого газа, тогда как для других эта же газовая среда окажется благоприятной.
Кислородные оптимумы дыхания неодинаковы на разных этапах развития плода, а также смещаются под воздействием температурного фактора. При 30 °С оптимальной является 5 %-ная концентрация кислорода, при температуре 10 °С и ниже—1 %-ная. Повышенные концентрации углекислого газа особенно благоприятны для задержки обменных процессов в плодах в постклимактерическом состоянии.
Итак, состав газовой среды при хранении плодов должен быть таким, чтобы сохранить нормальный дыхательный газообмен, не нарушить соотношение между аэробной и анаэробной фазами дыхания при одновременном замедлении процессов созревания. При этом необходима правильная корреляция между температурой хранения, концентрациями углекислого газа и кислорода в окружающей атмосфере и состоянием плодов применительно к данному сорту и с учетом места его произрастания, размеров, степени зрелости, районов выращивания, климатических условий года и других факторов.
При хранении, плодов в атмосфере, содержащей повышенные количества углекислого газа, последний проникает в межклетники паренхимной ткани. Благодаря хорошей растворимости углекислого газа в водных растворах и липидах он поступает в клетки и растворяется в клеточном соке. Влияние повышенной концентрации углекислого газа на метаболизм клетки проявляется прежде всего в подавлении дыхания, что объясняется действием углекислого газа на процесс декарбоксилирования яблочной кислоты.
В реакции декарбоксилирования яблочной кислоты до пировиноградной углекислый газ является одним из конечных продуктов. Поэтому избыток его в среде сдвигает равновесие реакции влево, препятствуя расщеплению субстрата. Кроме этого углекислый газ блокирует систему малик-фермента. Он является также конкурентным ингибитором этилена и тормозит созревание плодов.
Углекислый газ стимулирует биосинтез жирных кислот, участвующих в образовании поверхностных восков, что приводит к снижению проницаемости кутикулы.
Являясь антисептиком, углекислый газ задерживает развитие микроорганизмов на поверхности плодов. Наиболее чувствительны к действию углекислого газа плесневые грибы.
Однако значительное повышение содержания углекислого газа может вызвать физиологические заболевания.
Допустимые концентрации углекислого газа находятся в пределах 2,5—10 %. При хранении плодов в атмосфере с повышенным содержанием углекислого газа не рекомендуется поддерживать температуру ниже 2—2,5 °С.
К повышенным концентрациям углекислого газа более чувствительны недозрелые плоды.
При хранении в измененной газовой атмосфере на биохимические процессы в плодах влияют не только повышенные концентрации углекислого газа, но и пониженные концентрации кислорода.
При понижении концентрации кислорода до 3—5 % дыхание ослабляется, но сохраняется его нормальный характер и дыхательный коэффициент равен единице. При дальнейшем понижении содержания кислорода в окружающей плоды атмосфере наблюдаются некомпенсированное выделение углекислого газа плодами и нарушение нормального процесса дыхания.
Влияние пониженного содержания кислорода сказывается в основном на биосинтезе этилена: плоды перестают синтезировать метионин, являющийся предшественником этилена, что приводит к задержке созревания плодов. Пониженные концентрации кислорода вызывают ослабление активности полифенол-оксидазы и снижают устойчивость плодов к действию низких положительных температур.
Таким образом, каждый компонент газовой среды оказывает специфическое воздействие на плоды при хранении, в то же время влияние на биохимические процессы в плодах измененной газовой среды более сложно. Характерно, что газовые смеси, различные по составу, используемые для хранения плодов, не снижают, а лишь отделяют на некоторый срок начало климактерического подъема дыхания: замедляются процессы распада и расходования основных запасных веществ в плодах, уменьшается расход сахара на дыхание, задерживается преобразование крахмала в сахар, тормозится гидролиз протопектина, приостанавливается изменение цвета, так как уменьшается гидролиз хлорофилла, хорошо сохраняется аромат плодов.
Разные виды и сорта плодов неодинаково реагируют на газовый состав.
Более того, в зависимости от условий произрастания один и тот же сорт яблок рекомендуется хранить при различном составе газовой среды. Так, для яблок сорта Джонатан, выращенных в Швейцарии, оптимальной газовой средой являются 4 %-ная концентрация кислорода и 3—4 %-ная концентрация углекислого газа при 4 °С. Яблоки этого же сорта, выращенные в США, рекомендуют хранить при 3 %-ной концентрации кислсрода и 5 %-ной концентрации углекислого газа при 0°С, а выращенные в Голландии—при 13 %-ной концентрации кислорода и 7 %-ной концентрации углекислого газа при 3,5 °С.
Известно, что объем газа, содержащегося в плодах разных сортов, колеблется и, как правило, в позднеспелых сортах он больше, чем у раннеспелых.
Растворимость углекислого газа в клеточном соке зависит от рН, и эта величина у плодов одного сорта, но разных мест произрастания может различаться на 0,5 и даже 1.
По-видимому, сорта плодов, в тканях которых может растворяться углекислый газ в большом количестве, лучше переносят хранение в измененной газовой среде, чем сорта плодов с пониженной растворимостью углекислого газа.
5) ОХЛАЖДЕНИЕ И ХРАНЕНИЕ ПЛОДОВ И ОВОЩЕЙ В ОХЛАЖДЕННОМ СОСТОЯНИИ
Режим хранения охлажденных продуктов растительного происхождения выбирается таким образом, чтобы создать условия для сохранения их жизнеспособности и естественного иммунитета при максимальном снижении интенсивности биохимических процессов и подавлении развития микрофлоры. Как известно, большинство биохимических реакций подчиняются правилу Вант.Гоффа. Для тканей продуктов растительного происхождения, величина Q равняется 1,8—6,8 и составляет для картофеля 1,85, лука репчатого—1,95, капусты белокочанной— 2,18, яблок—2,54, моркови—3,74, черной смородины—6,77. По величине Q можно судить прежде всего об интенсивности снижения окислительных процессов дыхания при хранении. Так, из приведенных данных видно, что пониженные температуры сильнее влияют на снижение дыхания черной смородины и моркови, чем яблок и картофеля. Однако по значению Q трудно судить о других внутриклеточных процессах, происходящих в плодах и овощах, не только из-за сложности биохимических реакций, но и из-за их взаимосвязи со структурными изменениями ткани, неоднозначного влияния пониженных температур на клеточные органоиды, отдельные компоненты клетки и процессы.
Согласно исследованиям Гора зависимость интенсивности дыхания Р от температуры t в интервале от исходной температуры продукта до температуры хранения (порядка 1—2 °С) можно выразить экспоненциальной функцией следующего вида:
P=P exp(kt),
где Р — интенсивность дыхания при 0°С; k — температурный коэффициент интенсивности дыхания.
Величина k для плодов каждого вида и овощей постоянна, по ее значению судят об устойчивости к хранению.
Пониженные температуры оказывают влияние на все структурные элементы клетки продуктов растительного происхождения и прежде всего мембраны. Следует отметить, что мембраны чрезвычайно чувствительны к малейшим изменениям внешней среды. При этом изменяется прежде всего состояние липидов и функциональной воды.
Согласно современным представлениям липиды, составляющие основу мембран, беспрерывно движутся, образуя так называемое липидное море, в котором плавают молекулы белка по отдельности или сгруппированные в определенных сочетаниях. Часть белков в мембране зафиксирована в определенном положении. Поэтому основными структурными компонентами мембран считают липопротеиновые комплексы со встроенными молекулами воды. Роль этой функциональной воды особенно велика: образуя водородные связи между белками и липидами, именно она определяет структуру мембран. Кроме того, эта вода является активным участником биохимических реакций, происходящих в мембранах.
Под влиянием пониженных температур уменьшается подвижность молекул липидов и белка молекул, что является одной из' причин снижения скорости реакций и нарушения структуры мембран, а также отражается на характере происходящих в клетке процессов.
При резком понижении температуры может произойти частичное разобщение дыхания, в результате чего. Возрастет тепловыделение. При пониженных температурах в клетках продуктов растительного происхождения наблюдается развитие альтернативных окислительных процессов дыхания с участием пероксидазы, сукцинатдегидрогеназы, полифенолоксидазы и аскорбиноксидазы. Замедление скорости внутриклеточных реакций при пониженных температурах приводит к снижению интенсивности дыхания. Однако в результате испарения воды дыхание может возрастать. У разных продуктов интенсивность испарения влаги зависит не только от параметров охлаждающей среды, но и от объекта. Большие размеры паренхимных клеток и межклетников, незначительная толщина покровных клеток, большей частью расположенных в один ряд, обусловливают интенсификацию испарения воды тканями продуктов растительного происхождения, особенно овощных культур.
Основная часть воды диффундирует через систему межклетников в направлении к покровной ткани. Даже плоды, покрытые толстым слоем кутикулярных веществ, например цитрусовые, теряют содержащуюся в них влагу в результате испарения.
Испарение влаги при хранении плодов и овощей нарушает нормальное течение обмена веществ в тканях, вызывает ослабление тургора и их увядание. В результате увядания ускоряются процессы распада содержащихся в клетках веществ, увеличивается их расход на дыхание, нарушается энергетический баланс, что приводит к снижению устойчивости плодов и овощей к поражению микроорганизмами и ухудшению качества.
Под влиянием пониженных температур изменяются вязкость и подвижность протоплазмы. Как известно, вязкость протоплазмы клеток продуктов растительного происхождения в 12— 20 раз больше вязкости воды и зависит от процессов жизнедеятельности клетки. При понижении температуры в связи с возрастанием вязкости может произойти нарушение структуры протоплазмы и тем самым жизнеспособности клетки.
Интервал температур, в котором жизнедеятельность клеток продуктов растительного происхождения сохраняется, довольно широк. Но для успешного холодильного консервирования этот интервал сокращается: от температуры замерзания продукта до 11—12 °С.
Стремясь максимально понизить интенсивность процессов и в то же время не нарушить нормальную жизнедеятельность организма растительного происхождения, плоды и овощи, как правило, хранят обычно при температуре, примерно на 1 °С превышающей температуру замерзания. Исключение составляют продукты растительного происхождения, подверженные при пониженных температурах физиологическим заболеваниям, например бананы хранят при 11—13 °С, цитрусовые—при 3— 4 °С.
При хранении в продуктах растительного происхождения продолжаются, но крайне медленно, физиологические процессы. В плодах снижается интенсивность дыхания и отдаляется состояние климактерия (рис. 11). Из рис. 4 видно, что плоды при пониженных температурах сохраняются в течение более длительного времени. В плодах медленно увеличивается содержание Сахаров, снижается содержание органических кислот, происходят процессы, приводящие к улучшению вкуса, аромата, а часто и цвета плода. К концу хранения усиленно расходуются органические кислоты, содержание их в ткани снижается. Особенно уменьшается количество яблочной кислоты. В результате анаэробного дыхания возрастает содержание этилового спирта и ацетальдегида. Так, через 7,5 мес хранения яблок Ренет Симиренко потери Сахаров составили 20 %, органических кислот — 50 % при одновременном увеличении содержания спирта и ацетальдегида в 4—5 раз.
В плодах частично уменьшается содержание аскорбиновой кислоты. Наименьшие потери витамина С отмечены у цитрусовых, причем в мякоти содержание его практически не изменяется. Чем ниже допустимая температура хранения, тем меньше потери витаминов.
При пониженных температурах хранения у овощей большинства видов интенсифицируются процессы расщепления крахмала и образования сахаров. У овощного гороха, фасоли, сахарной кукурузы и некоторых других культур при хранении, наоборот, синтезируется крахмал.
Картофелю особенно свойственно влияние температуры на направленность реакции крахмал ↔ сахар, что необходимо учитывать при разработке условий его хранения. При понижении температуры в клубнях происходит накопление сахаров, а при повышении увеличивается содержание крахмала, что связано с активностью ферментов, катализирующих прямую и обратную реакции и имеющих различную оптимальную температуру действия. С понижением температуры возрастает растворимость углекислого газа во внутриклеточном соке, изменяется рН последнего и возрастает скорость распада крахмала.
6) ЗАМОРАЖИВАНИЕ И ХРАНЕНИЕ ПРОДУКТОВ РАСТИТЕЛЬНОГО ПРОИСХОЖДЕНИЯ В ЗАМОРОЖЕННОМ СОСТОЯНИИ
Основными факторами, определяющими степень обратимости замораживания, являются характер кристаллообразования и локализация льда.
Как правило, первые кристаллы льда образуются в межклетниках. При понижении температуры ниже точки замерзания водяной пар в крупных межклетниках начинает конденсироваться в виде капелек воды на прилегающих клеточных стенках. Эта вода и превращается в первые микроскопические кристаллики льда. Эти кристаллики лъда распространяются по межклетникам, обволакивая стенки клеток. Кристаллики бывают в виде линз или разветвленных кристаллов, разрастающихся между клетками эпидермиса и паренхимы. Далее рост кристалликов происходит за счет воды, содержащейся в. клетках, что объясняется разностью между давлениями пара внутри клетки и вне ее. Активность протекания этого процесса зависит от химического состава, проницаемости клеточных стенок, содержания свободной воды в клетке, вязкости протоплазмы, индивидуальных особенностей ткани, степени за-каливания и др.
Вследствие дегидратации температура замерзания содержимого клеток понижается. При понижении температуры в клетках сначала наступает состояние переохлаждения, а затем в них спонтанно возникают центры кристаллизации, приводящие к образованию внутриклеточного льда.
При понижении температуры вязкость протоплазмы возрастает в результате объединения отдельных белков в длинные цепи, образующие, трехмерную сетчатую структуру. Протоплазма вследствие связывания воды переходит в гелеобразное состояние.
При созревании плодов происходит преобразование протопектина, входящего в состав стенок клеток. Образующийся пектин обладает высокими гидрофильными свойствами: он связывает большие количества воды и способствует образованию гелеобразной структуры, что положительно сказывается на обратимости процесса замораживания. В недозрелых плодах содержится больше свободной воды и происходит в основном внутриклеточная кристаллизация, приводящая к гибели плодов.
Клетки листовой ткани окружены оболочками, состоящими из одного слоя стенок, поэтому эта ткань подвергается разрушительному воздействию отрицательных температур.
Содержащийся в овощах крахмал оказывает определенное влияние на характер кристаллизации. Многие овощи, например лук, картофель, покрыты плотной естественной оболочкой, что способствует переохлаждению, тогда как другие, например капуста белокочанная, не имеющая подобной оболочки, не переохлаждается, что объясняется наличием крупных межклетников и большим содержанием свободной воды.
У клубней картофеля в состоянии покоя повышается газо-и водопроницаемость покровных тканей, что обусловливает большую вероятность внеклеточной кристаллизации льда.
Процесс замораживания, в частности внеклеточная кристаллизация, зависит от холодостойкости и степени закаливания продукта данного вида.
При понижении температуры тканей уменьшается кинетическая энергия молекул, повышается вязкость внутриклеточной жидкости, уменьшаются растворимость газов и диффузия веществ, что в совокупности приводит к снижению скорости химических реакций. В жидкой фазе отмечается повышенная концентрация электролитов, среди которых интерес представляют хлористые соли натрия, аммония, калия и органических веществ (сахаров, нуклёотидов и низкомолекулярных белковых соединений). Концентрированные растворы агрессивны по отношению к белкам, прежде всего ферментам. Поэтому, несмотря на снижение кинетической энергии и ограничение возможности диффузии, некоторые ферментативные реакции в замороженных тканях могут ускоряться либо будет изменяться их течение. Замораживание действует прежде всего на липопротеиновые комплексы. Разрыв водородных связей в совокупности с повышением ионной силы внутриклеточных растворов приводит к разрушению комплексов.
Из ферментов легче других повреждаются те, которые образуют сложные мультиферментные системы, локализованные на внутренних мембранах органоидов клетки: ферментные системы дыхательной цепи и окислительного фосфорилирования митохондрий. При этом нарушаются координация и сбалансированность отдельных реакций, а также их синхронность. Происходит разобщение процессов дыхания (окислительного фосфолирирования), что выражается в утрате организмом основных жизненных функций, а именно дыхания и способности к генерации энергии.
В вакуоли локализована инвертаза, особенностью которой является активность в широком диапазоне рН (3,0—7,5). Поэтому изменение кислотности среды при замораживании не снижает ее активности. Активируемые инвертазой реакции обусловливают накопление сахара в замороженной ткани.
Сохранение активности пектолитических ферментов способствует повышению гидрофильных свойств коллоидов и уменьшению степени повреждения стенок.
Каталаза и пероксидаза катализируют дегидрирование фенолов, аминов, флавонов и аминокислот. Их действие иногда является причиной появления у замороженных плодов и овощей постороннего привкуса. Из этих ферментов пероксидаза более устойчива к действию отрицательных температур.
Существуют и такие ферменты, активность которых повышается при замораживании. К ним относятся полифенолоксидаза и липолитические ферменты. Действие липазы проявляется даже при температуре —40 °С.
Пектолитические ферменты в зависимости от вида продукта оказывают различное действие. Так, в яблоках их активность приводит к размягчению ткани.
7)РЕЖИМЫ ХРАНЕНИЯ
Для каждого вида продукции разработаны оптимальные режимы хранения (табл. 1(а)). Температуру и влажность воздуха в хранилищах измеряют психрометром Августа. В небольших подвалах его размещают в средней части прохода на высоте 1,5 м от пола. В погребах и хранилищах, удаленных от жилища, должно быть четыре-пять таких психрометров. Их располагают в середине, в конце (на расстоянии 1 м от пола) и под коньком.
РЕЖИМ ХРАНЕНИЯ ОВОЩЕЙ И КАРТОФЕЛЯ
|
Продукция |
Тсмпсретура, °С |
Относитсяьн»» влажность воздух», % |
Продолжительность хранения, мес |
|
Томаты: |
|||
|
зеленые |
12...15 |
85—90 |
1—2 |
|
розовые |
8...IO |
85—90 |
До I |
|
красные |
0...2 |
85—90 |
До 0,5 |
|
Перцы |
8...IO |
90—95 |
До 0,5 |
|
Баклажаны |
7...10 |
90—95 |
0,5-0,7 |
|
Морковь |
1...0 |
90-95 |
4—7 |
|
Свекла |
0...1 |
90—95 |
- 8—10 |
|
Редис |
0 |
90—95 |
До 1 |
|
Петрушка |
0...2 |
95 |
2—2,5 |
|
Сельдерей |
- 0.5...0 |
90—95 |
2-4 |
|
Лук (репчатый) |
- 3...0 |
70—80 |
6—8 |
|
Лук-перо |
0 |
90—95 |
До 1 |
|
Чеснок |
- 3...0 |
70—80 |
6-8 |
|
Картофель |
4...5 |
90—95 |
7—8 |
Таблица 1(а).
Осенью, когда температура воздуха бывает повышенной, на ночь для создания сквозняка открывают все окна, вытяжные трубы, люки, двери, а днем их закрывают, чтобы сохранить запас холода. Иногда в погребах отмечается высокая влажность воздуха. Ее признаками служат затхлый спертый воздух, ощущение сырости, появление плесени и конденсированной влаги на потолке, стенах, стеллажах, ящиках.
Чтобы понизить чрезмерную влажность воздуха, одновременно открывают двери и отдушины. Можно поставить в подвал ящик с негашеной известью, солью или древесным углем — эти материалы впитывают воду.
При похолоданиях утепляют крышу. С декабря наступает опасность промерзания хранилища. К этому времени двери, а при необходимости и стены обивают матами и рогожей. Вентиляционные трубы, кроме нужных для работы, плотно закрывают соломой или другими материалами.
8)БЫСТРОЕ ЗАМОРАЖИВАНИЕ
Цель быстрого замораживания, как и любого другого консервирующего процесса, свести на нет или, по крайней мере, замедлить реакции, ухудшающие качество продуктов и в конечном итоге делающие их непригодными к употреблению.
Практически можно говорить только о том, что же преобладает в применяемом процессе. По этому решающему действию или характеру процесса его относят к той или иной группе.
Любой процесс консервирования тем лучше, чем меньшие изменения он вызывает в продуктах в сравнении с их первоначальными свойствами и чем более длительный срок хранения он обеспечивает. В настоящее время из применяемых в промышленных масштабах методов консервирования продуктов процессы холодильной обработки и замораживания лучше всего удовлетворяют упомянутым выше требованиям, поскольку, с одной стороны, они по отношению к другим процессам консервирования вызывают более слабые изменения свойств продуктов, а с другой—обеспечивают достаточно долгую для практических целей сохраняемость пищи.
Процесс консервирования путем холодильной обработки занимает особое место еще и потому, что после охлаждения или замораживания продукты можно сохранять с помощью любого другого процесса консервирования и, наоборот, продукты, законсервированные с помощью других методов, хранятся дольше всего только в охлажденном состоянии;
Процессы, ухудшающие качество продуктов, или процессы, приводящие в конечном итоге к их порче, развиваются в комплексных условиях. Различают физические, биохимические и биологические процессы порчи продуктов. Холодильная обработка независимо от характера обрабатываемого продукта действует на эти процессы с определенной закономерностью.
Из физических процессов порчи к самым основным относятся процессы усушки продуктов или потери массы. Они вызывают не только количественные изменения (уменьшение массы), но и могут привести к ухудшению качества. Потеря массы продуктов подчинена следующей закономерности:
ΔS = Βa(Pп-φp)
где Δ S — теряемое продуктами количество воды за единицу времени, г/ч; Β — коэффициент испарения воды, г/(Н-ч); А—площадь поверхности продуктов, м2;
рп — давление водяных паров при температуре поверхности продуктов, Па; φ — относительная влажность воздуха в виде десятичной дроби; р—давление насыщенных водяных паров при температуре окружающей среды, Па.
При данной поверхности продуктов и данной величине коэффициента испарения потеря массы зависит от величин Pп, φр и р. Если одни и те же продукты с равновесным паросодержанием, например 95%, хранить при различных температурах в помещении с относительной влажностью 80%, то величина Рп—φр будет изменяться следующим образом (при условии, что температура продуктов равна температуре окружающего пространства):
|
Температура, ˚C |
40 |
30 |
20 |
10 |
0 |
-10 |
-20 |
|
Рп—φр |
10.8 |
4.9 |
2.7 |
1.3 |
0.69 |
0.29 |
0.11 |
Таким образом, при прочих равных условиях потеря массы при температуре 40° С примерно в 100 раз больше, чем потеря массы при —20° С, т. е. низкие температуры значительно уменьшают степень усушки продуктов.
Мы говорили уже о том, что потерю массы можно уменьшить с помощью соответствующей упаковки, но даже в этом случае остается явление так называемой внутриупаковочной потери массы.
Биохимические процессы порчи играют исключительно важную роль. Сюда относятся вызванные ферментами катализные процессы распада, включая даже микробиологические процессы порчи, поскольку ясно, что и эти процессы при ближайшем рассмотрении тоже носят биохимический характер. При высоких температурах химические изменения вещества проходят очень быстро, а скорость реакций высока, при низких температурах эти изменения часто едва заметны. Понижение температуры приводит к замедлению постоянных химических реакций, т. е. к уменьшению их скорости. Этот факт выражается законом Аррениуса, который связывает абсолютную температуру Т и логарифм коэффициента химической реакции следующим соотношением:
logK=a—b/T.
Коэффициенты а и b уравнения Аррениуса не зависят от температуры и находятся экспериментальным путем для каждой кон- j кретной реакции. Приведенное соотношение дает возможность вы- i числить скорость химической реакции при любой температуре. На основании этих расчетов и экспериментальных данных можно сделать вывод, что скорость наиболее простых (мономолекулярных, ;
бимолекулярных) реакций при повышении температуры на 10° С увеличивается в 2—3 раза и соответственно понижение температуры на 10° С уменьшает ее до половины или до одной трети. На основании такой закономерности легко видеть, что скорость химических и биохимических реакций сильно замедляется с понижением температуры. Предположим, начальная скорость какой-нибудь реакции v при понижении температуры на 10° С уменьшилась до '/з своей величины, т. е. стала равна v/3. Если температура уменьшится теперь еще на 10° С, то скорость станет равной уже v/(3-3)=v/32 начальной скорости.
Рассуждая подобным образом, увидим, что скорость уменьшается по степенному закону:
v/33, v/3…v/з.
Показатель степени п равен десятой части разности температур реакции. Например, если скорость реакции при 40ْС равна v, то при температуре —20° С она составит уже v/729.
Интересно, однако, заметить, что биохимические реакции проходят на основании этого закона даже при очень низких температурах, только их скорость становится исключительно малой.
Степень снижения скорости биохимических реакций при уменьшении температуры на 10°С, следуя ван Гоффу, принято обозначать
Qio. Степенью Qio принято, однако, описывать снижение скорости не только какой-нибудь одной определенной реакции. Эту величину используют также для характеристики снижения скорости всей совокупности биохимических реакций в целом. Например, поскольку сохраняемость зависит главным образом от действия совокупности биохимических реакций, то срок хранения также может характеризоваться величиной Qio.
Куприянов установил для различных видов пищи увеличение времени ее сохраняемости при изменении температуры на 10° С.
Qio =
где Qio—температурный коэффициент сохраняемости; длительность хранения при t=10°C; —продолжительность хранения при t С.
Как установил Куприянов, при низкотемпературной хранении быстрозамороженных овощей величина Qio равна 2, нежнрной рыбы—3,1, а птицы—4.
Интересная картина получается, если срок хранения какого-нибудь продукта, определенный различными авторами (например, продолжительность хранения говядины или свинины), отложить на графике зависимости от температуры в температурной области ниже и выше точки замерзания (рис. ).
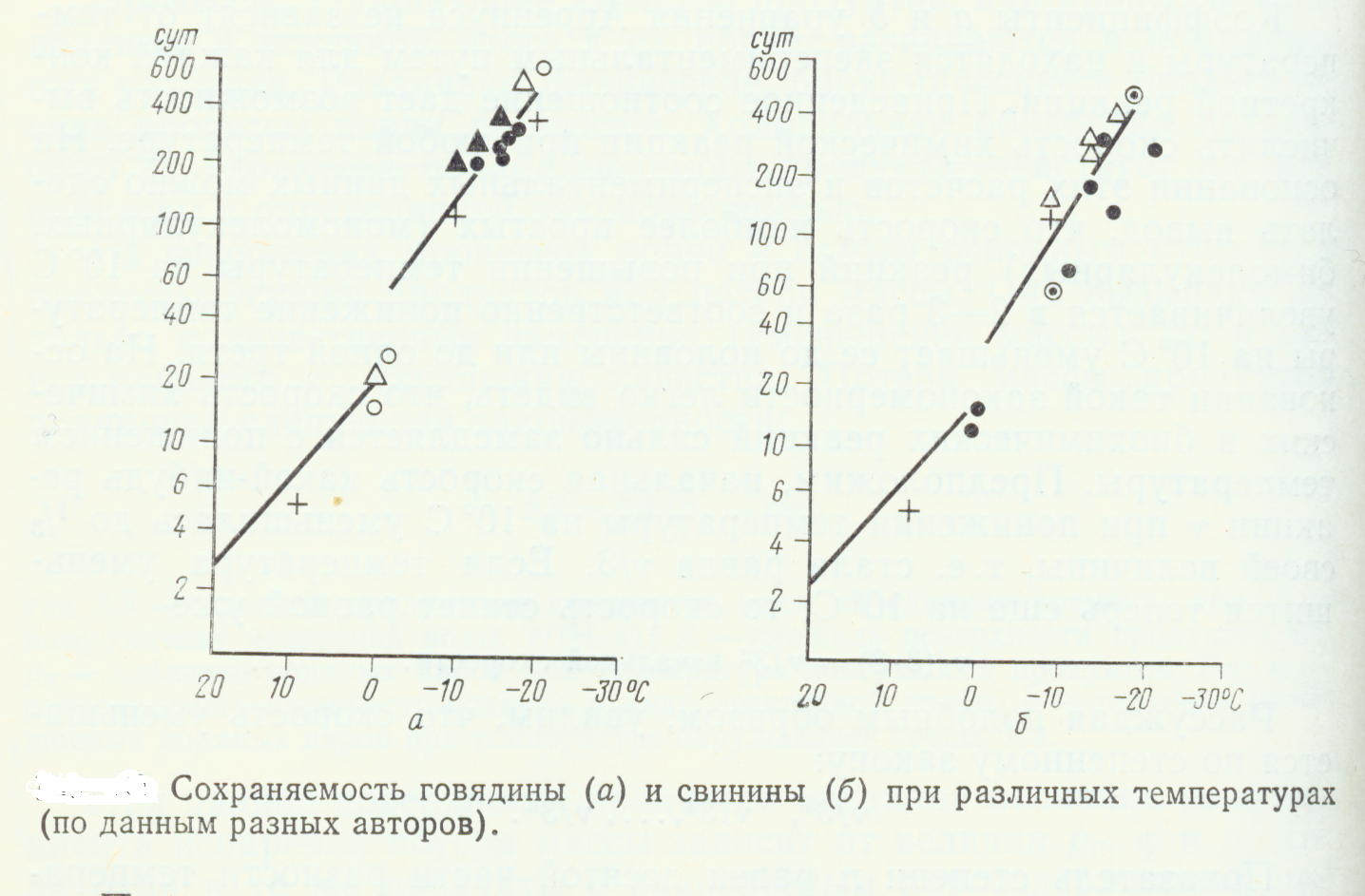
Проделав соответствующие расчеты для значения Qio в области температур выше и ниже точки замерзания, мы получим следующие величины:
Qio=3,85
Qio=5,5
Qio=4,16 Qio=4,85
Говядина выше точки замерзания » ниже » »
Свинина выше точки замерзания » ниже » »
Видно, что при температурах ниже точки замерзания продолжительность хранения увеличивается быстрее при уменьшении температуры, чем в температурной области выше точки замерзания. Таким образом, после перехода через точку замерзания не только уменьшается скорость биохимических реакций, но вступает в силу и новый фактор, влияющий на удлинение срока сохраняемости продуктов. Таким фактором является вымерзание находящейся в продуктах воды (табл. ).

Снижение температуры оказывает многостороннее и комплексное воздействие на биологические процессы. Ниже мы будем рассматривать этот вопрос более подробно. Сейчас только заметим, что применение низких температур приводит к приостановлению и замедлению биологических процессов. С другой стороны, при низких температурах эти процессы могут прекратиться вообще, что вызывает иногда вредные, а иногда полезные изменения.
Во время быстрого замораживания таких влагосодержащих веществ, как пищевые продукты, значительная часть воды становится льдом. Поэтому применение низких температур в подобных процессах консервирования обязательно сопровождается явлением замерзания. Следует, однако, упомянуть, что все-таки есть и такие процессы, где применение низких температур не сопровождается вымерзанием влаги из обрабатываемого продукта. Явление вымерзания удается исключить, когда точка замораживания продуктов сильно понижается с помощью введения в них каких-нибудь веществ, например, сахара или соли. Тогда температура хранения может быть ниже температуры замерзания. Другой пример тому—наличие связанной воды в материале, подвергаемом процессу быстрого замораживания. Связанная вода не вымерзает даже при очень низких температурах, например даже при вымерзании бактериальных спор. Так как процесс замерзания обычно играет важную роль при быстром замораживании продуктов в изменении их качества, мы познакомимся с ним более подробно.
9) ЗАМОРАЖИВАНИЕ КЛЕТОК
Как животные, так и растительные клетки можно заморозить и в живом, и в мертвом состоянии. Замораживаются отдельные клетки в суспензии и клетки, образующие структуру тканей. Однако для всех этих случаев можно установить и одинаково справедливые общие закономерности.
Как известно, химический состав клеток сложен. С точки зрения замораживания главную роль играет поведение клеточных белковых соединений. Благодаря своим физическим свойствам белки относятся к коллоидам. Растворяясь в воде, они образуют гидрогели и гидрозоли. Та часть воды, активность которой в гидрозолях и гидрогелях ниже нормальной, называется связанной водой, а остальное водосодержание—свободной водой. Под действием таких внешних воздействий, как изменение температуры, золи переходят в гели и наоборот.
Изменения в клетках при их охлаждении начинают происходить уже вблизи точки замерзания. Вязкость клеточной протоплазмы возрастает, образуется гель, а содержащиеся в клетках липиды вымерзают. Существенные изменения, однако, начинаются после замерзания некоторого количества воды клеточной влаги. Замерзание клеточного коллоида обычно начинается так же, как и замерзание истинного раствора. Здесь тоже возникает явление понижения температуры замерзания, а из коллоидного раствора вымерзает только чистая вода. Вследствие вымерзания воды остаточная концентрация раствора возрастает, и температура его замерзания подобно истинному раствору уменьшается. Чем ниже температура клеток, тем больше из них вымерзает воды, но благодаря коллоидному характеру системы связанная вода обычно не замерзает. Так, по данным Морана, в 2%-ном растворе желатина примерно 35% воды не вымораживается даже при температуре жидкого воздуха (—196° С).
На рис. температура t соответствует точке замерзания. Ее значение зависит от концентрации растворенных в клеточной влаге веществ. При понижении температуры и дальнейшем отборе тепла некоторая часть воды клеточной влаги вымерзает, но жизнеспособность организма пока не изменяется и не исчезает. Это явление справедливо даже для самых общих случаев. Такое состояние продолжается до температуры t.
Говоря о вымерзании воды на участке t1—t2, мы имели в виду биохимически (и даже биологически) свободную воду. На этом участке скорость замерзания не играет особой роли. Случается даже так, что при медленном замерзании биологическая обратимость оказывается лучшей и большее количество клеток остается в живом состоянии. После оттаивания клеточный обмен веществ снова возобновляется. Тут, однако, становится важным фактор времени, в течение которого клетки находились при температуре t1 до tг, и то, каким образом происходило оттаивание. Значение температуры t2, или границы биологической обратимости, неоди- наково для разных клеток и клеточных структур. Биологическая обратимость при замораживании или полное восстановление жизненных функций после замораживания характерно только для некоторых пород рыб (карасей), некоторых растений и обычно структур более низкого порядка и организации.
При охлаждении ниже температуры t2 процесс вымерзания воды продолжается и, хотя клетки и клеточные структуры отмира-
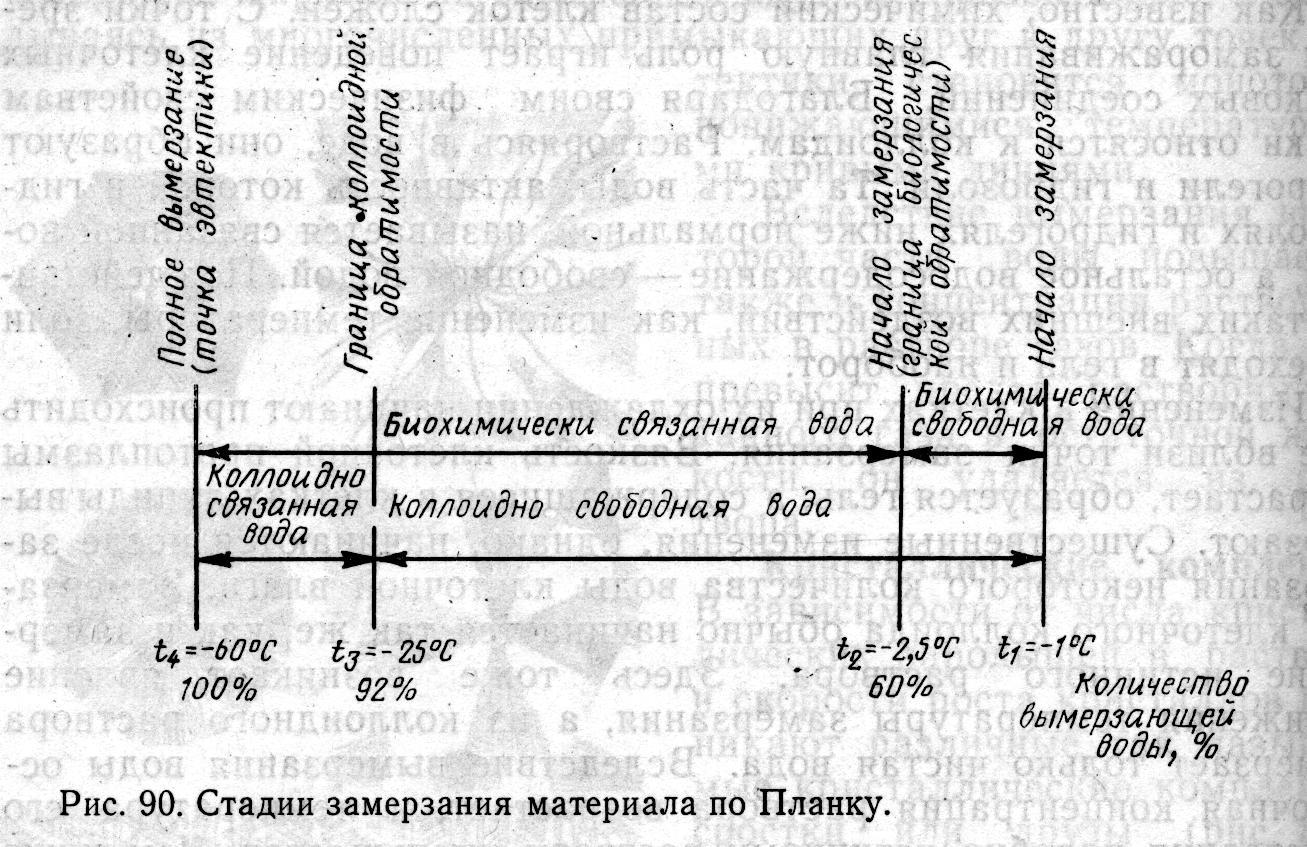
ют, вола вымерзает из коллоидного раствора обратимым образом Вымерзшее количество воды до температуры t3 полностью представлено коллоидно свободной водой. У фруктов температуры t2 и t3 находятся обычно очень близко друг от друга, т. е. прекращений жизнедеятельности практически сопровождается наступлением коллоидной необратимости процесса. Ниже температуры tз начинает замерзать связанная коллоидная вода. Коллоидные частички которые до этого были надежно отделены друг от друга оболочкой гидратов, теперь могут сближаться настолько, что различны силы притяжения склеивают мицеллы. Высвобожденные связи, на которых раньше находились молекулы воды, теперь действуют друг на друга так, что при взаимодействии некоторых главных соединений коллоидных частичек могут произойти химические изменения и коллоид может денатурироваться. Если позже к местам этих связей подойдут молекулы воды, то они не смогут взаимодействовать с ними. Исследования Риделя показали, что даже если температура клеток понизится ниже температуры t3, то и тогда вся вода не вымерзнет из растворов, т. е. содержащуюся в клетках воду невозможно выморозить полностью.
Линг (1968) объясняет состояние связанной воды наличием на| молекулярной поверхности вещества, образующего коллоидный раствор, многослойного поляризованного покрытия из водяных молекул (рис. ). Свойства поляризованных водных молекул (их точка замерзания, способность растворять различные вещества и т.д.) отличаются от свойств нормальных неполяризованных молекул. Можно предположить, что полное влагосодержание живых клеток состоит именно из таких поляризованных молекул.
Уменьшение обратимости коллоидов по мере вымерзания воды ведет к уплотнению остаточного раствора. Водные растворы, образующие клеточную влагу, в зависимости от вида продуктов со-

Рис. 91. Расположение слоев поляризованных молекул воды между молекулами белка. Черточки внутри кружков, обозначающих молекулы воды, показывают направления дипольных моментов.
Рис. 92. Влияние температуры на обратимость замерзания. Кривая / показывает, что уменьшение температуры и увеличение скорости замерзания благоприятно действуют на обратимость, а кривая // показывает, что снижение температуры вследствие концентрации клеточной влаги действует на обратимость неблагоприятным образом. Характеризующая суммарное воздействие этих процессов кривая /// получается суперпозицией кривых / и //.
держат соли, кислоты, сахара, коллоидные белки и т. д. Концентрация образующих электролиты или находящихся в диссоциированном состоянии солей и кислот вызывает коагуляцию или денатурирование белков.
Во время замораживания недостаточно амортизированных систем может изменяться величина рН. Так, например, рН овощей:
при замораживании может уменьшаться, а рН мяса увеличиваться. Эти данные указывают на то, что во время замораживания величина рН может достичь изоэлектрической точки выпадения белков или приблизиться к ней. Изоэлектрическая точка глютенина и яичного белка при рН 4,8, оксигемоглобина—при рН 6,8. С другой стороны, отсюда следует, что путем должной фиксации величины рН можно избежать выделения белков. Последнее удалось подтвердить экспериментально замораживанием мяса с фиксированным значением рН 6,3, после оттаивания которого потеря сока, характеризующая степень денатурирования белков, оказалась очень малой.
10)Количество вымерзающей воды. Вымерзание воды из растительных и животных клеток начинается при температуре их замерзания. По мере отбора тепла вымерзает все больше воды, а концентрация растворенных в клеточной влаге веществ увеличивается. После достижения определенной точки снижать температуру далее бесполезно, поскольку вода из клеток больше не вымерзает. Некоторое ее количество даже при очень низких температурах остается в жидкой фазе. С помощью современного дифференциального термоанализа Дакуорт (1971) установил количество невымерзающей из продуктов воды при температуре —180° С (табл. ).
Наличие в продуктах невымерзшей воды, находящейся в жидком агрегатном состоянии даже при очень низких температурах, указывает на тот факт, что активизация некоторой части водосо-держания клеток сильно затруднена. Независимо от того, вызывается такого рода связанность образованием поляризованной многослойной «шубы» молекул воды или водородными связями, ее энергия достаточно велика. По данным Оленева и Чижова (1973), энергия связи слабосвязанной воды крахмала составляет 155 кДж/кг, энергия промежуточных связей—155—655 кДж/кг, энергия связи сильно связанной и не вымерзающей даже при очень низких температурах воды составляет более чем 655 кДж/кг. Если вспомнить, что удельная теплота испарения воды 2345 кДж/кг, то эти величины представляются не такими уж малыми.
Согласно Дорсею, даже в чистом льде остается невымерзшая жидкая фаза, которая покрывает кристаллики льда пленкой толщиной примерно 8 мкм.
11)Рекристаллизация. Как явствует из изложенного выше, замерзшие клетки и ткани содержат кристаллики льда различных размеров и незамерзшую воду в жидком агрегатном состоянии, т. е. определенный раствор. Наличие кристалликов различной величины связано с разницей скоростей замерзания находящихся на неодинаковой глубине от поверхности продуктов слоев клеток, содержащих воду. Эта гетерогенная система кристалликов разной величины слабо устойчива. Она постоянно изменяется в сторону уменьшения числа кристаллов льда и увеличения объема кристаллов. Процесс рекристаллизации, по нашим современным представлениям, продолжается более или менее длительный промежуток времени до температуры —30° С.
Явление рекристаллизации возникает, как можно предположить, по двум причинам. Первая причина заключается в том, что парциальное давление водяных паров над мелкими кристалликами льда выше, чем над крупными. Вследствие разницы давлений водяных паров молекулы воды постоянно сублимируют с мелких кристалликов и осаждаются на крупных.

Вторая причина—непрерывное колебание температуры хранения продуктов. Поскольку температура плавления мелких кристалликов ниже, чем крупных, при небольшом подогреве они подтаивают в первую очередь. Образовавшаяся от их таяния вода при следующем за небольшим повышением температуры охлаждении намерзает на небольшие кристаллики, увеличивая тем самым их размеры.
Рекристаллизация сильно замедляется при понижении температуры хранения. Разность давлений пара над мелкими и крупными кристалликами при температуре —25°С составляет только '/4 разности давлений при температуре —10° С. Кроме того, колебанию температуры продуктов около 0,05° С, которое возникает в результате колебаний температуры хранилища на 1° С, при охлаждении до —10° С соответствует изменение агрегатного состояния, т.е. расплавление или замерзание примерно 0,04% воды, а при охлаждении до —25° С агрегатное состояние изменяют всего 0,005% воды.
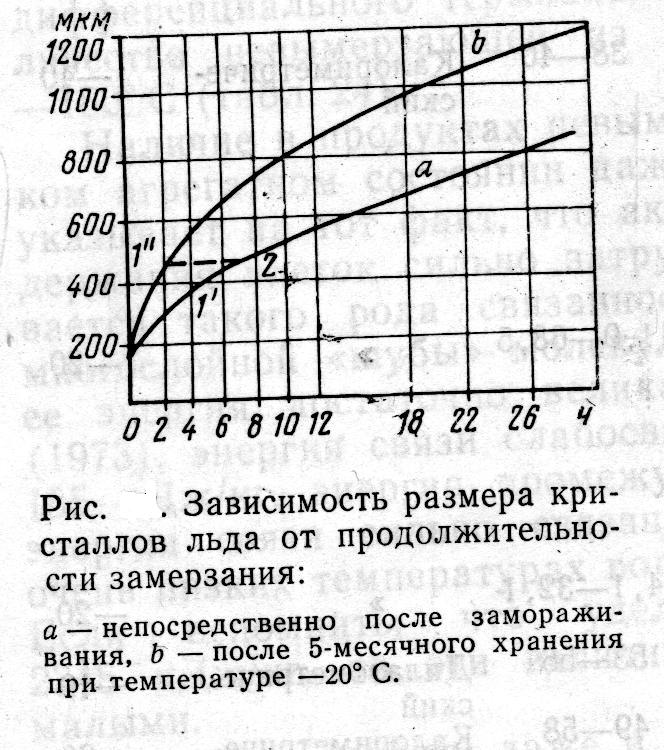
На опыте хорошо видно, как разница в числе и размерах кристалликов льда у быстро и медленно замороженных продуктов постепенно исчезает при хранении.
Очень интересными представляются с этой точки зрения исследования Нотеварпа, который измерял размеры кристаллов льда в замороженной с разной скоростью треске через различные промежутки хранения при температуре —20° С. Например, через 2,5 ч после замораживания (кривая а на рис. ) средние размеры кристаллов составляли 250 мкм. После 5-месячного хранения при температуре —20° С они увеличились до 480 мкм (кривая ///). Из рисунка видно также, что до 480 мкм кристаллы вырастают через 8 ч после замораживания (кривая b).
12)Замораживание продуктов бестканевой структуры. К этой группе можно отнести продукты, которые ведут себя как настоящие растворы, например процеженные, осветленные фруктовые соки и широкий круг продуктов от молока до хлеба, которые проявляют свойства коллоидных растворов или гелей.
На фруктовые соки, кофейные экстракты и т. д., которые можно считать настоящими растворами, замораживание (образование льда), не оказывает никакого влияния. После замораживания, хранения и оттаивания качество продуктов полностью сохраняется. Совсем другое положение возникает тогда, когда целью замораживания является сгущение, криогенное концентрирование. В этом случае исключительно важным с точки зрения следующих фаз технологии, разделения концентрата и кристаллов льда становятся их соответствующие размеры, а в некоторых аспектах' и правильное направление кристаллообразования.
Разделение облегчается, а потери вещества уменьшаются, если в процессе замораживания, проведенного с целью криоконцентри-рования, в растворе образуются большие кристаллы. В этом случае удельная поверхность льда уменьшается, а значит, и становится меньшим количество связанного вследствие адсорбции и трудно отделимого концентрата.
Для облегчения центробежного разделения в процессе Крау-зе—Линде обычно стремятся получить рост кристаллов в радиальном направлении перпендикулярно оси вещества, состоящего из льда и концентрата.
Обратное положение возникает при производстве мороженого, когда существенно, чтобы образующиеся кристаллы льда имели как можно меньшие размеры и давали при потреблении желаемую «гладкую» консистенцию. Эта цель достигается не только посредством быстрого замораживания, но и путем применения различных эмульгаторов (яичный желток, желатин, агар и т. д.), которые при смешивании способствуют образованию мелких ледяных кристаллов.
Можно еще также упомянуть о том, что во время лиофилизации растворов большую роль играют размеры кристаллов льда, возникающих при замораживании перед проведением основной операции. Согласно экспериментальным данным, образование как слишком больших, так и слишком маленьких кристаллов не способствует высококачественному проведению технологического процесса, поскольку маленькие кристаллы затрудняют сублимацию водяного пара, а большие после своего исчезновения оставляют много нежелательных пустот.
Преобладающее большинство принадлежащих к этой группе продуктов являются коллоидными растворами или гелями. Изучением изменения структуры коллоидов при замораживании задалось много исследователей. Современная точка зрения на этот счет может быть сформулирована следующим образом.
Коллоидные растворы под действием замораживания страдают большей частью от необратимых, но частично и от обратимых физических изменений. Такими изменениями может быть дезагрегация, а иногда и агрегация коллоидных частиц.
В жидких растворах наступает главным образом дезагрегация, а в концентрированных растворах — агрегация частиц. Установлено также, что однополярные коллоиды при замораживании в неполярных растворителях агрегируют, а в полярных растворителях дезагрегируют. Дезагрегация обратима.
Гетерополярные коллоиды в полярных растворителях могут покаазать оба этих физических изменения. Агрегация возникает под действием сил Ван-дер-Ваальса, а дезагрегация — под действием ктростатических и дипольных сил. В продуктах коллоидного актера имеет место обычно процесс агрегации. Однако в то время увеличение активности ферментов при замораживании объясняется явлением дезагрегации.
13.Водосодержание, теплоемкость и теплота замерзания некоторых продуктов.
|
Водосодсржание, теплоемкость и теплота замерзания некоторых продуктов |
|||||||
|
Теплоемкость |
|||||||
|
Продукт |
Водосо- Держание, % |
Теплота замерзания или плавления |
|||||
|
выше точки замерзания |
ниже точки замерзания |
||||||
|
са |
|||||||
|
кДж/(кг*К) |
ккал/(кг*град) |
кДж/(кг*К) |
ккал/(кг*град |
кДж/(кг) |
ккал/(кг) |
||
|
Яблоки |
83 |
3,85 |
0,92 |
1,76 |
0,42 |
280,52 |
67 |
|
.Бананы |
75 |
3,35 |
0,80 |
— |
— |
251,21 |
60 |
|
Птица |
173 |
2,93-3,18 |
0.70-0,76 |
1,68 |
0,40 |
247,02 |
59 |
|
Вино |
— |
3,77 |
0,90 |
— |
— |
— |
— |
|
Картофель |
74 |
3,35 |
0,80 |
1,76 |
0,42 |
242,83 |
58 |
|
Лимоны |
83—89 |
3,85 |
0,92 |
1,93 |
0,46 276—297 |
66-71 |
|
|
Сахар |
0,1 |
— |
— |
1,26 |
0,30 |
— |
|
|
Черешня |
82 |
3,64 |
0,87 |
1,84 |
0,44 |
276,33 |
66 |
|
Орехи |
7,2 |
1,05 |
0,25 |
0,92 |
0,22 |
37,68 |
9 |
|
Крыжовник |
90 |
3,85 |
0,92 |
1,93 |
0,46 |
301,45 |
72 |
|
Мороженое |
40—65 |
3,27 |
0,78 |
1,88 |
0,45 |
217,71 |
52 |
|
Чеснок |
91 |
3,89 |
0,93 |
2,01 |
0,48 |
305,64 |
73 |
|
Арбузы |
89 |
3,85 |
0,92 |
1,93 |
0,46 |
297,26 |
71 |
|
Рыба |
|||||||
|
жирная |
60 |
2,85 |
0,68 |
1,59 |
0,38 |
209,34 |
50 |
|
тощая |
73 |
3,43 |
0,82 |
1,80 |
0,43 |
255,34 |
61 |
|
Телятина |
63 |
2,95 |
0.70 |
1,60 |
0,40 |
209,34 |
50 |
|
Говядина |
|||||||
|
тощая |
72 |
3,25 |
0,78 |
1,76 |
0,42 |
234,46 |
56 |
|
упитанная |
51 |
2,55 |
0,61 |
1,49 |
0,36 |
171,66 |
41 |
|
Свинина |
|||||||
|
упитанная |
39—46 |
2,13 |
0,51 |
1,34 |
0,32 129—154 31-37 |
||
|
Бобовые |
89 |
3,85 |
0,92 |
1,97 |
0,47 |
297,26 |
71 |
|
Савойская ка |
91 |
3,89 |
0,93 |
3,01 |
0,48 |
305,64 |
73 |
|
пуста |
|||||||
|
Груши |
83 |
3,85 |
0,92 |
1,76 |
0,42 |
280,52 |
67 |
|
Маргарин |
18—18 |
2,72—2,93 |
0,65—0,70 |
1,47 |
0,35 |
— |
— |
|
Апельсины |
84 |
3,85 - |
0,92 |
1,84 |
0,44 |
284,70 |
68 |
|
Персики |
87 |
3,85 |
0,92 |
1,72 |
0,41 |
293,08 |
70 |
|
Томаты |
94 |
3,89 |
0,93 |
2,05 |
0,49 |
314,01 |
75 |
|
Сыр |
|||||||
|
нежирный |
53 |
2,85 |
0,68 |
1,68 |
0,40 |
175,85 |
42 |
|
жирный |
35—50 |
1,88—2,51 |
0,45—0,60 |
1,26 |
0,30 108 —155 26-37 |
||
|
Морковь |
83 |
3,64 |
0,87 |
1,88 |
0,45 |
276,33 |
66 |
|
Пиво |
89—91 |
3,77 |
0,90 |
— |
— |
301,45 |
72 |
|
Спаржа |
94 |
3,89 |
0,93 |
1,97 |
0,47 |
314,01 |
75 |
|
Сало |
. |
2,30 |
0,55 |
1,30 |
0,31 |
71,18 |
17 |
|
Ежевика |
90 |
3,85 |
0,92 |
1,97 |
0,47 |
299,78 |
72 |
|
Виноград |
81 |
3,68 |
0,88 |
1,88 |
0,45 |
263,77 |
63 |
|
Молоко |
88 |
3,94 |
0,94 |
2,51 |
0,60 |
293,08 |
70 |
|
Сливки, смета |
59 |
3,56 |
0,85 |
1,51 |
0,36 |
196,78 |
47 |
|
на |
|||||||
|
Морские раки |
77 |
3,39 |
0,81 |
1,80 |
0,43 |
259,58 |
62 |
|
Тесто |
— |
1,88 |
0,45 |
— |
— |
— |
— |
|
Яйца |
70 |
3,18 |
0,76 |
1,68 |
0,40 |
234,46 |
56 |
|
Творог |
80 |
2,93 |
0,70 |
1,88 |
0,45 |
— |
— |
|
Масло |
14—15 |
2,51—2,68 |
0,60—0,64 |
1,26 |
0,30 |
— |
- |
Преимущества и недостатки.
Здесь можно отметить, что для каждого продукта есть свои режимы охлаждения
(замораживания) при которых этот продукт пробудет без значительного изменения
своей структуры. Например, при замораживании пищевых продуктов бестканевой
структуры, таких как осветленные фруктовые соки и широкий круг продуктов
от молока до хлеба, которые проявляют свойства коллоидных растворов или
гелей замораживание не оказывает никакого влияния, после замораживания,
хранения и оттаивания качество продуктов полностью сохраняется. В отличии
от пищевых продуктов бестканевой структуры продукты, имеющие тканевую
структуру имеют способность окисляться, развивать микрофлору, что приводит
к нежелательным изменениям в структуре продуктов, а в свою очередь это
приводит к порче продукта. Так же следует учитывать время в течении, которого
тот или иной продукт охлаждается (замораживается), немаловажно учитывать
продолжительность хранения, ведь при хранении происходит испарение пищевых
продуктов. Большую роль играет исходное состояние пищевых продуктов
питания. Скорость охлаждения (замораживания) влияет на исходное состояние
кристаллов перед тем как продукт будет помещен в хранилище, каждому продукту
соответствует своя скорость охлаждения (замораживания) которая формирует
размер кристаллов так чтобы они не повреждали тканевую структуру продукта
и что в свою очередь влияло бы на более долгий срок хранения.
15.Список используемой литературы.
1) Э.Алмаши, Л.Эдели, Т.Шарой “Быстрое замораживание пищевых продуктов”
Н.А.Головкин “Холодильная технология пищевых продуктов”
А.Ф.Намесников “Консервирование плодов и овощей в домашних условиях”