Регуляция дыхания
Федеральное агентство по образованию
ФГОУ ВПО «ЮЖНЫЙ ФЕДЕРАЛЬНЫЙ УНИВЕРСИТЕТ»
Ростовский Государственный Педагогический Университет
Курсовая работа
Тема:
Регуляция дыхания
Дисциплина: Физиология человека
Ростов-на-Дону 2009
Содержание
Введение
1. Дыхательный центр
2. Регуляция деятельности дыхательного центра
3. Рефлексы дыхательного центра и рефлекторное влияние на дыхание
4. Механизм адаптации дыхания к мышечной деятельности
Заключение
Список литературы
Введение
Дыхание – это неотъемлемый признак жизни. Мы дышим постоянно с момента рождения и до самой смерти, дышим днем и ночью во время глубокого сна, в состоянии здоровья и болезни.
В организме человека и животных запасы кислорода ограничены, поэтому организм нуждается в непрерывном поступлении кислорода из окружающей среды. Также постоянно и непрерывно из организма должен удаляться углекислый газ, который всегда образуется в процессе обмена веществ и в больших количествах является токсичным соединением.
Дыхание – сложный непрерывный процесс, в результате которого постоянно обновляется газовый состав крови и происходит биологическое окисление в тканях. В этом заключается его сущность.
Нормальное функционирование организма человека возможно только при условии пополнения энергией, которая непрерывно расходуется. Организм получает энергию за счет окисления органических веществ – белков, жиров, углеводов. При этом освобождается скрытая химическая энергия, которая является источником жизнедеятельности, развития и роста организма. Таким образом, значение дыхания состоит в поддержании в организме оптимального уровня окислительно-восстановительных процессов.
Состав выдыхаемого воздуха весьма непостоянен и зависит от интенсивности обмена веществ, а также от частоты и глубины дыхания. Стоит задержать дыхание или сделать несколько глубоких дыхательных движений, как состав выдыхаемого воздуха изменится.
Важную роль в жизнедеятельности человека играет регуляция дыхания.
Регуляция деятельности дыхательного центра, расположенного в продолговатом мозге, осуществляется гуморально, за счет рефлекторных воздействий и нервных импульсов, поступающих из отделов головного мозга.
В курсовой работе рассмотрены вопросы регуляции деятельности дыхательного центра и механизмы адаптации дыхания к мышечной деятельности.
1. Дыхательный центр
Дыхательным центром называют совокупность нервных клеток, расположенных в разных отделах центральной нервной системы, обеспечивающих координированную ритмическую деятельность дыхательных мышц и приспособление дыхания к изменяющимся условиям внешней и внутренней среды организма.
Некоторые группы нервных клеток являются необходимыми для ритмической деятельности дыхательных мышц. Они расположены в ретикулярной формации продолговатого мозга, составляя дыхательный центр в узком смысле слова. Нарушение функции этих клеток приводит к прекращению дыхания вследствие паралича дыхательных мышц.
Дыхательный центр продолговатого мозга посылает импульсы к мотонейронам спинного мозга, иннервирующим дыхательную мускулатуру.
Мотонейроны, отростки которых образуют диафрагмальные нервы, иннервирующие диафрагму, находятся в передних рогах III…IV шейных сегментов. Мотонейроны, отростки которых образуют межреберные нервы, иннервирующие межреберные мышцы, расположены в передних рогах грудного отдела спинного мозга. Отсюда понятно, что при перерезке спинного мозга между грудными и шейными сегментами прекращается реберное дыхание, а диафрагмальное дыхание сохраняется, так как двигательное ядро диафрагмального нерва, находящееся выше места перерезки, сохраняет связь с дыхательным центром и диафрагмой. При перерезке спинного мозга под продолговатым дыхание полностью прекращается и наступает гибель организма от удушения. Однако при такой перерезке мозга продолжаются в течение некоторого времени сокращения вспомогательных дыхательных мышц ноздрей и гортани, которые иннервируются нервами, выходящими непосредственно из продолговатого мозга.
Уже в древности было известно, что повреждение спинного мозга ниже продолговатого приводит к смерти. В 1812 г. Легаллуа путем перерезки мозга у птиц, а в 1842 г. Флуранс путем раздражения и разрушения участков продолговатого мозга дали объяснение этого факта и привели экспериментальные доказательства местонахождения дыхательного центра в продолговатом мозгу. Флуранс представлял дыхательный центр как ограниченную зону размером с булавочную головку и дал ему название «жизненного узла».
Н. А. Миславский в 1885 г., применяя методику точечного раздражения и разрушения отдельных участков продолговатого мозга, установил, что дыхательный центр расположен в ретикулярной формации продолговатого мозга, в области дна IV желудочка, и является парным, причем каждая его половина иннервирует дыхательные мышцы той же половины тела. Кроме того, Н. А. Миславский показал, что дыхательный центр представляет собой сложное образование, состоящее из центра вдоха (инспираторный центр) и центра выдоха (экспираторный центр). Он пришел к заключению, что определенный участок продолговатого мозга является центром, регулирующим и координирующим дыхательные движения.
Выводы Н.А. Миславского подтверждены многочисленными экспериментальными исследованиями, в частности проведенными в последнее время с помощью микроэлектродной техники. При записи электрических потенциалов отдельных нейронов дыхательного центра обнаружено, что в нем существуют нейроны, разряды которых резко учащаются в фазе вдоха, и другие нейроны, разряды которых учащаются в фазе выдоха. Раздражение отдельных точек продолговатого мозга электрическим током, проводимое с помощью микроэлектродов, также выявило наличие нейронов, стимуляция которых вызывает акт вдоха, и других нейронов, стимулирующих акт выдоха.
Баумгартен в 1956 г. показал, что нейроны дыхательного центра распределены в ретикулярной формации продолговатого мозга, вблизи от striae acusticae (рисунок 1). Точной границы между экспираторными и инспираторными нейронами не существует, но имеются участки, где преобладают одни из них: инспираторные – в каудальном отделе одиночного пучка, (tractus solitarius), экспираторные – в вентральном ядре (nucleus ambiguus).
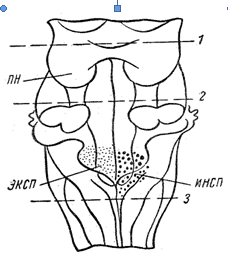
Рисунок 1 – Локализация дыхательных центров На рисунке – нижняя часть ствола мозга (вид сзади). ПН – центр пневмотаксиса; ИНСП – инспираторный центр; ЭКСП – экспираторный центр. Центры являются двусторонними, но для упрощения схемы на каждой из сторон изображен только один из центров. Перерезка выше линии 1 на дыхании не отражается. Перерезка по линии 2 отделяет центр пневмотаксиса. Перерезка ниже линии 3 вызывает прекращение дыхания
Лумсден и другие исследователи в опытах на теплокровных животных нашли, что дыхательный центр имеет более сложную структуру, чем предполагалось ранее. В верхней части варолиева моста находится так называемый пневмотаксический центр, который контролирует деятельность расположенных ниже дыхательных центров вдоха и выдоха и обеспечивает нормальные дыхательные движения. Полагают, что значение пневмотаксического центра состоит в том, что во время вдоха он вызывает возбуждение центра выдоха и, таким образом, обеспечивает ритмическое чередование вдоха и выдоха.
Деятельность всей совокупности нейронов, образующих дыхательный центр, необходима для сохранения нормального дыхания. Однако в процессах регуляции дыхания принимают участие также вышележащие отделы центральной нервной системы, которые обеспечивают тонкие приспособительные изменения дыхания при различных видах деятельности организма. Важная роль в регуляции дыхания принадлежит большим полушариям головного мозга и их коре, благодаря которой осуществляется приспособление дыхательных движений при разговоре, пении, спорте и трудовой деятельности.
Регуляция деятельности дыхательного центра осуществляется гуморально, за счет рефлекторных воздействий и нервных импульсов, поступающих из вышележащих отделов головного мозга.
По И.П. Павлову, деятельность дыхательного центра зависит от химических свойств крови и от рефлекторных влияний, в первую очередь с легочной ткани.
Нейронам дыхательного центра свойственна ритмическая автоматия. Это видно из того, что даже после полного выключения приходящих к дыхательному центру афферентных импульсов в его нейронах возникают ритмические колебания биопотенциалов, которые можно зарегистрировать электроизмерительным прибором. Впервые это явление обнаружил еще в 1882 г. И. М. Сеченов. Много позднее Эдриан и Бутендайк посредством осциллографа с усилителем зарегистрировали ритмические колебания электрических потенциалов в изолированном стволе мозга золотой рыбки. Б. Д. Кравчинский наблюдал подобные ритмические колебания электрических потенциалов, происходящие в ритме дыхания, в изолированном продолговатом мозге лягушки.
Автоматическое возбуждение дыхательного центра обусловлено протекающими в нем самом процессами обмена веществ и его высокой чувствительностью к углекислоте. Автоматия центра регулируется нервными импульсами, приходящими от рецепторов легких, сосудистых рефлексогенных зон, дыхательных и скелетных мышц, а также импульсами из вышележащих отделов центральной нервной системы и, наконец, гуморальными влияниями.
2. Регуляция деятельности дыхательного центра
Дыхательный центр не только обеспечивает ритмическое чередование вдоха и выдоха, но и способен изменять глубину и частоту дыхательных движений, приспосабливая тем самым легочную вентиляцию к текущим потребностям организма. Факторы внешней среды, например состав и давление атмосферного воздуха, окружающая температура, и изменения состояния организма, например при мышечной работе, эмоциональном возбуждении и др., влияя на интенсивность обмена веществ, а, следовательно, потребление кислорода и выделение углекислого газа, действуют на функциональное состояние дыхательного центра. В результате меняется объем легочной вентиляции.
Как и все другие процессы автоматической регуляции физиологических функций, регуляция дыхания осуществляется в организме на основе принципа обратной связи. Это значит, что деятельность дыхательного центра, регулирующего снабжение организма кислородом и удаление образующегося в нем углекислого газа, определяется состоянием регулируемого им процесса. Накопление в крови углекислоты, а также недостаток кислорода являются факторами, вызывающими возбуждение дыхательного центра.
Значение газового состава крови в регуляции дыхания было показано Фредериком путем опыта с перекрестным кровообращением. Для этого у двух собак, находившихся под наркозом, перерезали и соединяли перекрестно их сонные артерии и отдельно яремные вены (рисунок 2) После такого соединения этих и зажатия других сосудов шеи голова первой собаки снабжалась кровью не от собственного туловища, а от туловища второй собаки, голова же второй собаки – от туловища первой.
Если у одной из этих собак зажать трахею и таким образом производить удушение организма, то через некоторое время у нее происходит остановка дыхания (апноэ), у второй же собаки возникает резкая одышка (диспноэ). Это объясняется тем, что зажатие трахеи у первой собаки вызывает накопление СО>2> в крови ее туловища (гиперкапния) и уменьшение содержания кислорода (гипоксемия). Кровь из туловища первой собаки поступает в голову второй собаки и стимулирует ее дыхательный центр. В результате возникает усиленное дыхание – гипервентиляция – у второй собаки, что приводит к снижению напряжения СО>2 >и повышению напряжения О>2> в крови сосудов туловища второй собаки. Богатая кислородом и бедная углекислым газом кровь из туловища этой собаки поступает в голову первой и вызывает у нее апноэ.
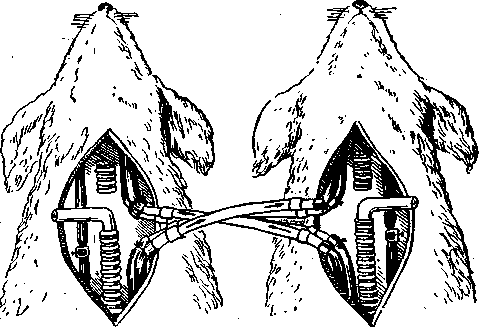
Рисунок 2 – Схема опыта Фредерика с перекрестным кровообращением
Опыт Фредерика показывает, что деятельность дыхательного центра изменяется при изменении напряжения СО>2> и О>2> в крови. Рассмотрим влияние на дыхание каждого из этих газов в отдельности.
Значение напряжения углекислого газа в крови в регуляции дыхания. Повышение напряжения углекислого газа в крови вызывает возбуждение дыхательного центра, приводящее к увеличению вентиляции легких, а понижение напряжения углекислого газа в крови угнетает деятельность дыхательного центра, что приводит к уменьшению вентиляции легких. Роль углекислого газа в регуляции дыхания доказана Холденом в опытах, в которых человек находился в замкнутом пространстве небольшого объема. По мере того как во вдыхаемом воздухе уменьшается содержание кислорода и увеличивается содержание углекислого газа, начинает развиваться диспноэ. Если же поглощать выделяющийся углекислый газ натронной известью, содержание кислорода во вдыхаемом воздухе может снизиться до 12%, причем заметного увеличения легочной вентиляции не наступает. Таким образом, увеличение объема вентиляции легких в этом опыте обусловлено повышением содержания во вдыхаемом воздухе углекислого газа.
В другой серии экспериментов Холден определял объем вентиляции легких и содержание углекислого газа в альвеолярном воздухе при дыхании газовой смесью с разным содержанием углекислого газа. Полученные результаты приведены в таблице 1.
дыхание мышечная газовый кровь
Таблица 1 – Объем вентиляции легких и содержание углекислого газа в альвеолярном воздухе
|
Содержание СО>2>> >во вдыхаемом воздухе, % |
Содержание СО>2>> >в альвеолярном воздухе, % |
Вентиляция легких, % |
|
0,03 3,98 5,28 |
5,71 6,03 6,55 |
100 277 477 |
Данные, приведенные в таблице 1, показывают, что одновременно с увеличением содержания углекислого газа во вдыхаемом воздухе нарастает его содержание в альвеолярном воздухе, а значит, и в артериальной крови. При этом происходит увеличение вентиляции легких.
Результаты экспериментов дали убедительное доказательство того, что состояние дыхательного центра зависит от содержания углекислого газа в альвеолярном воздухе. Выявлено, что увеличение содержания СО>2> в альвеолах на 0,2% вызывает увеличение вентиляции легких на 100%.
Уменьшение содержания углекислого газа в альвеолярном воздухе (и, следовательно, уменьшение напряжения его в крови) понижает деятельность дыхательного центра. Это происходит, например, в результате искусственной гипервентиляции, т. е. усиленного глубокого и частого дыхания, которое приводит к снижению парциального давления СО>2> в альвеолярном воздухе и напряжения СО>2> в крови. В результате наступает остановка дыхания. Пользуясь таким способом, т. е. производя предварительную гипервентиляцию, можно значительно увеличить время произвольной задержки дыхания. Так поступают ныряльщики, когда им нужно провести под водой 2…3 минуты (обычная длительность произвольной задержки дыхания составляет 40…60 секунд).
Прямое возбуждающее действие углекислоты на дыхательный центр доказано путем различных экспериментов. Инъекция 0,01 мл раствора, содержащего углекислоту или ее соль, в определенный участок продолговатого мозга вызывает усиление дыхательных движений. Эйлер подвергал изолированный продолговатый мозг кошки действию углекислого газа и наблюдал, что это вызывает увеличение частоты электрических разрядов (потенциалов действия), свидетельствующее о возбуждении дыхательного центра.
На дыхательный центр оказывает влияние повышение концентрации водородных ионов. Винтерштейн в 1911 г. высказал точку зрения, что возбуждение дыхательного центра вызывает не сама угольная кислота, а, повышение концентрации водородных ионов вследствие увеличения ее содержания в клетках дыхательного центра. Это мнение основывается на том, что усиление дыхательных движений наблюдается при введении в артерии, питающие мозг, не только угольной кислоты, но и других кислот, например молочной. Возникающая при увеличении концентрации водородных ионов в крови и тканях гипервентиляция способствует выделению из организма части содержащейся в крови углекислоты и тем самым приводит к уменьшению концентрации водородных ионов. Согласно этим экспериментам, дыхательный центр является регулятором постоянства не только напряжения углекислоты в крови, но и концентрации водородных ионов.
Установленные Винтерштейном факты нашли подтверждение в экспериментальных исследованиях. Вместе с тем ряд физиологов настаивал на том, что угольная кислота является специфическим раздражителем дыхательного центра и оказывает на него более сильное возбуждающее действие, чем другие кислоты. Причиной этого оказалось то, что углекислый газ легче, чем Н+-ион, проникает через гематоэнцефалический барьер, отделяющий кровь от цереброспинальной жидкости, которая является непосредственной средой, омывающей нервные клетки, и легче проходит через мембрану самих нервных клеток. При поступлении СО>2> внутрь клетки образуется Н>2>СО>3>, которая диссоциирует с освобождением Н+-ионов. Последние и являются возбудителями клеток дыхательного центра.
Другой причиной более сильного по сравнению с другими кислотами действия Н>2>СО>3> является, по мнению ряда исследователей, то, что она специфически влияет на некоторые биохимические процессы в клетке.
Стимулирующее влияние углекислого газа на дыхательный центр является основанием одного мероприятия, нашедшего применение в клинической практике. При ослаблении функции дыхательного центра и возникающем при этом недостаточном снабжении организма кислородом больного заставляют дышать через маску смесью кислорода с 6% углекислого газа. Такая газовая смесь носит название карбогена.
Механизм действия повышенного напряжения СО>2> и увеличенной концентрации Н+-ионов в крови на дыхание. Долгое время считалось, что повышение напряжения углекислого газа и увеличение концентрации Н+-ионов в крови и цереброспинальной жидкости (ликворе) влияют непосредственно на инспираторные нейроны дыхательного центра. В настоящее же время установлено, что изменения напряжения СО>2> и концентрации Н+-ионов действуют на дыхание, возбуждая находящиеся вблизи дыхательного центра хеморецепторы, чувствительные к указанным выше изменениям. Эти хеморецепторы находятся в тельцах диаметром около 2 мм, расположенных симметрично с двух сторон продолговатого мозга на вентролатеральной его поверхности поблизости от места выхода подъязычного нерва.
Значение хеморецепторов продолговатого мозга видно из следующих фактов. При воздействии на эти хеморецепторы углекислого газа или растворов с повышенной концентрацией Н+-ионов наблюдается стимуляция дыхания. Охлаждение одного из хеморецепторных телец продолговатого мозга влечет за собой, согласно опытам Лешке, прекращение дыхательных движений на противоположной стороне тела. Если хеморецепторные тельца разрушены или отравлены новокаином, дыхание прекращается.
Наряду с хеморецепторами продолговатого мозга в регуляции дыхания важная роль принадлежит хеморецепторам, находящимся в каротидном и аортальном тельцах. Это было доказано Геймансом в методически сложных опытах, в которых сосуды двух животных соединялись так, что каротидный синус и каротидное тельце или дуга аорты и аортальное тельце одного животного снабжались кровью другого животного. Оказалось, что увеличение концентрации Н+-ионов в крови и повышение напряжения СО>2> вызывают возбуждение каротидных и аортальных хеморецепторов и рефлекторное усиление дыхательных движений.
Имеются данные, что 35% эффекта, вызываемого вдыханием воздуха с высоким содержанием углекислого газа, обусловлены влиянием на хеморецепторы увеличенной концентрации Н+-ионов в крови, а 65% являются результатом повышения напряжения СО>2>. Действие СО>2> объясняется быстрой диффузией углекислого газа через мембрану хеморецептора и сдвигом концентрации Н+-ионов внутри клетки.
Рассмотрим влияние недостатка кислорода на дыхание. Возбуждение инспираторных нейронов дыхательного центра возникает не только при повышении напряжения углекислого газа в крови, но и при понижении напряжения кислорода.
Пониженное напряжение кислорода в крови вызывает рефлекторное усиление дыхательных движений, действуя на хеморецепторы сосудистых рефлексогенных зон. Прямое доказательство того, что понижение напряжения кислорода в крови возбуждает хеморецепторы каротидного тельца, получено Геймансом, Нилом и другими физиологами путем регистрации биоэлектрических потенциалов в синокаротидном нерве. Перфузия каротидного синуса кровью с пониженным напряжением кислорода приводит к учащению потенциалов действия в этом нерве (рисунок 3) и к учащению дыхания. После разрушения хеморецепторов понижение напряжения кислорода в крови не вызывает изменений дыхания.
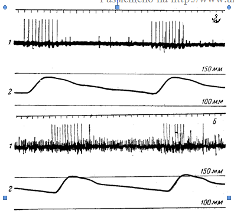
Рисунок 3 – Электрическая активность синусного нерва (по Нилу) А – при дыхании атмосферным воздухом; Б – при дыхании газовой смесью, содержащей 10% кислорода и 90% азота. 1 – запись электрической активности нерва; 2 – запись двух пульсовых колебаний артериального давления. Калибровочные линии соответствуют величине давления 100 и 150 мм рт. ст.
Запись электрических потенциалов Б показывает непрерывную частую импульсадию, возникающую при раздражении хеморецепторов недостатком кислорода. Высокоамплитудные потенциалы в периоды пульсовых повышений артериального давления обусловлены импульсацией прессорецепторов каротидного синуса.
Тот факт, что раздражителем хеморецепторов является понижение напряжения кислорода в плазме крови, а не уменьшение общего содержания его в крови, доказывается следующими наблюдениями Л. Л. Шика. При понижении количества гемоглобина или при связывании его угарным газом содержание кислорода в крови резко уменьшено, но растворение О>2> в плазме крови не нарушено и напряжение его в плазме остается нормальным. При этом возбуждения хеморецепторов не происходит и дыхание не меняется, хотя транспорт кислорода резко нарушен и ткани испытывают состояние кислородного голодания, так как недостаточно кислорода доставляется им гемоглобином. При понижении атмосферного давления, когда уменьшается напряжение кислорода в крови, возникает возбуждение хеморецепторов и учащение дыхания.
Характер изменения дыхания при избытке углекислоты и понижении напряжения кислорода в крови различен. При небольшом понижении напряжения кислорода в крови наблюдается рефлекторное учащение ритма дыхания, а при незначительном повышении напряжения углекислоты в крови происходит рефлекторное углубление дыхательных движений.
Таким образом, деятельность дыхательного центра регулируется воздействием повышенной концентрации Н+-ионов и увеличенного напряжения СО>2> на хеморецепторы продолговатого мозга и на хеморецепторы каротидного и аортального телец, а также действием на хеморецепторы указанных сосудистых рефлексогенных зон понижения напряжения кислорода в артериальной крови.
Причины первого вдоха новорожденного объясняются тем, что в утробе матери газообмен плода происходит через пупочные сосуды, тесно контактирующие с материнской кровью в плаценте. Прекращение этой связи с матерью при рождении приводит к понижению напряжения кислорода и накоплению углекислоты в крови плода. Это, по данным Баркрофта, вызывает раздражение дыхательного центра и приводит к вдоху.
Для наступления первого вдоха важно, чтобы прекращение эмбрионального дыхания произошло внезапно: при медленном зажатии пуповины дыхательный центр не возбуждается и плод погибает, не совершив ни единого вдоха.
Следует учитывать также, что переход в новые условия вызывает у новорожденного раздражение ряда рецепторов и поступление по афферентным нервам потока импульсов, повышающих возбудимость центральной нервной системы, в том числе и дыхательного центра (И. А. Аршавский).
Значение механорецепторов в регуляции дыхания. Дыхательный центр получает афферентные импульсы не только от хеморецепторов, но и от прессорецепторов сосудистых рефлексогенных зон, а также от механорецепторов легких, дыхательных путей и дыхательных мышц.
Влияние прессорецепторов сосудистых рефлексогенных зон обнаруживается в том, что повышение давления в изолированном каротидном синусе, связанном с организмом только нервными волокнами, приводит к угнетению дыхательных движений. Это происходит и в организме при повышении артериального давления. Наоборот, при понижении артериального давления дыхание учащается и углубляется.
Важное значение в регуляции дыхания имеют импульсы, поступающие к дыхательному центру по блуждающим нервам от рецепторов легких. От них в значительной степени зависит глубина вдоха и выдоха. Наличие рефлекторных влияний с легких было описано в 1868 г. Герингом и Брейером и легло в основу представления о рефлекторной саморегуляции дыхания. Она проявляется в том, что при вдохе в рецепторах, находящихся в стенках альвеол, возникают импульсы, рефлекторно тормозящие вдох, и стимулирующих выдох, а при очень резком выдохе, при крайней степени уменьшения объема легких возникают импульсы, поступающие к дыхательному центру и рефлекторно стимулирующие вдох. О наличии такой рефлекторной регуляции свидетельствуют следующие факты:
в легочной ткани в стенках альвеол, т. е. в наиболее растяжимой части легкого, имеются интерорецепторы, представляющие собой воспринимающие раздражения окончания афферентных волокон блуждающего нерва;
после перерезки блуждающих нервов дыхание становится резко замедленным и глубоким;
при раздувании легкого индифферентным газом, например азотом, при обязательном условии целости блуждающих нервов, мускулатура диафрагмы и межреберий внезапно перестает сокращаться, вдох останавливается, не достигнув обычной глубины; наоборот, при искусственном отсасывании воздуха из легкого наступает сокращение диафрагмы.
На основании всех этих фактов авторы пришли к выводу, что растяжение легочных альвеол во время вдоха вызывает раздражение рецепторов легких, вследствие чего учащаются импульсы, приходящие к дыхательному центру по легочным ветвям блуждающих нервов, а это рефлекторно возбуждает экспираторные нейроны дыхательного центра, и, следовательно, влечет за собой возникновение выдоха. Таким образом, как писали Геринг и Брейер, «каждый вдох, поскольку он растягивает легкие, сам подготовляет свой конец».
Если соединить с осциллографом периферические концы перерезанных блуждающих нервов, можно зарегистрировать потенциалы действия, возникающие в рецепторах легких и идущие по блуждающим нервам к центральной нервной системе не только при раздувании легких, но и при искусственном отсасывании из них воздуха. При естественном же дыхании частые токи действия в блуждающем нерве обнаруживаются только во время вдоха; во время же естественного выдоха их не наблюдается (рисунок 4).
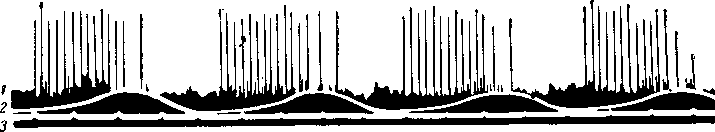
Рисунок 4 – Токи действия в блуждающем нерве при растяжении легочной ткани во время вдоха (по Эдриану) Сверху вниз: 1 – афферентные импульсы в блуждающем нерве: 2 – запись дыхания (вдох – вверх, выдох – вниз); 3 – отметка времени
Следовательно, спадение легких обусловливает рефлекторное раздражение дыхательного центра только при таком сильном их сжатии, какого не бывает при нормальном, обычном выдохе. Это наблюдается лишь при очень глубоком выдохе или внезапном двустороннем пневмотораксе, на что диафрагма рефлекторно реагирует сокращением. Во время естественного дыхания рецепторы блуждающих нервов раздражаются только при растяжении легких и рефлекторно стимулируют выдох.
Помимо механорецепторов легких, в регуляции дыхания принимают участие механорецепторы межреберных мышц и диафрагмы. Они возбуждаются растяжением при выдохе и рефлекторно стимулируют вдох (С. И. Франштейн).
Соотношения между инспираторными и экспираторными нейронами дыхательного центра. Между инспираторными и экспираторными нейронами существуют сложные реципрокные (сопряженные) соотношения. Это означает, что возбуждение инспираторных нейронов тормозит экспираторные, а возбуждение экспираторных нейронов тормозит инспиряторные. Такие явления частично обусловлены наличием прямых связей, существующих между нейронами дыхательного центра, но в основном они зависят от рефлекторных влияний и от функционирования центра пневмотаксиса.
Взаимодействие между нейронами дыхательного центра в настоящее время представляют следующим образом. Вследствие рефлекторного (через хеморецепторы) действия углекислоты на дыхательный центр возникает возбуждение инспираторных нейронов, которое передается на мотонейроны, иннервирующие дыхательные мышцы, вызывая акт вдоха. Одновременно импульсы от инспираторных нейронов поступают к центру пневмотаксиса, расположенному в варолиевом мосту, а от него по отросткам его нейронов импульсы приходят к экспираторным нейронам дыхательного центра продолговатого мозга, вызывая возбуждение этих нейронов, прекращение вдоха и стимуляцию выдоха. Кроме того, возбуждение экспираторных нейронов во время вдоха осуществляется и рефлекторно посредством рефлекса Геринга – Брейера. После перерезки блуждающих нервов приток импульсов от механорецепторов легких прекращается и экспираторные нейроны могут возбуждаться лишь посредством импульсов, приходящих из центра пневмотаксиса. Импульсация, возбуждающая центр выдоха, значительно уменьшается и возбуждение его несколько запаздывает. Поэтому после перерезки блуждающих нервов вдох продолжается значительно дольше и сменяется выдохом позднее, чем до перерезки нервов. Дыхание становится редким и глубоким.
Аналогичные изменения дыхания при целых блуждающих нервах возникают после перерезки ствола мозга на уровне варолиева моста, отделяющей центр пневмотаксиса от продолговатого мозга (см. рисунок 1, рисунок 5). После такой перерезки поступление импульсов, возбуждающих центр выдоха, также уменьшается, и дыхание становится редким и глубоким. Возбуждение центра выдоха в этом случае осуществляется только импульсами, поступающими к нему по блуждающим нервам. Если у такого животного произвести еще и перерезку блуждающих нервов или прервать распространение импульсов по этим нервам путем охлаждения их, то возбуждения центра выдоха не наступает и дыхание останавливается в фазе максимального вдоха. Если после этого восстановить проводимость блуждающих нервов путем согревания их, то вновь периодически возникает возбуждение центра выдоха и восстанавливается ритмическое дыхание (рисунок 6).
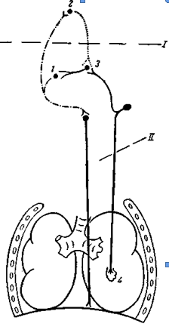
Рисунок 5 – Схема нервных связей дыхательного центра 1– инспираторный центр; 2 – центр пневмотаксиса; 3 –экспираторный центр; 4 – механорецепторы легкого. После перезки по линиям / и // в отдельности ритмическая деятельность дыхательного центра сохраняется. При одновременной перерезке происходит остановка дыхания в фазе вдоха.
Таким образом, жизненно важная функция дыхания, возможная лишь при ритмическом чередовании вдоха и выдоха, регулируется сложным нервным механизмом. При его изучении обращает на себя внимание множественное обеспечение работы этого механизма. Возбуждение центра вдоха возникает как под влиянием увеличения концентрации водородных ионов (повышения напряжения СО>2>) в крови, вызывающего возбуждение хеморецепторов продолговатого мозга и хеморецепторов сосудистых рефлексогенных зон, так и в результате влияния пониженного напряжения кислорода на аортальные и каротидные хеморецепторы. Возбуждение центра выдоха обусловлено как рефлекторными импульсами, приходящими к нему по афферентным волокнам блуждающих нервов, так и влиянием центра вдоха, осуществляемым через центр пневмотаксиса.
Возбудимость дыхательного центра изменяется при действии нервных импульсов, поступающих по шейному симпатическому нерву. Раздражение этого нерва повышает возбудимость центра дыхания, что усиливает и учащает дыхание.
Влиянием симпатических нервов на дыхательный центр отчасти объясняются изменения дыхания при эмоциях.
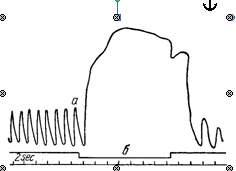
Рисунок 6 – Влияние выключения блуждающих нервов на дыхание после перерезания мозга на уровне между линиями I и II (см. рисунок 5) (по Стелла) а – запись дыхания; б – отметка охлаждения нервов
3. Рефлексы дыхательного центра и рефлекторное влияние на дыхание
На активность нейронов дыхательного центра выраженное влияние оказывают рефлекторные воздействия. Различают постоянные и непостоянные (эпизодические) рефлекторные влияния на дыхательный центр.
Постоянные рефлекторные влияния возникают в результате раздражения рецепторов альвеол (рефлекс Геринга – Брейера), корня легкого и плевры (пульмоторакальный рефлекс), хеморецепторов дуги аорты и каротидных синусов (рефлекс Гейманса), механорецепторов указанных сосудистых областей, проприорецепторов дыхательных мышц.
Наиболее важным рефлексом этой группы является рефлекс Геринга –Брейера. В альвеолах легких заложены механорецепторы растяжения и спадения, являющиеся чувствительными нервными окончаниями блуждающего нерва. Рецепторы растяжения возбуждаются при обычном и максимальном вдохе, т. е. любое увеличение объема легочных альвеол возбуждает эти рецепторы. Рецепторы спадения становятся активными только в условиях патологии (при максимальном спадении альвеол).
В экспериментах на животных установлено, что при увеличении объема легких (вдувание в легкие воздуха) наблюдается рефлекторный выдох, выкачивание же воздуха из легких приводит к быстрому рефлекторному вдоху. Указанные реакции не возникали при перерезке блуждающих нервов. Следовательно, нервные импульсы в центральную нервную систему поступают по блуждающим нервам.
Рефлекс Геринга – Брейера относится к механизмам саморегуляции дыхательного процесса, обеспечивая смену актов вдоха и выдоха. При растяжении альвеол во время вдоха нервные импульсы от рецепторов растяжения по блуждающему нерву идут к экспираторным нейронам, которые возбуждаясь, тормозят активность инспираторных нейронов, что приводит к пассивному выдоху. Легочные альвеолы спадаются, и нервные импульсы от рецепторов растяжения уже не поступают к экспираторным нейронам. Активность их падает, что создает, условия для повышения возбудимости инспипараторной части дыхательного центра и активного вдоха. Кроме того, активность инспираторных нейронов повышается при нарастании концентрации углекислого газа в крови, что также способствует осуществлению акта вдоха.
Таким образом, саморегуляция дыхания осуществляется на основе взаимодействия нервного и гуморального механизмов регуляции активности нейронов дыхательного центра.
Пульмоторакальный рефлекс возникает при возбуждении рецепторов, заложенных в легочной ткани и плевре. Проявляется этот рефлекс при растяжении легких и плевры. Рефлекторная дуга замыкается на уровне шейных и грудных сегментов спинного мозга. Конечным эффектом рефлекса является изменение тонуса дыхательной мускулатуры, благодаря чему происходит увеличение или уменьшение среднего объема легких.
К дыхательному центру постоянно идут нервные импульсы от проприорецепторов дыхательных мышц. Во время вдоха происходит возбуждение проприорецепторов дыхательных мышц и нервные импульсы от них поступают к инспираторным нейронам дыхательного центра. Под влиянием нервных импульсов активность инспираторных нейронов тормозится, что способствует наступлению выдоха.
Непостоянные рефлекторные влияния на активность дыхательных нейронов связаны с возбуждением разнообразных по своим функциям экстеро- и интерорецепторов.
К непостоянным рефлекторным воздействиям, оказывающим влияние на активность дыхательного центра, относятся рефлексы, возникающие при раздражении рецепторов слизистой оболочки верхних дыхательных путей, носа, носоглотки, температурных и болевых рецепторов кожи, проприорецепторов скелетных мышц, интерорецепторов. Так, например, при внезапном вдыхании паров аммиака, хлора, сернистого ангидрида, табачного дыма и некоторых других веществ происходит раздражение рецепторов слизистой оболочки носа, глотки, гортани, что приводит к рефлекторному спазму голосовой щели, а иногда даже мускулатуры бронхов и рефлекторной задержке дыхания.
При раздражении эпителия дыхательных путей накопившейся пылью, слизью, а также попавшими химическими раздражителями и инородными телами наблюдаются чиханье и кашель. Чиханье возникает при раздражении рецепторов слизистой оболочки носа, а кашель – при возбуждении рецепторов гортани, трахеи, бронхов.
Кашель и чиханье начинаются с глубокого вдоха, который возникает рефлекторно. Затем происходит спазм голосовой щели и одновременно активный выдох. Вследствие этого давление в альвеолах и воздухоносных путях значительно возрастает. Следующее за этим раскрытие голосовой щели приводит к выбросу воздуха из легких толчком в дыхательные пути и наружу через нос (при чиханье) или через рот (при кашле). Пыль, слизь, инородные тела увлекаются этой струей воздуха и выбрасываются из легких и дыхательных путей.
Кашель и чиханье в условиях нормы относят к категории защитных рефлексов. Эти рефлексы называют защитными потому, что они препятствуют попаданию вредных веществ в дыхательные пути или же способствуют их удалению.
Раздражение температурных рецепторов кожи, в частности холодовых, приводит к рефлекторной задержке дыхания. Возбуждение болевых рецепторов кожи, как правило, сопровождается учащением дыхательных движений.
Возбуждение проприорецепторов скелетных мышц обусловливает стимуляцию акта дыхания. Повышенная активность дыхательного центра в этом случае является важным приспособительным механизмом, обеспечивающим увеличенные потребности организма в кислороде при мышечной работе.
Раздражение интерорецепторов, например механо-рецепторов желудка при его растяжении, приводит к торможению не только сердечной деятельности, но и дыхательных движений.
При возбуждении механорецепторов сосудистых рефлексогенных зон (дуга аорты, каротидные синусы) в результате изменения величины артериального давления наблюдаются сдвиги в активности дыхательного центра. Так, повышение артериального давления сопровождается рефлекторной задержкой дыхания, понижение приводит к стимуляции дыхательных движений.
Таким образом, нейроны дыхательного центра чрезвычайно чувствительны к воздействиям, обусловливающим возбуждение экстеро-, проприо- и интерорецепторов, что приводит к изменению глубины и ритма дыхательных движений в соответствии с условиями жизнедеятельности организма.
На активность дыхательного центра оказывает влияние коры головного мозга. Регуляция дыхания корой больших полушарий имеет свои качественные особенности. В опытах с прямым раздражением электрическим током отдельных областей коры головного мозга было показано выраженное влияние ее на глубину и частоту дыхательных движений. Результаты исследований М. В. Сергиевского и его сотрудников, полученные при непосредственном раздражении различных участков коры больших полушарий электрическим током в острых, полухронических и хронических опытах (вживленные электроды), свидетельствуют о том, что нейроны коры не всегда оказывают однозначное влияние на дыхание. Конечный эффект зависит от ряда факторов, главным образом от силы, продолжительности и частоты применяемых раздражений, функционального состояния коры головного мозга и дыхательного центра.
Важные факты были установлены Э. А. Асратяном и его сотрудниками. Было обнаружено, что у животных с удаленной корой головного мозга отсутствовали приспособительные реакции внешнего дыхания на изменения условий жизнедеятельности. Так, мышечная активность у таких животных не сопровождалась стимуляцией дыхательных движений, а приводила к длительной одышке и дискоординации дыхания.
Для оценки роли коры головного мозга в регуляции дыхания большое значение имеют данные, полученные с помощью метода условных рефлексов. Если у человека или животных звук метронома сопровождать вдыханием газовой смеси с повышенным содержанием углекислого газа, то это приведет к увеличению легочной вентиляции. Через 10…15 сочетаний изолированное включение метронома (условный сигнал) вызовет стимуляцию дыхательных движений – образовался условный дыхательный рефлекс на избранное количество ударов метронома в единицу времени.
Учащение и углубление дыхания, которые наступают до начала физической работы или спортивных состязаний, также осуществляются по механизму условных рефлексов. Эти изменения в дыхательных движениях отражают сдвиги в активности дыхательного центра и имеют приспособительное значение, способствуя подготовке организма к выполнению работы, требующей большой затраты энергии и усиления окислительных процессов.
По мнению М.Е. Маршака, корковая: регуляция дыхания обеспечивает необходимый уровень легочной вентиляции, темп и ритм дыхания, постоянство уровня углекислого газа в альвеолярном воздухе и артериальной крови.
Приспособление дыхания к внешней среде и сдвигам, наблюдаемым во внутренней среде организма, связано с обширной нервной информацией, поступающей в дыхательный центр, которая предварительно перерабатывается, главным образом в нейронах моста мозга (варолиев мост), среднего и промежуточного мозга и в клетках коры головного мозга.
Таким образом, регуляция активности дыхательного центра сложна. По М.В. Сергиевскому, она состоит из трех уровней.
Первый уровень регуляции представлен спинным мозгом. Здесь располагаются центры диафрагмальных и межреберных нервов. Эти центры обусловливают сокращение дыхательных мышц. Однако этот уровень регуляции дыхания не может обеспечить ритмичную смену фаз дыхательного цикла, так как огромное количество афферентных импульсов от дыхательного аппарата, минуя спинной мозг, направляется непосредственно в продолговатый мозг.
Второй уровень регуляции связан с функциональной активностью продолговатого мозга. Здесь находится дыхательный центр, который воспринимает разнообразные афферентные импульсы, идущие от дыхательного аппарата, а также от основных рефлексогенных сосудистых зон. Этот уровень регуляции обеспечивает ритмичную смену фаз дыхания и активность спинномозговых мотонейронов, аксоны которых иннервируют дыхательную мускулатуру.
Третий уровень регуляции – это верхние отделы головного мозга, включающие и корковые нейроны. Только при наличии коры полушарий головного мозга возможно адекватное приспособление реакций системы дыхания к изменяющимся условиям существования организма.
4. Механизм адаптации дыхания к мышечной деятельности
Интенсивность дыхания тесно связана с интенсивностью окислительных процессов: глубина и частота дыхательных движений уменьшаются при покое и увеличиваются при работе, притом тем сильнее, чем напряженнее работа. Так, у тренированных людей при напряженной мышечной работе объем легочной вентиляции возрастает до 50 и даже до 100 л в минуту.
Одновременно с усилением дыхания во время работы наступает усиление деятельности сердца, приводящее к увеличению минутного объема кровотока. Вентиляция легких и минутный объем кровотока нарастают в соответствии с величиной выполняемой работы и усилением окислительных процессов.
У человека потребление кислорода составляет в покое 250…350 мл в минуту, а во время работы может достигать 4500…5000 мл. Транспорт такого большого количества кислорода возможен потому, что при работе систолический объем может увеличиваться втрое (с 70 до 200 мл), а частота сердечных сокращений в 2 и даже 3 раза (с 70 до 150 и даже 200 сокращений в минуту).
Вычислено, что при повышении потребления кислорода при мышечной работе на 100 мл в минуту минутный объем кровотока возрастает примерно на 800…1000 мл. Увеличению транспорта кислорода при тяжелой мышечной работе способствует также выбрасывание эритроцитов из кровяных депо и обеднение крови водой вследствие потения, что ведет к некоторому сгущению крови и повышению концентрации гемоглобина, а, следовательно, и к увеличению кислородной емкости крови.
Значительно увеличивается при работе коэффициент утилизации кислорода. Из каждого литра крови, протекающей по большому кругу, клетки организма утилизируют в покое 60…80 мл кислорода, а во время работы – до 120 мл (кислородная емкость 1 л крови равна около 200 мл О>2>).
Повышенное поступление кислорода в ткани при мышечной работе зависит от того, что понижение напряжения кислорода в работающих мышцах, увеличение напряжения углекислого газа и концентрации Н+-ионов в крови способствуют увеличению диссоциации оксигемоглобина. Особенно значителен прирост утилизации кислорода у тренированных людей. Крог объяснял это еще и тем, что у тренированных людей во время работы происходит раскрытие большего количества капилляров, чем у нетренированных.
Одной из причин увеличения легочной вентиляции при интенсивной мышечной работе является накопление молочной кислоты в тканях и переход ее в кровь. Содержание молочной кислоты в крови может достигать при этом 50…100 и даже 200 мг % вместо 5…22 мг % в условиях мышечного покоя. Молочная кислота вытесняет угольную кислоту из ее связей с ионами натрия и калия, что приводит к повышению напряжения углекислого газа в крови и к возбуждению дыхательного центра.
Накопление молочной кислоты при мышечной работе возникает потому, что интенсивно работающие мышечные волокна испытывают недостаток в кислороде и часть молочной кислоты не может окислиться до конечных продуктов – углекислого газа и воды. Такое состояние Хилл назвал кислородной задолженностью. Оно возникает при весьма интенсивной мышечной работе, например у спортсменов во время напряженных соревнований.
Окисление образовавшейся во время работы мышц молочной кислоты завершается уже после окончания работы – во время восстановительного периода, в течение которого сохраняется интенсивное дыхание, достаточное для того, чтобы излишние количества накопившейся в организме молочной кислоты были ликвидированы.
Накопление в организме молочной кислоты – не единственная причина усиления дыхания и кровообращения при работе мышц. Как показали исследования М. Е. Маршака, мышечная работа ведет к усилению дыхания даже в том случае, если у человека, работающего на эргометриеском велосипеде, конечности перетянуты жгутом, препятствующим поступлению молочной кислоты и других продуктов из работающих мышц в кровь. Усиление дыхания возникает при этом рефлекторным путем. Сигналом, вызывающим усиление дыхания и кровообращения, является возникающее при сокращении раздражение проприорецепторов мышц. Этот рефлекторный компонент принимает участие в любом усилении дыхания при мышечной работе.
Таким образом, усиление вентиляции при мышечной работе обусловлено, с одной стороны, химическими изменениями, происходящими в организме, – накоплением углекислоты и недоокисленных продуктов обмена, а с другой – рефлекторными влияниями.
Значительную роль в координации функций органов и физиологических систем при мышечной работе играет кора головного мозга. Так, в предстартовом состоянии у спортсменов отмечается увеличение силы и частоты сердечных сокращений, возрастает легочная вентиляция, повышается кровяное давление. Следовательно, условнорефлекторный механизм – один из важнейших нервных механизмов адаптации организма к меняющимся условиях внешней среды.
Система дыхания обеспечивает возросшие потребности организма в кислороде. Системы же кровообращения и крови, перестраиваясь на новый функциональный уровень, способствуют транспорту кислорода к тканям и углекислого газа к легким.
Заключение
Как и все другие процессы автоматической регуляции физиологических функций, регуляция дыхания осуществляется в организме на основе принципа обратной связи. Это значит, что деятельность дыхательного центра, регулирующего снабжение организма кислородом и удаление образующегося в нем углекислого газа, определяется состоянием регулируемого им процесса. Накопление в крови углекислоты, а также недостаток кислорода являются факторами, вызывающими возбуждение дыхательного центра.
В регуляции дыхания важное значение имеет газовый состав крови.
На дыхательный центр оказывает влияние повышение концентрации водородных ионов. В настоящее же время установлено, что изменения напряжения СО>2> и концентрации Н+-ионов действуют на дыхание, возбуждая находящиеся вблизи дыхательного центра хеморецепторы, чувствительные к указанным выше изменениям. Эти хеморецепторы находятся в тельцах диаметром около 2 мм, расположенных симметрично с двух сторон продолговатого мозга на вентролатеральной его поверхности поблизости от места выхода подъязычного нерва.
Пониженное напряжение кислорода в крови вызывает рефлекторное усиление дыхательных движений, действуя на хеморецепторы сосудистых рефлексогенных зон. Деятельность дыхательного центра регулируется воздействием повышенной концентрации Н+-ионов и увеличенного напряжения СО>2> на хеморецепторы продолговатого мозга и на хеморецепторы каротидного и аортального телец, а также действием на хеморецепторы указанных сосудистых рефлексогенных зон понижения напряжения кислорода в артериальной крови.
Дыхательный центр получает афферентные импульсы не только от хеморецепторов, но и от прессорецепторов сосудистых рефлексогенных зон, а также от механорецепторов легких, дыхательных путей и дыхательных мышц.
Влияние прессорецепторов сосудистых рефлексогенных зон обнаруживается в том, что повышение давления в изолированном каротидном синусе, связанном с организмом только нервными волокнами, приводит к угнетению дыхательных движений. Это происходит и в организме при повышении артериального давления. Наоборот, при понижении артериального давления дыхание учащается и углубляется.
Важное значение в регуляции дыхания имеют импульсы, поступающие к дыхательному центру по блуждающим нервам от рецепторов легких. От них в значительной степени зависит глубина вдоха и выдоха.
Помимо механорецепторов легких, в регуляции дыхания принимают участие механорецепторы межреберных мышц и диафрагмы. Они возбуждаются растяжением при выдохе и рефлекторно стимулируют вдох.
Таким образом, жизненно важная функция дыхания, возможная лишь при ритмическом чередовании вдоха и выдоха, регулируется сложным нервным механизмом.
На активность дыхательного центра оказывает влияние коры головного мозга. Для оценки роли коры головного мозга в регуляции дыхания большое значение имеют данные, полученные с помощью метода условных рефлексов.
Учащение и углубление дыхания, которые наступают до начала физической работы или спортивных состязаний, также осуществляются по механизму условных рефлексов.
Эти изменения в дыхательных движениях отражают сдвиги в активности дыхательного центра и имеют приспособительное значение, способствуя подготовке организма к выполнению работы, требующей большой затраты энергии и усиления окислительных процессов.
Список литературы
Анатомия, физиология, психология человека. Краткий иллюстрированный словарь / Под ред. А.С. Петрова. – СПб: Питер, 2002. – 256 с., ил.
Атлас по нормальной физиологии. Пособие для студ. мед. и биол. спец. вузов / Под ред. Н.А. Агаджаняна. – М.: Высш. шк., 1986. – 351 с., ил.
Бабский Е.Б., Забков А.А., Косицкий Г.И., Ходоров Б.И. Физиология человека / Под ред. Е.Б. Бабского. – М.: Издательство «Медицина, 1972. – 656 с.
Сапин М.Р. Анатомия и физиология человека: Уч. Для студ. Образоват.учреждений / М.Р. Сапин, В.И.Сивоглазов. – 6-е изд., стер. –М.: Издательский центр «Академия», 2008. – 384 с.: ил.
Физиология человека /С.А. Георгиева, Н.В. Белинина, Л.И. Прокофьева, Г.В. Коршунов, В.Ф. Киричук, В.М. Головченко, Л.К.Токарева. – М.: Медицина, 1989. – 480 с.
Фомин Н.А. Физиология человека: Уч. пособие для студентов фак. физ. воспитания пед. ин-тов. – М.: Просвещение, 1982. – 320 с., ил.