Vectors for moleculars cloning
National university of life and environmental sciences of ukraine
Chair of molecular genetics and biosafety
Term paper
Vectors For The Molecular cloning
done by:
third year student
group №2
department of ecology
and biotechnology
Pereguda Olga
Scientific advisor
Professor Starodub N. F.
Kyiv 2010
Abstract
Term paper on “Vectors for the moleculars cloning” consist of two sections: conclusions list and of the references.
The object of research:different vectors for the moleculars cloning.
The tasks of term paper:
Learned the vectors for the moleculars cloning
Consider and study vectors of molecular cloning, and functions, properties etc.
The results presented in the form of conclusions at the end of term paper.
Contents
Key words
Abstract
Conditional shortenings
Introducting
Literature review
1. Plasmid vectors
2. Cosmids
3. Phagemids
4. Bacteriophage vectors
4.1. Filamentous phage
4.2.Double-stranded phage
5. Scope of Present Review
6. Life cycle and genetics of Lambda
6.1. Development of Lambda
7. Phage Lambda as a vector
7.1. Size Limitation for Packaging
7.2. Transfection of Recombinant Molecules
7.3. Biological Containment
8. Phage vectors
8.1.Replacement Vectors
8.2. Insertion Vectors
8.3.Storage of Lambda Stocks
Conclusion
Literature
Key words
Cosmids - an extrachromosomal circular DNA molecule that combines features of plasmids and phage; cloning limit - 35-50 kb.
DNA – a long chain polymer of deoxyribonucleotides. DNA constitutes the genetic material of most known organisms and organelles, and usually is in the form of a double helix, although some viral genomes consist of a single strand of DNA, and others of a single- or a double-stranded RNA.
Enzyme – a biological catalyst, usually a protein, that can speed up a chemical reaction by lowering it’s energy of activation without being used up in the reaction. Helicase – a type of enzyme that breaks hydrogen bonds between complementary base pairs of DNA, thereby causing the double strand to spit into separate single strands.
Molecular cloning – is process of creating an identical copy of DNA fragments. Phage - derivatives of bacteriophage lambda; linear DNA molecules, whose region can be replaced with foreign DNA without disrupting its life cycle. Plasmid - an extrachromosomal circular DNA molecule that autonomously replicates inside the bacterial cell.
Promoter - a specific DNA sequence that serves as a binding site for RNA polymerase near each gene.
Replicon – a block of DNA between two adjacent replication origins.
Vector – is an agent that can carry out a DNA fragment into a host cell.
Conditional shortenings
BAC – Bacterial Artificial Chromosome
cos – cohesice end site
DNA - deoxyribonucleic acid
Kb – Kilobases
Kbp – Kilobase pair
nt - necleotides
PCR – Polymarase chain reaction
pUc, pBluscript – phagemid vectors
RNA – ribonucleic acid
Sp6, T7 - promoters
Introduction
Cloning - is the process of creating an identical copy of something. In Biology, it collectively refers to processes used to create copies of DNA fragments (Molecular Cloning), cells (Cell Cloning), or organisms. The term also encompases situations, whereby organisms reproduce asexually, but in common parlance refers to intentionally created copies of organisms.
In 1972, Paul Berg and colleagues made the first “artificial” recombinant DNA molecule. The molecular analysis of DNA has been made possible by the cloning of DNA. The two molecules that are required for cloning are the DNA to be cloned and a cloning vector.
Cloning vector - a DNA molecule that carries foreign DNA into a host cell, replicates inside a bacterial (or yeast) cell and produces many copies of itself and the foreign DNA. Types of Cloning Vectors are Plasmid, Phage, Cosmids.
Molecular cloning refers to the process of making multiple molecules. Cloning is commonly used to amplify DNA fragments containing whole genes, but it can also be used to amplify any DNA sequence such as promoters, non-coding sequences and randomly fragmented DNA. It is used in a wide array of biological experiments and practical applications ranging from genetic fingerprinting to large scale protein production. Occasionally, the term cloning is misleadingly used to refer to the identification of the chromosomal location of a gene associated with a particular phenotype of interest, such as in positional cloning. In practice, localization of the gene to a chromosome or genomic region does not necessarily enable one to isolate or amplify the relevant genomic sequence. To amplify any DNA sequence in a living organism, that sequence must be linked to an origin of replication, which is a sequence of DNA capable of directing the propagation of itself and any linked sequence. However, a number of other features are needed and a variety of specialised cloning vectors (small piece of DNA into which a foreign DNA fragment can be inserted) exist that allow protein expression, tagging, single stranded RNA and DNA production and a host of other manipulations.
Cloning of any DNA fragment essentially involves four steps
fragmentation - breaking apart a strand of DNA
ligation - gluing together pieces of DNA in a desired sequence
transfection - inserting the newly formed pieces of DNA into cells
screening/selection - selecting out the cells that were successfully transfected with the new DNA
Recombinant DNA techniques have allowed the isolation and propagation of specific DNA fragments which can be easily sequenced and/or used as highly specific probes. In vitro site-directed modifications of these fragments and their reintroduction into the genome result in a modified genetic makeup of an organism. In addition, it is now possible to induce overproduction of commercially important proteins by genetically tailored microorganisms[8].
Several cloning strategies have been developed to meet various specific requirements. Cloning protocols have been designed for a variety of host systems. However, Escherichia coli still remains the most popular host of choice since its genetics, physiology, and molecular biology have been studied in great detail and a wealth of information is readily available. Many cloning vectors have also been constructed for use with E. coli as a host. Although this review focuses on the basic and applied aspects of bacteriophage lambda ectors, an overview of other vectors is included for comparison.
In general, cloning vectors can be broadly classified as plasmid and phage vectors.
So, the aim of this work is: to consider and study vectors of molecular cloning, and functions, properties etc.
Literature review
Plasmids are useful for a wide range of molecular genetic, genomic and proteomic approaches. In recent years, plasmid clone production has increased dramatically in response to the availability of genome information and new technologies.[9]
In 1952, Joshua Lederberg coined the term plasmid to describe any bacterial genetic element that exists in an extrachromosomal state for at least part of its replication cycle. As this description included bacterial viruses, the definition of what constitutes a plasmid was sub>sequently refined to describe exclusively or predominantly extrachromosomal genetic elements that replicate autonomously. [1]
Most plasmids possess a circular geometry, there are now many examples in a variety of bacteria of plasmids that are linear. As linear plasmids require specialized mechanisms to replicate their ends, which circular plasmids and chromosomes do not, linear plasmids tend to exist in bacteria that also have linear chromosomes [1]
Plasmids, like chromosomes, are replicated during the bacterial cell cycle so that the new cells can each be provided with at least one plasmid copy at cell division [1]
Frederick Twort (1915) and Felix d’Herelle (1917) were the first to recognize viruses which infect bacteria, which d'Herelle called bacteriophages (eaters of bacteria). [7]
Lambda (λ) bacteriophages are viruses that specifically infect bacteria. The genome of λ-phage is a double-stranded DNA molecule approx 50 kb in length. In bacterial cells, λ-phage employs one of two pathways of replication: lytic or lysogenic. [2]
In lytic growth, approx 100 new virions are synthesized and packaged before lysing the host cell, releasing the progeny phage to infect new hosts. In lysogeny, the phage genome undergoes recombination into the host chromosome, where it is replicated and inherited along with the host DNA. [2]
Cosmids - an extrachromosomal circular DNA molecule that combines features of plasmids and phage. [8]
Cosmids are conventional vectors that contain a small region of bacteriophage λ DNA containing the cohesive end site (cos). This contains all of the cis-acting elements for packaging of viral DNA into λ particles [4]
Plasmid Vectors
In 1952, Joshua Lederberg coined the term plasmid to describe any bacterial genetic element that exists in an extrachromosomal state for at least part of its replication cycle. As this description included bacterial viruses, the definition of what constitutes a plasmid was sub>sequently refined to describe exclusively or predominantly extrachromosomal genetic elements that replicate autonomously.
Figure 1. Joshua Lederberg

Plasmid vectors are convenient for cloning of small DNA fragments for restriction mapping and for studying regulatory regions. However, these vectors have a relatively small insert capacity. Therefore, a large number of clones are required for screening of a single-copy DNA fragment of higher eukaryotes. Second, the handling and storage of these clones is time-consuming and difficult. The repeated sub>cultures of recombinants may result in deletions in the inserts.
The plasmid vectors can be of three main types:
generalpurpose cloning vectors,
expression vectors,
promoter probe or terminator probe vectors.
Figure 2. Cloning into a plasmid

General-purpose cloning vectors
Cloning of foreign DNA fragments in general-purpose cloning vectors [11] selectively inactivates one of the markers (insertional inactivation) or derepresses a silent marker (positive selection) so as to differentiate the recombinants from the native phenotype of the vector.
Expression vectors
In expression vectors, DNA to be cloned and expressed is inserted downstream of a strong promoter present in the vector. The expression of the foreign gene is regulated by the vector promoter irrespective of the recognition of its own regulatory sequence.
Promoter probe and terminator probe vectors
Promoter probe and terminator probe vectors are useful for the isolation of regulatory sequences such as promoters or terminators and for studying their recognition by a specific host. They possess a structural gene devoid of the promoter or the terminator sequence [8].
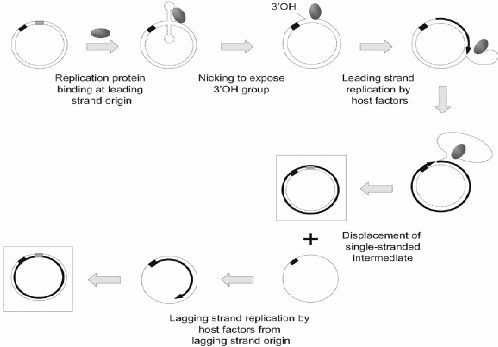
Figure 3. Replication of rolling-circle plasmids
Cosmids
A cosmid, first described by Collins and Hohn in 1978, is a type of hybrid plasmid (often used as a cloning vector) that contains cos sequences, DNA sequences originally from the Lambda phage. Cosmids can be used to build genomic libraries.
Cosmids are able to contain 37 to 52 kb of DNA, while normal plasmids are able to carry only 1–20 kb. They can replicate as plasmids if they have a suitable origin of replication: for example SV40 ori in mammalian cells, ColE1 ori for double-stranded DNA replication or f1 ori for single-stranded DNA replication in prokaryotes. They frequently also contain a gene for selection such as antibiotic resistance, so that the transfected cells can be identified by plating on a medium containing the antibiotic. Those cells which did not take up the cosmid would be unable to grow.
Unlike plasmids, they can also be packaged in phage capsids, which allows the foreign genes to be transferred into or between cells by transduction. Plasmids become unstable after a certain amount of DNA has been inserted into them, because their increased size is more conducive to recombination. To circumvent this, phage transduction is used instead. This is made possible by the cohesive ends, also known as cos sites. In this way, they are similar to using the lambda phage as a vector, but only that all the lambda genes have been deleted with the exception of the cos sequence.
Cos sequences are ~200 base pairs long and essential for packaging. They contain a cosN site where DNA is nicked at each strand, 12bp apart, by terminase. This causes linearization of the circular cosmid with two "cohesive" or "sticky ends" of 12bp. (The DNA must be linear to fit into a phage head.) The cosB site holds the terminase while it is nicking and separating the strands. The cosQ site of next cosmid (as rolling circle replication often results in linear concatemers) is held by the terminase after the previous cosmid has been packaged, to prevent degradation by cellular DNases.
Figure 4. Cloning by using Cosmid method

Cosmid features and uses
Cosmids are predominantly plasmids with a bacterial oriV, an antibiotic selection marker and a cloning site, but they carry one, or more recently two cos sites derived from bacteriophage lambda. Depending on the particular aim of the experiment broad host range cosmids, shuttle cosmids or 'mammalian' cosmids (linked to SV40 oriV and mammalian selection markers) are available. The loading capacity of cosmids varies depending on the size of the vector itself but usually lies around 40–45 kb. The cloning procedure involves the generation of two vector arms which are then joined to the foreign DNA. Selection against wildtype cosmid DNA is simply done via size exclusion. Cosmids, however, always form colonies and not plaques. Also the clone density is much lower with around 105 - 106 CFU per µg of ligated DNA.
After the construction of recombinant lambda or cosmid libraries the total DNA is transferred into an appropriate E.coli host via a technique called in vitro packaging. The necessary packaging extracts are derived from E.coli cI857 lysogens (red- gam- Sam and Dam (head assembly) and Eam (tail assembly) respectively). These extracts will recognize and package the recombinant molecules in vitro, generating either mature phage particles (lambda-based vectors) or recombinant plasmids contained in phage shells (cosmids). These differences are reflected in the different infection frequencies seen in favour of lambda-replacement vectors. This compensates for their slightly lower loading capacity. Phage library are also stored and screened easier than cosmid (colonies!) libraries.
Target DNA: the genomic DNA to be cloned has to be cut into the appropriate size range of restriction fragments. This is usually done by partial restriction followed by either size fractionation or dephosphorylation (using calf-intestine phosphatase) to avoid chromosome scrambling, i.e. the ligation of physically unlinked fragments.
Phagemids
Phagemids combine desirable properties of both plasmids and filamentous phages. They carry
the ColEl origin of replication,
a selectable marker such as antibiotic resistance,
the major intergenic region of a filamentous phage .
The segments of foreign DNA cloned in these vectors can be propagated as plasmids. When cells harboring these plasmids are infected with a suitable helper bacteriophage, the mode of replication of the plasmid changes under the influence of the gene II product of the incoming virus.
Interaction of the intergenic region of the plasmid with the gene II protein initiates the rolling-circle replication to generate copies of one strand of the plasmid DNA, which are packaged into progeny bacteriophage particles. The single-stranded DNA purified from these particles is used as a template to determine the nucleotide sequence of one strand of the foreign DNA segment, for site-directed mutagenesis or as a strand-specific probe. Phagemids provide high yields of double-stranded DNA and render unnecessary the time-consuming process of sub>cloning DNA fragments from plasmids to filamentous bacteriophages.
Bacteriophage Vectors
Both single-stranded (filamentous) and double-stranded E.coli phages have been exploited as cloning vectors.
Frederick Twort (1915) and Felix d’Herelle (1917) were the first to recognize viruses which infect bacteria, which d'Herelle called bacteriophages (eaters of bacteria). [7]
Figure 5. Frederick Twort and Felix d’Herelle


4.1 Filamentous phages
Filamentous phages are not lytic. They coexist with the infected cells for several generations and are convenient for cloning genes which produce toxic products. Among the filamentous phages, fd, fl, and M13 have been well characterized and their genomes have been sequenced [4]. Their gene functions and molecular mode of propagation are very similar. They infect cells via F pili, and the first mature phage appears within 15 min [6].
Phage M13 is widely used in nucleotide sequencing and site-directed mutagenesis since its genome can exist either in a single-stranded form inside a phage coat or as a doublestranded replicative form within the infected cell. During replication, only the plus strand of the replicative form is selectively packaged by the phage proteins [1]. The replicative form is a covalently closed circular molecule and hence can be used as a plasmid vector and transformed into the host by the usual transformation procedures. The vectors derived from M13, have the same polylinker as that of pUC18 and pUC19, respectively [2]. The DNA fragments having noncomplementary ends can be directionally cloned in this pair of vectors, and the two strands of DNA can be sequenced independently.
4.2.Double-stranded phage vectors
Of the double-stranded phages, bacteriophage lambda-derived vectors are the most popular tools for several reasons:
acceptance by the phage of large foreign DNA fragments, thereby increasing the chances of screening a single clone carrying a DNA sequence corresponding to a complete gene;
development and availability of refined techniques aimed at minimizing the problems of background due to nonrecombinants;
the possibility of screening several thousand clones at a time from a single petri plate; and, finally,
the ease with which the phage library can be stored as a clear lysate at 4°C for months without significant loss in plaque-forming activity [7].
Recently, a bacteriophage P1 cloning system has been developed which permits cloning of DNA fragments as large as 100 kbp with an efficiency that is intermediate between cosmids and yeast artificial chromosomes .
Scope of Present Review
The extensive knowledge of the basic biology of lambda has permitted modifications of its genome to suit the given experimental conditions. In the present review we describe how the utility of lambda as a cloning vector rests essentially in its intrinsic molecular organization. The following sections give an account of various problems encountered in constructing lambda vectors and the strategies that have been adopted to overcome them. A few commonly used vectors are described in detail, taking into account their special values and limitations. The different methods for screening and storage of genomic and cDNA libraries in lambda vectors are also discussed.
Life cycle and genetics of Lambda
An understanding of the basic biology of lambda, its mode of propagation, and the genetic and molecular mechanisms that control its life cycle is needed before its applications for genetic manipulations are discussed. This section deals with the basic biology of lambda.
The lambda virus particle contains a linear DNA of 48,502 bp with a single-stranded 5' extension of 12 bases at both ends; these extensions are complementary to each other.
These ends are called cohesive ends or cos. During infection, the right 5' extension (cosR), followed by the entire genome, enters the host cell. Both the cos ends are ligated by E. coli DNA ligase, forming a covalently closed circular DNA which is acted upon by the host DNA gyrase, resulting in a supercoiled structure.
6.1 Development of Lambda
Two Alternative Modes. After infecting the host, the lambda genome may start its replication; this results in the formation of multiple copies of the genome. The protein components necessary for the assembly of mature phage particles are synthesized by the coordinated expression of phage genes. Phage DNA is packaged inside a coat, and the mature phages are released into the environment after cell lysis. This mode of propagation is called the lytic cycle.
Alternatively, the phage genome may enter a dormant stage (prophage) by integrating itself into a bacterial genome by site-specific recombination; during this stage it is propagated along with the host in the sub>sequent progeny. This stage is termed lysogeny. Changes in environmental and physiological conditions may activate the prophage stage and trigger lytic events.
Phage Lambda as a vector
Figure 6. Bacteriophage

The large genome size and complex genetic organization of lambda had posed initial problems with its use as a vector. The problems, however, were surmounted through the sustained efforts of researchers, and lambda has been developed into an efficient vector.
The broad objectives in constructing various phage vectors are
the presence of cloning sites only in the dispensable fragments,
the capacity to accommodate foreign DNA fragments of various sizes,
the presence of multiple cloning sites,
an indication of incorporation of DNA fragments by a change in the plaque type,
the ability to control transcription of a cloned fragment from promoters on the vector,
the possibility of growing vectors and clones to high yield,
easy and ready recovery of cloned DNA,
introduction of features contributing to better biological containment.
There are several difficulties in the use of lambda as a vector.
Some of the problems and the general strategies adopted to overcome them are discussed in this section. Manipulation of Restriction Sites The major obstacle to the use of phage lambda as a cloning vector was essentially the presence of multiple recognition sites for a number of restriction enzymes in its genome.
Initially, all attempts were directed toward minimizing the number of EcoRI sites. Murray and Murray in 1974 were able to construct derivatives of lambda with only one or two EcoRI sites. Similarly, Rambach and Toillais constructed lambda derivatives with EcoRI sites only in the nonessential region of the genome by repeated transfer on restrictive and nonrestrictive hosts . After several cycles of digestion, packaging, and growth, phage derivatives with desirable restriction sites and full retention of infectivity were obtained. All but one HindIII sites were removed by recombination of known deletion mutants or sub>stitutions. Recently, oligonucleotides with specific sequences have been synthesized and introduced into the bacteriophage lambda genome. This has provided a variety of cloning sites in the genome [5].
7.1 Size Limitation for Packaging
The second problem was the requirement of a minimum and maximum genome length (38 and 53 kbp, respectively) for the efficient packaging and for the production of viable phage particles. The viability of the bacteriophage decreases when its genome length is greater than 105% or less than 78% of that of wild-type lambda. Genetic studies of specialized transducing bacteriophages showed, however, that the central one-third of the genome, i.e., the region between the J and Ngenes, is not essential for lytic growth. The presence of a nonessential middle fragment of the phage genome was also revealed during construction of viable deletion mutants. These mutants lack most of the two central EcoRI B fragments which are not essential for lytic growth. However, too much DNA cannot be deleted because there is a minimum 38-kbp requirement essential for efficient packaging. The de novo insertion of DNA (even if heterogeneous) is essential for the formation of viable phages. This constitutes a positive selection for recombinant phages carrying insertions. This approach was successfully exploited in constructing recombinant phages carrying E. coli and Drosophila melanogaster DNA [8].
7.2 Transfection of Recombinant Molecules
The problem of transfection of recombinant molecules constructed in vitro was overcome by the successful in vitro assembly of viable and infectious phage particles. Two types of in vitro packaging systems have been developed so far, i.e., two-strain packaging and single-strain packaging.
Two-strain packaging.
The basis of the two-strain in vitrop ackaging system is the complementation of two amber mutations. Two lambda lysogens, each carrying a single amber mutation in a distinctly different gene, are induced and grown separately so that they can synthesize the necessary proteins. Neither of the lysogens alone is capable of packaging the phage DNA. The role of various phage products in DNA packaging has been studied in detail[3]. The E protein is the major component of the bacteriophage head, and in its absence all the viral capsid components accumulate. The D protein is involved in the coupled process of insertion of bacteriophage DNA into the prehead precursor and the sub>sequent maturation of the head. The A protein is required for the cleavage of the concatenated precursor DNA at the cos sites. Two phage lysogens carrying A and E or D and E mutations in the phage genome are induced separately, and cell extracts are prepared. Neither of the extracts can produce infectious phage particles. However, when the extracts are mixed, mature phage particles are produced by complementation.
The major drawback of the two-strain system is the competition of native phage DNA with recombinant molecules. In both the cell extracts, native phage DNA is also present and can be packaged with an efficiency equal to that of the chimeric DNA. This reduces the proportion of recombinants obtained in a library. The problem of regeneration of endogenous phages obtained in the library was partially overcome by the use of b2-deleted prophages, which poorly excise out of the host chromosome or by UV irradiation of packaging extracts.
Single-strain packaging.
Rosenberg have successfully developed a single-strain packaging system by introducing deletion in the cos region of prophage, rendering the prophage DNA unpackagable because cos is the packaging origin. Induction of the lysogen results in the intracellular accumulation of all protein components needed for packaging.
However, packaging of phage DNA is prevented by the lack of cos sites on the prophage DNA. On the other hand, exogenous DNA with cos sites is packaged efficiently to produce an infectious bacteriophage particle. The single-strain system is superior to two-strain system in having a lower background of parental phages. In addition, it uses E. coli C, which lacks the EcoK restriction system, as the host for the lysogen.
7.3 Biological Containment
The biological containment of recombinant phages is an important aspect from the point of view of ethics and eventual biohazards. It is desirable that cloning vectors and recombinants have poor survival in the natural environment and require special laboratory conditions for their replication and survival. According to Blattner, the lytic phages offer a natural advantage in this respect since the phage and the sensitive bacteria coexist only briefly. A newly inserted segment may not be compatible with E. coli metabolism for extended periods. To make the phage vectors more safe, three amber mutations were introduced in its genome. The new vector Xgt WES XC is safer because an amber suppressor host strain is a very rare occurrence in the natural environment. Many vectors carry one of the amber mutations on the genome so that they can be propagated only on an appropriate suppressor host.
Phage vectors
Many phage vectors have been constructed in the recent past, each with its own special features. There is no universal lambda vector which can fulfill all the desired objectives of the cloning experiments.
The choice of a vector depends mainly on
the size of a DNA fragment to be inserted,
the restriction enzymes to be used,
the necessity for expression of the cloned fragment,
the method of screening to be used to select the desired clones.
Bacteriophage lambda vectors can be broadly classified into two types:
replacement vectors ,
insertion vectors.
Figure 7. Lambda Phage genome

8.1 Replacement Vectors
Taking advantage of the maximum and minimum genome size essential for efficient packaging and the presence of the nonessential central fragment, it is possible to remove the stuffer fragment and replace it with a foreign DNA fragment in the desired size range. This forms the basis of lambdaderived replacement vectors.
Cloning of a foreign DNA in these vectors involves
preparation of left and right arms by physical elimination of the nonessential region,
ligation of the foreign DNA fragment between the arms,
in vitro packaging and infection.
The replacement vectors contain a pair of restriction sites to excise the central stuffer fragments, which can be replaced by a desired DNA sequence with compatible ends. The presence of identical sites within the stuffer fragment but not in the arms facilitates the separation of the arms and the stuffer on density gradient centrifugation. In many vectors, sets of such sites are provided on attached polylinkers so that an insert can be easily excised. Two purified arms cannot be packaged despite their being ligated to each other, because they fall short of the minimum length required for packaging. This provides positive selection of recombinants. The replacement vectors are convenient for cloning of large (in some cases up to 24 kbp) DNA fragments and are useful in the construction of genomic libraries of higher eukaryotes. Charon and EMBL are among the popular replacement vectors.
8.2 Insertion Vectors
Because the maximum packagable size of lambda genome is 53 kb, small DNA fragments can be introduced without removal of the nonessential (stuffer) fragment. These vectors are therefore called insertion vectors. Cloning of foreign DNA in these vectors exploits the insertional inactivation of the biological function, which differentiates recombinants from nonrecombinants. Insertion vectors are particularly useful in cloning of small DNA fragments such as cDNA.
AgtlO and Agtll are examples of this type of vector. In recent years a multitude of lambda vectors have been constructed. Many innovative approaches have been used to introduce desired properties into the vectors. The following section deals with the strategies adopted for the construction of some of the commonly used vectors and their salient features, utilities, and limitations.
8.3 Storage of Lambda Stocks
Most of the lambda strains are stable for several years when stored at 4°C in SM buffer containing 0.3% freshly distilled chloroform (94). The master stocks of bacteriophage lambda are kept in 0.7% (vol/vol) dimethyl sulfoxide at -70°C for long-term storage. Klinman and Cohen have developed a method for storage of a phage library at -70°C by using top agar containing 30% glycerol.
Conclusion
In my work I determined investigations in Molecular cloning, familiarized with vectors for molecular cloning, summarized the received information and made consequences of scientists researches, defined the main tasks of molecular cloning, and made such conclusions:
sequences that permit the propagation of itself in bacteria (or in yeast for YACs) .
a cloning site to insert foreign DNA; the most versatile vectors contain a site that can be cut by many restriction enzymes .
a method of selecting for bacteria (or yeast for YACs) containing a vector with foreign DNA; uually accomplished by selectable markers for drug resistance .
Cloning vector - a DNA molecule that carries foreign DNA into a host cell, replicates inside a bacterial (or yeast) cell and produces many copies of itself and the foreign DNA .
General Steps of Cloning with Any Vector :
prepare the vector and DNA to be cloned by digestion with restriction enzymes to generate complementary ends ;
ligate the foreign DNA into the vector with the enzyme DNA ligase;
introduce the DNA into bacterial cells (or yeast cells for YACs) by transformation ;
select cells containing foreign DNA by screening for selectable markers (usually drug resistance);
Literature
Finbar Hayes The Function and Organization of Plasmids// E. coli Plasmid Vectors Methods and Applications.- 2007.- vol.235 – pp. 1-18.
Mallory J. A. White and Wade A. Nichols Cosmid Packaging and Infection of E. coli// E. coli Plasmid Vectors Methods and Applications.- 2007.- vol.235 – pp. 67-70
Tim S. Poulsen and Hans E. Johnsen BAC End Sequencing // Bacterial Artificial Chromosomes Volume 1: Library Construction, Physical Mapping, and Sequencing.- 2007. – vol.255 - pp.157-162.
Andrew Preston Choosing a Cloning Vector// E. coli Plasmid Vectors Methods and Applications.- 2007.- vol.235 – pp. 19-22.
Sambrook, J., Fritsch, E. F., and Maniatis, T. (eds.) (1989) Bacteriophage λvectors, in Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, pp. 2.3–2.125.
Srividya Swaminathan and Shyam K. Sharan Bacterial Artificial Chromosome Engineering// Bacterial Artificial Chromosomes Volume 2 :Functional Studies.- 2007. – vol.256 – pp. 089-106
www.Microbiologybytes.com
www.wikigenes.org
http:// plasmid.hms.harvard.edu
www. Bookrags.com/YAC