Методы теоретической популяционной генетики
Конец формы
Конец формы
Общие модели эволюции. Методы теоретической популяционной генетики. Теория нейтральности М.Кимуры
1. Классическая популяционная генетика
В этой лекции мы рассмотрим модели, характеризующие общие свойства эволюции. Начнем с синтетической теории эволюции. Эта теория была развита в начале 20-го века. Она основана на учении Ч.Дарвина о естественном отборе и на представлениях Г.Менделя о генах - дискретных элементах передачи наследственных признаков. Большую роль в становлении синтетической теории эволюции сыграла маленькая плодовая мушка Drosophila. Именно эксперименты на этой мушке позволили примирить кажущиеся противоречия между Дарвиновским представлением о постепенном накоплении полезных изменений и наследовании этих изменений и дискретным характером Менделевской генетики. Эксперименты на дрозофиле показали, что мутационные изменения могут быть очень небольшими.
Математические модели синтетической теории эволюции были разработаны Р. Фишером, Дж. Холдейном и С. Райтом. В основном эта математическая теория классической популяционной генетики была завершена к началу 30-х годов.
Согласно синтетической теории эволюции, основным механизмом прогрессивной эволюции является отбор организмов, которые получают выгодные мутации.
2. Математические методы популяционной генетики
Математические модели популяционной генетики количественно характеризуют динамику распределения частот генов в эволюционирующей популяции [1-4,6,8]. Есть два основных типа моделей: 1) детерминистические модели и 2) стохастические модели.
Детерминистические модели предполагают, что численность популяции бесконечно велика, в этом случае флуктуациями в распределении частот генов можно пренебречь, и динамику популяции можно описать в терминах средних частот генов.
Стохастические модели описывают вероятностные процессы в популяциях конечной численности.
Здесь мы кратко охарактеризуем основные уравнения и математические методы популяционной генетики. Наше изложение будет основываться на рассмотрении наиболее характерных примеров. Уравнения моделей мы будем приводить в основном в демонстрационных целях – без вывода, с пояснением смысла этих уравнений; тем не менее, мы будем приводить ссылки на литературу, в которой сделаны соответствующие математические выводы.
2.1. Детерминистические модели
Рассмотрим популяцию диплоидных1) организмов, которые могут иметь несколько аллелей2) A>1> , A>2 >,..., A>K >в некотором локусе3). Мы предполагаем, что приспособленности организмов определяются в основном рассматриваемым локусом. Обозначая число организмов и приспособленность генной пары A>i> A>j >через n>ij> и W>ij> , соответственно, мы можем определить частоты генотипа и гена P>ij> и P>i> , а также средние приспособленности генов W>i> в соответствии с выражениями [1,2,4]:
P>ij> = n>ij >/n , P>i >= S >j >P>ij> , и W>i> =P>i>-1 S >j >W>ij >P>ij> , (1)
где n – численность популяции, индекс i относится к классу организмов {A>i> A>j>} , j = 1,2,..., K> >, которые содержат ген A>i> . Популяция предполагается панмиктической4) : при скрещивании новые комбинации генов выбираются случайным образом из всей популяции.
Для панмиктической популяции приближенно справедлив принцип Харди-Вайнберга [1]:
P>ij> =P>i> P>j> , i, j = 1,..., K. (2)
Уравнение (2) означает, что во время скрещивания генотипы формируются пропорционально частотам генов.
Эволюционная динамика популяции в терминах частот генов P>i >может быть описана следующими дифференциальными уравнениями [1,2,4]:
dP>i> /dt = W>i> P>i> - <W> P>i> - S >j >u>ji >P>i> + S >j >u>ij >P>j> , i = 1,..., K, (3)
где t – время, <W> = S >ij >W>ij >P>ij >– средняя приспособленность в популяции; u>ij> – параметры, характеризующие интенсивности мутационных переходов A>j >--> A>i> , u>ii> =0 (i, j = 1,..., K). Первое слагаемое в правой части уравнения (3) характеризует отбор организмов в соответствии с их приспособленностями, второе слагаемое учитывает условие S >i >P>i> = 1, третье и четвертое слагаемые описывают мутационные переходы.
Отметим, что подобные уравнения используются в модели квазивидов [5], см Лекция 2
Пренебрегая мутациями, мы можем анализировать динамику генов в популяции посредством уравнений:
dP>i> /dt = W>i> P>i> - <W> P>i> , i = 1,..., K. (4)
Используя (1), (2), (4), можно получить (при условии, что величины W>ij >постоянны), что
скорость роста средней приспособленности пропорциональна дисперсии приспособленности V = S >i> P>i> (> >W>i> - <W>)2 [1,3]:
d<W>/dt = 2 S >i> P>i> (> >W>i> - <W>)2. (5)
Таким образом, средняя приспособленность – неубывающая величина. В соответствии с (4), (5), величина L = W>max> - <W> есть функция Ляпунова для рассматриваемой динамической системы (W>max> – локальный или глобальный максимум приспособленности, в окрестности которого рассматривается динамика популяции) [3]. Это означает, что величина L всегда уменьшается до тех пор, пока не будет достигнуто равновесное состояние (dP>i> /dt = 0).
Уравнение (5) характеризует фундаментальную теорему естественного отбора (Р.Фишер,1930), которая в нашем случае может быть сформулирована следующим образом [3]:
"В достаточно большой панмиктической популяции, наследование в которой определяется одним n-аллельным геном, а давление отбора, задаваемое W>ij >, постоянно, средняя приспособленность популяции возрастает, достигая стационарного значения в одном из состояний генетического равновесия. Скорость изменения средней приспособленности пропорциональна аддитивной генной дисперсии и обращается в нуль при достижении генетического равновесия."
Описанная модель – простой пример модели, использующей детерминистический подход. В рамках этого подхода был разработан широкий спектр аналогичных моделей, которые описывают различные особенности динамики генных распределений, например, учитывают несколько генных локусов, возраст особей, число мужских и женских особей, пространственную миграцию особей, подразделение популяции на субпопуляции и т.п. Многие из моделей и расчетов были предназначены для интерпретации конкретных генетических экспериментальных данных [1,3,4] .
2.2. Стохастические модели
Детерминистические модели позволяют эффективно описывать динамику распределения генов в эволюционирующих популяциях. Однако эти модели основаны на предположении бесконечного размера популяции, которое является слишком сильным для многих реальных случаев. Чтобы преодолеть это ограничение, были разработаны вероятностные методы теоретической популяционной генетики [1,3,4,6-8]. Эти методы включают анализ с помощью цепей Маркова (в частности, метод производящих функций) [4,7], и диффузионные [1,3,4,6,8] методы.
Ниже мы кратко рассмотрим основные уравнения и характерные примеры применения диффузионного метода. Этот метод достаточно нетривиален и его применение приводит к достаточно содержательным результатам.
2.2.1. Прямое и обратное уравнения Колмогорова
Рассмотрим популяцию диплоидных организмов с двумя аллелями A>1> и A>2 >в некотором локусе. Численность популяции n предполагается конечной, но достаточно большой, так что частоты гена могут быть описаны непрерывными величинами. Мы также предполагаем, что численность популяции n постоянна.
Введем функцию j = j (X,t|P,0) , которая характеризует плотность вероятности того, что частота гена A>1> равна X в момент времени t при условии, что начальная частота (в момент t = 0) была равна P. В предположении малого изменения частот генов за одно поколение, динамика популяции может быть описана приближенно следующими дифференциальными уравнениями в частных производных [1,3,4,8]:
¶ j /¶ t = - ¶ (M>d> >X >j )/¶ X + (1/2)¶ 2(V>dX >j> >)/¶ X 2 , (6)
¶ j/¶ t = M>d P> ¶ j> >/¶ P + (1/2)V>d> >P >¶ 2j/¶ P 2 , (7)
где M>d> >X >, M>d P >и V>dX >, V>d> >P >– средние значения и дисперсии изменения частот X, P за одно поколение, соответственно; единица времени равна длительности одного поколения. Уравнение (6) есть прямое уравнение Колмогорова. (В физике это уравнение называют уравнением Фоккера-Планка), уравнение (7) – обратное уравнение Колмогорова.
Первые слагаемые справа в уравнениях (6), (7) описывают давление отбора, которое обусловлено разностью приспособленностей генов A>1> и A>2>. Вторые слагаемые характеризуют случайный дрейф частот, который обусловлен флуктуациями в популяции конечной численности.
Используя уравнение (6), можно определять динамику частот генов во времени. Уравнение (7) позволяет оценивать вероятности фиксации генов.
Предполагая, что 1) приспособленности генов A>1> и A>2> равны 1 и 1 - s , соответственно и 2) вклады генов в приспособленности генных пар A>1> A>1>, A>1> A>2> и A>2> A>2> аддитивны, можно получить, что величины M>d> >X >, M>d P >и V>dX >, V>d> >P> определяются следующими выражениями [1,3,4,8]:
M>d> >X >= sX(1-X), M>d> >P> = sP(1-P), V>d> >X> = X(1-X)/(2n), V>d> >P> = P(1-P)/(2n) . (8)
2.2.2. Случай чисто нейтральной эволюции
Если эволюция чисто нейтральная (s = 0), то уравнение (6) принимает вид:
¶ j/¶ t = (1/4n)¶ 2[X(1-X)j]/¶ X 2 . (9)
Это уравнение было решено аналитически М. Кимурой [1,6]. Само решение имеет сложный вид, основные результаты этого решения сводятся к следующему: 1) в конечной популяции фиксируется только один ген (A>1>> >либо A>2>); 2) типичное время перехода от начального распределения к конечному составляет величину порядка 2n поколений. Отметим, что этот результат согласуется с оценками лекции 4 , где была рассмотрена несколько иная модель "чисто нейтральной" эволюции.
2.2.3. Вероятность фиксации гена
Используя уравнение (7), мы можем оценить вероятность фиксации гена A>1 >в конечной популяции. Действительно, рассматривая асимптотику при времени, стремящемся к бесконечности ( t --> inf ), мы можем положить ¶ j /¶ t = 0 и X = 1 ; тогда аппроксимируя вероятность u(P) , которую нужно найти, величиной u(P) = j (1, inf |P,0)/(2n) (здесь u(P) = j(1, inf |P,0)DX , где DX = 1/2n – минимальный шаг изменения частоты в популяции, см. также [3] для более строгого рассмотрения) и комбинируя (7), (8), мы получаем:
s> >du /dP + (1/4n) d 2u /dP 2 = 0 . (10)
Решая это простое уравнение при естественных граничных условиях: u (1) = 1, u (0) = 0 , мы получим вероятность фиксации гена A>1> в конечной популяции [1,3,6]:
u(P) = [1 - exp (- 4nsP)] [1 - exp (- 4ns)]-1 . (11)
Выражение (11) показывает, что если 4ns < < 1 , то имеет место нейтральная фиксация гена: u(P) » P , если 4ns > > 1, то отбирается предпочтительный ген A>1>> >: u(P) » 1; размер популяции n>c> ~ (4s)-1 есть граничное значение, разделяющее области "нейтрального" и "селективного" отбора.
Итак, математические методы популяционной генетики описывают динамику частот генов в эволюционирующих популяциях. Детерминистические методы используются при описании динамики частот в среднем; стохастические методы учитывают флуктуации в популяциях конечной численности.
3. Молекулярная эволюция: теория нейтральности
Классическая теория популяционной генетики, содержательно основанная на синтетической концепции эволюции, интенсивно развивалась до 1960-х годов, до тех пор, пока не возникли трудности интерпретации экспериментальных данных молекулярной биологии. В лекции 1 я уже отмечал, в 1950-1960-х годах произошла революция в молекулярной биологии. Была определена структура ДНК, расшифрован генетический код, ученые установили общие принципы работы молекулярно-генетической системы живой клетки.
Интенсивные исследования молекулярной биологии привели к серьезным результатам, касающимся биологической эволюции: была оценена скорость аминокислотных замен в белках, а также получены оценки, характеризующие полиморфизм белков.
Анализируя экспериментальные данные, М.Кимура обнаружил, что когда он пытался объяснить эти эксперименты на основе селекции благоприятных мутаций путем Дарвиновского отбора, то возникли серьезные затруднения. В своей книге [6] Кимура подробно описывает идеи, послужившие основанием для изобретения теории нейтральности. Например, в некоторых своих оценках, основанных на Дарвинском отборе, он получил, что для объяснения экспериментальных данных нужно потребовать, чтобы каждая особь в процессе эволюции давала 22 000 потомков. И для того, чтобы проинтерпретировать данные по молекулярной эволюции белков, Кимура предложил теорию нейтральности [6,9].
Основное предположение этой теории состоит в следующем: на молекулярном уровне мутации (замены аминокислот или нуклеотидов) преимущественно нейтральны или слабо вредны (существенно вредные мутации также возможны, но они элиминируются из популяции селекцией). Это предположение согласуется с экспериментально наблюдаемой скоростью аминокислотных замен и с тем фактом, что скорость замен в менее важных частях белков значительно больше, чем для активных центров макромолекул.
Используя математические методы популяционной генетики, Кимура получил ряд следствий теории, которые находятся в довольно хорошем согласии с данными молекулярной генетики [6].
Математические модели теории нейтральности существенно стохастические, т.е. относительно малая численность популяции играет важную роль в фиксации нейтральных мутаций. См. примеры расчетов, приведенных выше.
Но если молекулярные замены преимущественно нейтральны, как возможна прогрессивная эволюция? Чтобы ответить на этот вопрос, Кимура использует концепцию дупликации генов, развитую С.Оно [10]. Согласно теории Кимуры, дупликация генных участков создает дополнительные, избыточные ДНК-последовательности, которые в свою очередь дрейфуют далее за счет случайных мутаций, предоставляя тем самым сырой материал, из которого могут возникать новые, биологически значимые гены (Рис.1).
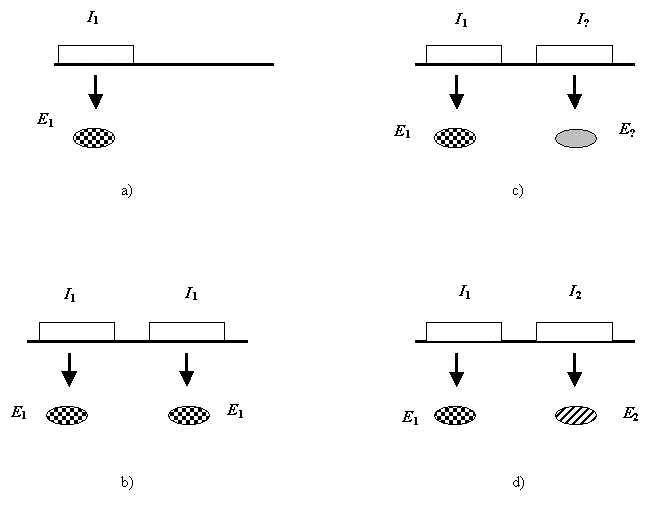
Рис. 1. Иллюстрация к механизму прогрессивной эволюции в теории нейтральности. Схема появления нового биологически значимого белка. Показаны участки ДНК ( I>i> ) и кодируемые ими белки ( E>i> ). a) ген I>1>> >кодирует белок E>1>, b) дупликация гена I>1>, новый участок (справа) кодирует тот же белок E>1>, c) случайный дрейф правого участка, d) возникновение нового биологически значимого белка E>2> кодируемого участком ДНК I>2>> >.
Заключая наш сжатый обзор теории нейтральности, процитируем пять принципов этой теории [6]. Первые четыре из них – эмпирические, а пятый установлен теоретическим путем.
Скорость эволюции любого белка, выраженная через число аминокислотных замен на сайт в год, приблизительно постоянна и одинакова в разных филогенетических линиях, если только функция и третичная структура этого белка остаются в основном неизменными.
Функционально менее важные молекулы и их части эволюционируют (накапливая мутационные замены) быстрее, чем более важные.
Мутационные замены, приводящие к меньшим нарушениям структуры и функции молекулы (консервативные замены), в ходе эволюции происходят чаще тех, которые вызывают более существенное нарушение структуры и функции этой молекулы.
Появлению нового в функциональном отношении гена всегда должна предшествовать дупликация гена.
Селективная элиминация вредных мутаций и случайная фиксация селективно нейтральных или очень слабо вредных мутаций происходят в ходе эволюции гораздо чаще, чем положительный дарвиновский отбор благоприятных мутаций.
4. Другие модели, характеризующие общие закономерности эволюции
Теория нейтральности – одна из наиболее разработанных общих теорий эволюции. Однако есть ряд моделей и концепций, также характеризующих эволюцию на молекулярном уровне, которые в основном дополняют теорию нейтральности. Отметим наиболее известные из них.
В работах Д.С.Чернавского и Н.М.Чернавской [11,12] сделана оценка вероятности случайного формирования нового биологически значимого белка с учетом того, что в белке есть активный центр, в котором замены аминокислот практически недопустимы, и участки, свойства которых не сильно меняются при многих аминокислотных заменах. Там же сделана оценка количества возникающей в геноме информации при появлении нового белка. Полученная оценка указывает на то, что случайное формирование белка было вполне вероятно в процессе эволюции.
Интересна, хотя, по-видимому, не бесспорна, модель блочно-иерархического эволюционного отбора [13,14], согласно которой новые генетические тексты большой длины сначала случайно составляются из коротких текстов, оптимизированных в предыдущие эволюционные эпохи, а после составления оптимизируются. Модель блочно-иерархического эволюционного отбора критически проанализирована в [15].
Блочно-модульный принцип организации и эволюции молекулярно-генетических систем управления обосновывается В.А.Ратнером [16]. Согласно этому принципу эволюция генов, РНК, белков, геномов и молекулярных систем управления на их основе шла путем комбинирования блоков (модулей) снизу доверху, причем модулями, из которых составлялись вновь возникающие молекулярно-генетические системы, служили уже функционирующие макромолекулярные компоненты. По сравнению с моделью блочно-иерархического отбора блочно-модульный принцип более гибок и более реалистичен.
В модели "генов-мутаторов" [17] предполагается, что уровень мутаций может меняться и наследоваться, в результате чего при попадании популяции в новую среду, когда выгоден активный поиск новых свойств, уровень мутаций возрастает, а при длительном нахождении в постоянной среде, где важно сохранение уже найденных свойств, интенсивность мутаций падает.
Интересно проанализировать, как могли возникать достаточно нетривиальные системы обработки информации. Для простейших организмов (вирусов и бактерий) в качестве таковых можно рассматривать регулирование синтеза белков (функциональных и структурных элементов организма) в соответствии с определенной "программой". Например, в процессе онтогенеза фага Т4 происходит образование сложной пространственной структуры, в формировании которой участвуют несколько десятков белков, синтезируемых в соответствии с программой, закодированной геномом фага [18]. Иллюстративная модель эволюционного возникновения подобных "программ жизнедеятельности" предложена [19]. Согласно модели в процессе эволюционного формирования этих программ в генотип закладывается некоторая избыточность, которая приводит к тому, что при небольшой модификации генома часть блоков программ сохраняется. При введении в модель представлений о "генах-мутаторах" наблюдалось поведение, качественно сходное с явлением каскадного мутагенеза [20] – резким возрастанием интенсивности мутаций после дестабилизации генома.
В чрезвычайно интересном цикле работ С.Кауффмана с сотрудниками [21,22] исследуется эволюция автоматов, состоящих из соединенных между собой логических элементов. Отдельный автомат можно рассматривать как модель молекулярно-генетической системы управления живой клетки, при этом каждый логический элемент интерпретируется как регулятор синтеза определенного фермента. Модели Кауффмана позволяют сделать ряд предсказаний относительно "программ" жизнедеятельности клеток. В частности, продемонстрировано, что для одновременного обеспечения устойчивости и гибкости программы число входов логических элементов должно быть ограничено определенным интервалом, а именно составлять величину примерно равную 2-3. Для моделей Кауффмана разработаны эффективные методы анализа на базе статистической физики, эти модели получили широкую известность и исследовались рядом ученых. Подробнее основные результаты этой модели мы обсудим в следующей лекции.
Специальные термины:
1) Диплоидный организм: особь, имеющая два набора хромосом в каждой из ее клеток.
2) Аллель: Одна из различных форм гена, который может быть в заданном локусе.
3) Локус: участок хромосомы, в котором локализован ген.
4) Панмиксия: полностью случайное скрещивание.
Литература
J.F. Crow, M. Kimura. "An introduction to population genetics theory". New York etc, Harper & Row. 1970.
T. Nagylaki. "Introduction to theoretical population genetics ". Berlin etc, Springer Verlag. 1992.
Свирежев Ю.М., Пасеков В.П. Основы математической генетики. М.: Наука, 1982, 511 с.
P.A.P. Moran. “The statistical processes of evolutionary theory”, Oxford, Clarendon Press, 1962. Имеется перевод: П. Моран. Статистические процессы эволюционной теории. М.: Наука, 1973. 288 с.
Эйген М., Шустер П. Гиперцикл. Принципы самоорганизации макромолекул. М.: Мир, 1982. 270 с.
Кимура М. Молекулярная эволюция: теория нейтральности. М.: Мир, 1985, 400 с.
S. Karlin. "A first course in stochastic processes". Academic Press. New York, London, 1968. Имеется перевод: С.Карлин. Основы теории случайных процессов. М.: Мир, 1975.
Ратнер В.А. Математическая популяционная генетика. Новосибирск: Наука, 1976. 128 с.
M. Kimura. Evolutionary rate at the molecular level // Nature. London, 1968.V.217. PP.624-626.
Оно С. Генетические механизмы прогрессивной эволюции. М.: Мир, 1973, 228 с
Чернавская Н.М., Чернавский Д.С. Проблема возникновения новой информации в эволюции // Термодинамика и регуляция биологических процессов. Теория информации, управление в живых системах, проблема самоорганизации, эволюция и онтогенез. М.: Наука. 1984. С.247-255
Романовский Ю.М., Степанова Н.В., Чернавский Д.С. Математическая биофизика. М.: Наука, 1984. 304 с.
Иваницкий Г.Р., Есипова Н.Г., Абагян Р.А., Шноль С.Э. Блочное совершенствование генетического текста как фактор ускорения биологической эволюции // Биофизика. 1985. Т.30. N.3. С.418-421.
Шноль С.Э. Хватает ли времени для дарвиновской эволюции? // Природа, 1990. N.11. С. 23-26.
Моносов Я.А. О факторах ускорения биологической эволюции // Биофизика. 1991. Т.36. N.5. С.920-922.
Ратнер В.А. Блочно-модульный принцип организации и эволюции молекулярно-генетических систем управления (МГСУ) // Генетика. 1992. Т.28. N.2. С.5-23.
Семенов М.А., Теркель Д.А. Об эволюции механизмов изменчивости посредством косвенного отбора // Журн. общ. биологии. 1985.Т. 46. N.2. С. 271 -277.
Ратнер В.А. Молекулярно-генетические системы управления. Новосибирск: Наука, 1975. 288 с.
Редько В.Г. К теории эволюции. Модель происхождения "программ жизнедеятельности" // Журн. общ. биологии. 1991.Т.52. N.3. С. 334-342.
Корогодин В.И. Кариотаксоны, надежность генома и прогрессивная эволюция // Природа. 1984. N.2. С.3-14.
Kauffman S.A., Smith R.G. Adaptive automata based on Darwinian selection // Physica D. 1986. V.22. N.1-3. P.68-82.
Кауффман С. Антихаос и приспособление // В мире науки. 1991. № 10. С. 58.
Copyright © Vladimir Red'ko, Oct 25, 1999 ( redko@keldysh.ru )
Last modified: Oct 25, 1999