Биологическая сущность микоризы
Федеральное агентство по образованию
Государственное образовательное учреждение высшего профессионального образования
УЛЬЯНОВСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
Институт медицины, экологии и физической культуры
Экологический факультет
Кафедра биоэкологии и генетики человека
КУРСОВАЯ РАБОТА
"МИКОРИЗА"
Студентка Левщанова Юлия Сергеевна,
1 курс, специальность биоэкология
Научный руководитель, д.б.н.,
профессор И.В. Благовещенский
Ульяновск, 2008
СОДЕРЖАНИЕ
Введение
1. Микориза
2. Генно-инженерные эксперименты арбускулярной микоризы
3. Грибы-симбионты
4. Типы микоризы
5. Биологическая сущность микоризы
Заключение
Список использованных источников
Введение
Симбиоз корней растения с грибами сложился исторически. Отпечатки грибных нитей ученые обнаружили у ископаемых растений, давно вымерших на земном шаре (например, у псилофитов). Остатки древней растительности, еще сохранившиеся в современной флоре (плауны или виды Psilotum) , требуют наличие микоризы. Голосеменные растения, куда входят наши хвойные породы, саговники, гинкго, представленные древесным типом, являются микоризными растениями. Много древесных покрытосеменных (цветковых) также сожительствуют с почвенными грибами. Это сожительство развилось, по-видимому, на торфянистых и перегнойных почвах, столь характерных для древней растительности. Азот этих почв мог стать доступным корням высших растений в значительной мере благодаря грибам, которые, пронизывая частицы этих почв, производят разложение перегнойных веществ и переводят содержащийся в них азот в доступные формы (аммонийные). В клетках коры корней, несущих внутреннюю микоризу, находят много сахарозы, которая не только усваивается грибом, но и откладывается им в запас. Эти же клетки коры оказываются богатыми фосфором и калием ( Катенин, 1972).
1. Микориза
Микориза - симбиотическое обитание грибов на корнях и в тканях корней высших растений. В микоризе гриб получает от корней углеводы и снабжает растение водой и минеральными элементами питания (рис. 1.).
Микориза (Mycorhiza) - термин, предложенный Франком, для обозначения корней, тесно сросшихся с грибом в один орган - грибо-корень (muchu -гриб, riza - корень). Такие корни встречаются у многих наших деревьев у дуба, бука, граба, орешника, ив, тополей, многих хвойных и др. Молодые корни этих растений сплошь оплетаются тоненькими гифами гриба; по мере нарастания корня нарастает и окутывающий его грибной чехол (Катенин, 1972).

Рис.1. Микориза (Катенин, 1972)
2. Генно-инженерные эксперименты арбускулярной микоризы
Эволюционный успех наземных растений во многом был обеспечен симбиозом с почвенными грибами и бактериями, которые снабжают растение соединениями азота и фосфора, получая взамен питательные вещества, образуемые растением входе фотосинтеза.
Самым древним вариантом такого симбиоза, по-видимому, является микориза, известная в двух основных вариантах: более простая эктомикориза (гриб не проникает внутрь растительных клеток) и эндомикориза, или арбускулярная микориза(АМ) (см. рис. 2.3), при которой гифы гриба врастают внутрь клеток корня. Судя по палеонтологическим данным, АМ существовала уже 450млн лет назад, в ордовикском периоде. О ее древности свидетельствует также ее широкое распространение во всех группах наземных растений. Симбиотические отношения с азотфиксирующими бактериями (к которым относятся ризобии и актинобактерии) тоже, скорее всего, имеют очень древнюю историю, однако в большинстве случаев речь идет о внеклеточных бактериальных симбионтах. Только некоторые покрытосеменные (цветковые) растения сравнительно недавно научились культивировать бактерии внутри клеток своих корней, в особых органах— клубеньках. Клубеньковые симбиозы бывают двух типов: 1)симбиоз бобовых с ризобиями (бактерии из группы альфапротеобактерий) (СБР), 2) актинориза (АР)— симбиоз с актинобактериями рода Frankia. Актинобактерии образуют мицелиальные структуры наподобие грибов; раньше их относили к грибам и называли актиномицетами. Клубеньковые симбиозы встречаются только в четырех группах (порядках) покрытосеменных: у бобовых (Fabales), розовых (Rosales), тыквенных (Cucurbitales) и буковых (Fagales), причем не у всех, а только участи представителей. Недавно на основе молекулярно-генетических данных было установлено, что эти четыре отряда представляют собой монофилетическую кладу, то есть группу, происходящую от единого общего предка. Было высказано предположение, что у общего предка этой группы произошли какие-то генетические изменения, обусловившие принципиальную возможность развития клубенькового симбиоза того или иного типа. Одни представители группы впоследствии воспользовались этой возможностью, другие нет. Симбиоз бобовых с ризобиями (СБР) изучен гораздо лучше, чем актинориза (АР). Удалось выявить связь между СБР и АМ: оказалось, что по меньшей мере семь генов задействованы в обоих симбиозах. Эти гены получили название "общих генов симбиоза" (common symbiosis genes). На этом основании было высказано предположение, что при становлении СБР была использована древняя генетическая программа, сложившаяся изначально для обслуживания внутриклеточного симбиоза с грибами(АМ). Однако какие именно изменения были внесены в эту программу, до сих пор оставалось не ясным.
И вот наконец британским и германским биологам удалось обнаружить одно существенное генетическое различие между "клубеньковыми" и "бесклубеньковыми" цветковыми растениями. Ученые проанализировали строение белков, кодируемых "общими генами симбиоза", у разных групп цветковых растений. Оказалось, что большинство этих белков имеют практически одинаковую ("консервативную") структуру у всех цветковых. И только один из них оказался вариабельным. Белок этот называется SYMRK (symbiosis receptor kinase). В чем именно состоит его функция, экспериментально не установлено, но многое можно сказать на основе анализа его доменной структуры (домен— функциональная часть или блок белковой молекулы, содержащий некий узнаваемый аминокислотный "мотив", по которому обычно можно судить о функции данного домена). У белка SMYRK есть, во-первых, трансмембранный домен, который, как видно из названия, располагается в толще клеточной мембраны. Внутриклеточная часть белка содержит домен протеин-киназы, функция которого состоит в переносе фосфата с АТФ на какой-нибудь белок (таким способом многие рецепторные белки передают полученный сигнал внутрь клетки, поскольку фосфорилирование белков оказывает радикальное влияние на их свойства— например, переводит их в активное состояние из неактивного). И трансмембранный домен, и домен протеин-киназы в этом белке примерно одинаковы у всех исследованных авторами видов цветковых растений. Основные различия сосредоточены во внеклеточной части белка, которая, несомненно, выполняет рецепторную функцию (улавливает какой-то химический сигнал). Исследователи выявили три основных варианта внеклеточной части белка SYMRK:
"длинный вариант", характерный для растений, образующих клубеньки, а также ближайших их родственников;
"средний вариант", характерный для более дальних двудольных родственников "клубеньковых" растений;
"короткий вариант" характерный для однодольных (риса и кукурузы) .
Клубеньки любого типа (СБР или АР) встречаются только у обладателей "длинного" варианта гена SYMRK. Арбускулярная микориза (АМ) встречается у обладателей всех трех вариантов гена (рис. 2.1).

Рис. 2.1. Структура гена SYMRK и типы внутриклеточного корневого симбиоза у цветковых растений. Слева— эволюционное древо цветковых. Серым прямоугольником обозначены четыре порядка, у представителей которых встречаются клубеньковые симбиозы (АР или СБР). В овальные рамки заключены родовые названия растений, у которых анализировалась структура гена. цветом выделены растения, образующие АМ и АР, серым— АМ и СБР, белым— только АМ. Схематичные рисунки изображают три типа внутриклеточного симбиоза: AR=АР, RLS=СБР, AM=АМ. Три длинных горизонтальных прямоугольника отображают структуру гена SYMRK. Буквами обозначены домены: NEC, LRR— предполагаемые рецепторные (внеклеточные) домены; TM— трансмембранный домен; PK— протеин-киназный (внутриклеточный) домен. (Рис. из обсуждаемой статьи вPLoS Biology )
Обнаружив эту закономерность, авторы, естественно, предположили, что приобретение "длинного" варианта SYMRK и было тем самым ключевым событием, которое создало необходимые предпосылки для развития клубеньковых симбиозов с бактериями— причем "генетическая программа" клубенькового симбиоза представляет собой модификацию "генетической программы" арбускулярной микоризы. Чтобы подтвердить или опровергнуть это предположение, ученые провели серию генно-инженерных экспериментов.

Рис. 2.2. Растение Datisca glomerata (Katharina Markmann, 2008)
Первый эксперимент показал, что ген SYMRK необходим не только для АМ и СБР (что было известно и ранее), но и для АР. Ученые отключили ген SYMRK у растения Datisca glomerata, корни которого в норме образуют АМ и АР (рис. 2.2). результате растение практически полностью утратило способность к формированию обоих симбиозов— и с грибом, и с актинобактерией Frankia. Тем самым впервые удалось показать, что ген SYMRK необходим для всех трех типов внутриклеточного симбиоза: АМ, СБР и АР. Стало ясно, что АР имеет, скорее всего, примерно ту же генетическую "основу", что и СБР ( ранее о генетики АР не было известно практически ничего).
Второй эксперимент показал, что ген SYMRK не служит для распознавания конкретных бактерий-симбионтов. Бобовое растение лядвенец японский (Lotus japonicus) образует СБР с бактерией Mesorhizobium loti, алюцерна (Medicago truncatula)— с бактерией Sinorhizobium melioti. Мутантной люцерне, имеющей "испорченный" ген SYMRK и не способной формировать ни АМ, ни СБР, пересадили "здоровый" ген SYMRK от лядвенца. Эта операция полностью восстановила способность люцерны образовывать оба типа симбиоза. При этом трансгенная люцерна стала образовывать СБР со "своей" исконной бактерией Sinorhizobium, а вовсе не с Mesorhizobium. Таким образом, SYMRK не отвечает за распознавание и выбор бактериального симбионта, а только за общую способность формировать внутриклеточный симбиоз с бактериями. Распознавание осуществляется другими белками, какими именно— пока не установлено.
Третий эксперимент показал, что для обеспечения нормального клубенькового симбиоза, равно как и микоризы(АМ), вполне подходит любой "длинный" вариант гена SYMRK, взятый хоть у бобового растения, образующего СБР, хоть у растения с АР, хоть у вида, вовсе не образующего клубеньков. Для эксперимента были использованы мутантные растения Lotus japonicus, у которых ген SYMRK кодирует нефункциональный белок с испорченным киназным доменом. Эти растения не могут образовывать ни СБР, ни АМ. Им пересаживали гены SYMRK от разных растений, имеющих длинный вариант этого гена: от других бобовых, образующих СБР (люцерны), от актиноризных растений (Datisca glomerata) и, наконец, от бесклубеньковых родственников (настурции Tropaeolum majus). Все эти операции привели к полному восстановлению у мутантного лядвенца АМ и СБР (разумеется, со"своей" бактерией Mesorhizobium).
Четвертый эксперимент показал, что укороченные варианты гена SYMRK достаточны для АМ, но не для клубеньковых симбиозов. Как и в третьем эксперименте, использовали мутантную форму лядвенца японского, не образующую ни АМ, ни СБР. Растениям пересаживали "средний" вариант гена, взятый у помидора, и "короткий" вариант, позаимствованный у риса. В обоих случаях у мутантного лядвенца восстановилась способность к формированию АМ, но не СБР. На основе этих и ряда других экспериментов и наблюдений ученые заключили, что белок SYMRK, по-видимому, необходим для формирования особых внутриклеточных структур— своеобразных "с имбионтоприемников" или "пре-инфекционных нитей" (pre-infection threads), которые впоследствии заселяются симбиотическими бактериями (и тогда их уже называют "инфекционными нитями"). Похожие "симбионтоприемники" образуются в клетках корней и перед принятием грибных симбионтов при формировании АМ (эти структуры называются pre-penetration apparatus). Сходство в строении и механизмах формирования этих "симбионтоприемников", по-видимому, отражает единство генетической программы, отвечающей за формирование всех трех типов внутриклеточного симбиоза: АМ, СБР и АР. Мутации в некоторых "общих генах симбиоза" приводят к нарушению формирования "симбионтоприемников" (SYMRK, конечно, не единственный ген, необходимый для их формирования). Следует подчеркнуть, что "длинная" версия белка SYMRK, очевидно, является необходимым, но недостаточным условием формирования клубеньковых симбиозов. Это видно из того, что такие симбиозы могут формировать не все, а только некоторые обладатели "длинной" версии. По-видимому, растения, образующие клубеньки, должны обладать еще какими-то генетическими особенностями, которые пока не удалось обнаружить.
В целом, однако, полученные результаты убедительно подтверждают гипотезу, согласно которой способность к формированию клубеньковых симбиозов (АР и СБР) развилась на основе древней генетической программы АМ. Ключевое эволюционное событие заключалось в том, что клетки корней приобрели способность реагировать формированием "симбионтоприемников" не только на присутствие симбиотических грибов, но и на близость азотфиксирующих бактерий. Очень похоже, что в основе этого события лежало изменение структуры белка SYMRK, а именно добавление двух новых рецепторных доменов к его внеклеточной части. Оба эти домена могли быть заимствованы у генов других белков, имеющихся в геноме высших растений. Таким образом, возникновение клубеньковых симбиозов— яркий пример формирования новой функции путем модификации генного комплекса, ранее служившего для иных целей. Арбускулы— видоизменения мицелия у грибов - микоризообразователей, аналогичные гаусториям. Являются многократно дихотомически разветвлёнными гифами сложной формы, проникающими в паренхимальные клетки корня. Окружены внутри клетки клеточной плазмалеммой. В арбускулах происходит наиболее интенсивный обмен метаболитами между компонентами микоризы, хотя они и существуют лишь несколько дней (в последствии растворяются). Исследователями считается, что арбускулы образуются под влиянием защитной реакции клеток растения (Katharina Markmann и др., 2008)
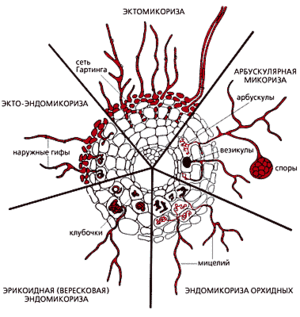
Рис. 2.3. Морфологические особенности различных типов микоризы (
3. Грибы-симбионты
Микоризные грибы (от греч. mykes - гриб, rhiza - корень) образуют симбиоз с высшими растениями. Микориза - очень интересное и уникальное явление в природе: абсолютно разные по строению и способу жизни организмы объединяются, оказывают друг другу помощь в существовании. Есть хорошая пословица "Леса нет - и гриба нет". Это своеобразное сообщество растений и грибов, которое очень часто отражается в названиях последних (подберезовик, подосиновик), нашло научное обоснование в открытии явления микоризы. Это представляется следующим образом: когда мицелий определенного вида изначально распространяется в почве и на каком-то этапе своего развития встречает мелкие корешки растений определенного вида, то оплетает их своеобразной, невидимой глазом "муфтой", образуя грибной чехлик. Это хорошо и для растения, и для гриба. Растение обеспечивает микоризный гриб углеводами, углекислым газом, а при недостатке - кислородом. В свою очередь гриб участвует в обеспечении растения макро- и микроэлементами, водой. Кроме этого, микоризный гриб предохраняет корни растения от вредных микроорганизмов.
Без микоризы мицелий симбиотических грибов может определенное время существовать в почве, но при этом никогда не будет образовывать плодовых тел. Поэтому в искусственных условиях нельзя получить плодовые тела белого гриба, сыроежковых грибов, мухоморов - все они являются микоризообразователями и без определенной древесной породы не будут плодоносить. В свою очередь растение без своих грибных симбионтов развивается плохо, медленно, легко подвергается заболеваниям и даже может погибнуть. Так, например, сеянцы сосны могут вырасти крепкими и высокими только в том случае, если их корни оплетают определенные виды грибов.
В середине 50-х годов XX в. было принято решение вокруг города Элиста (Калмыкия) насадить зеленое кольцо, чтобы предохранить город от сильных степных ветров. Для этой цели выбрали акацию. Посадили раз - все деревца погибли, посадили второй и третий - все тоже. Обратились к ученым за советом. Когда провели анализы почвы и древесных остатков, то установили, что в почве отсутствовали микоризообразователи акации. Уже на следующий год были высажены новые молодые деревца, которые прижились и начали расти, поскольку предварительно в почву был внесен мицелий соответствующих грибов.
Семена орхидей имеют микроскопически маленькие размеры и не содержат никаких питательных веществ, поэтому могут прорасти только при наличии гриба-симбионта. Ростки орхидных длительное время должны питаться через симбионтов, которые разрушают готовые органические вещества. Вследствие этого орхидные изначально обзаводятся целой свитой микоризных грибов, способных давать им возможность осваивать питательные вещества. Через определенное время симбиоз становится им не нужен и гриб погибает, но не раньше, чем успеет образовать большое количество дозревших спор, которые будут разнесены водой или ветром и вновь вступят в симбиотические отношения с новыми ростками орхидных.
Как видно из вышеприведенных примеров, значение микоризных грибов очень большое: они способствуют росту и успешному развитию древесных, кустарниковых и травянистых растений. К микоризным грибам относится большое количество микромицетов, практически все трубчатые, все сыроежковые и около 35 % всех пластинчатых грибов. Поэтому каждый, кто бывает в лесу, не должен уничтожать неизвестные ему и даже ядовитые грибы, поскольку они нужны самому лесу и его жителям ( Мухин и др., 1976).
4. Типы микоризы
Различают Микориза эктотрофную (наружную), при которой гриб оплетает покровную ткань окончаний молодых корней и проникает в межклетники самых наружных слоев коры, и эндотрофную (внутреннюю), которая характеризуется внедрением мицелия (гиф гриба) внутрь клеток. Эктотрофная Микориза характерна для многих деревьев (дуб, ель, сосна, берёза), кустарников (ива), некоторых кустарничков (дриада) и травянистых растений (гречиха живородящая). Молодые корни этих растений обычно ветвятся, окончания их утолщаются, растущая часть корней окутывается толстым плотным грибным чехлом, от которого в почву и по межклетникам в корень на глубину одного или несколько слоев коры отходят гифы гриба, образуя т. н. сеть Гартига; корневые волоски при этом отмирают (эуэктотрофный тип Микориза). У кустарничка арктоуса арктического и травянистого растения грушанки крупноцветковой гифы гриба проникают не только в межклетники, но и в клетки коры (эктоэндотрофный тип Микориза). Эктотрофные Микориза образуют чаще гименомицеты (роды Boletus, Lactarius, Russula, Amanita и др.), реже - гастеромицеты. В образовании Микориза на корнях одного растения может участвовать не один, а несколько видов грибов. Однако, как правило, в растительных сообществах встречаются лишь определённые грибы-микоризообразователи - симбионты данных видов растений.
При развитии эндотрофной Микориза форма корней не меняется, корневые волоски обычно не отмирают, грибной чехол и "сеть Гартига" не образуются; гифы гриба проникают внутрь клеток коровой паренхимы. У растений семейства вересковых, грушанковых, брусничных и шикшевых гифы гриба в клетках образуют клубки, позднее перевариваемые растением (эрикоидный тип Микориза). В образовании Микориза такого типа участвуют фикомицеты (роды Endogone, Pythium). У растений семейства орхидных гифы гриба из почвы проникают в семя, образуя клубки, перевариваемые затем клетками семени. Из грибов такой тип Микориза свойствен несовершенным (род Rhizoctonia) и реже - базидиальным (род Armillaria и др.). Наиболее распространён в природе - у многих однолетних и многолетних трав, кустарников и деревьев самых различных семейств - фикомицетный тип Микориза, при котором гифы гриба пронизывают насквозь клетки эпидермиса корня, локализуясь в межклетниках и клетках средних слоев коровой паренхимы.
Микориза оказывает на растение благоприятное воздействие: за счёт развитого мицелия увеличивается поглощающая поверхность корня и усиливается поступление в растение воды и питательных веществ. Грибы-микоризообразователи, вероятно, способны разлагать некоторые недоступные растению органические соединения почвы, вырабатывают вещества типа витаминов и активаторы роста. Гриб же использует некоторые вещества (возможно, углеводы), извлекаемые им из корня растения. При разведении леса на почве, не содержащей грибов-микоризообразователей, в неё вносят в небольших количествах лесную землю, например при посеве желудей - землю из старой дубравы ( Контримавичус, 1982).
5. Биологическая сущность микоризы
Всходы многих видов лесных деревьев, выращенные в стерильном питательном растворе, а затем перенесенные в луговую почву, будут плохо расти и даже погибать от недостатка пищи, хотя субстрат достаточно богат ею. Однако если добавить к почве вокруг корней сеянцев совсем немного (0,1 % по объёму) лесной почвы, содержащей соответствующие грибы, рост нормализуется. Это обусловлено микоризой ("грибокорнем"), тесным взаимовыгодным симбиозом корней и грибов. Микориза известна в большинстве групп сосудистых растений. Всего несколько семейств цветковых не образуют её или образуют очень редко, например крестоцветные и осоковые. Многие растения могут нормально развиваться и без микоризы при хорошем обеспечении незаменимыми элементами, особенно фосфором. Однако при ограниченной доступности этих минеральных веществ они растут без нее плохо или погибают. Участие микоризы в прямом транспорте фосфора из почвы в корни доказано экспериментально. В свою очередь растения снабжают симбиотические грибы углеводами. Гриб помогает растению усваивать минеральные соли и воду, а сам получает от него органические вещества, которые растения синтезируют из неорганических веществ. Микориза увеличивает способность корней поглощать вещества из почвы в тысячи (!) раз. С грибами вступают в симбиоз 80-98% (по разным источникам) наземных растений: деревья, кустарники, травы. Например, в итоге многолетней работы (Селиванов И.А. "Микосимбиотрофизм как форма консортивных связей в растительном покрове Советского Союза". М., 1981) было изучено 3449 видов (не экземпляров!) растений в разных климатических зонах.78% из них оказались микотрофными (т.е. те, которые могут образовывать микоризу): деревья, травы, кустарники. Иначе говоря, три четверти растительного мира питаются при помощи грибов!Дубы, сосны и многие другие растения без микоризы жить просто не способны. Степи, луга, леса в таком их виде, который для нас привычен, не смогли бы существовать без микоризы. Точно так же без микоризы большинство шляпочных грибов не смогло бы образовывать плодовые тела: на это просто не хватило бы сил. Редкое исключение из правила - уже упомянутые шампиньоны, которые микоризу не образуют. И всё-таки микоризные шляпочные грибы можно вырастить на выбранном месте - но не на грядке, а у подходящего дерева. Большое количество микоризных грибов могут жить не с одной, а со многими древесными породами. Например, подосиновик образует микоризу не только с осиной, но и с березой, а белый гриб сожительствует почти с пятью - десятью деревьями (рис. 5.1)
Макромицеты, образующие микоризу на корнях деревьев и кустарников, составляют 40% от общего количества шляпочных грибов. В почве мицелий гриба плотным чехлом гиф окутывает мелкие корни и корневые волоски растений, во много раз увеличивая площадь их питания.

Рис. 5.1. Подосиновик(Селиванов, 1981)
Многие необходимые растениям элементы, например, некоторые соединения фосфора, кальция, калия, азота, находятся в почве в недоступной для растений форме. И если бы симбиоз грибов и растений не был так широко распространен в природе, растения сильно страдали бы от минерального голода. Велика роль микоризы и в снабжении растений водой. Особенно важна эта функция в условиях недостаточной влажности, т.н. физиологической сухости и засоленности почв – в холодных таежных и сухих полупустынных и пустынных областях микориза становится практически единственной формой почвенного питания древесных растений. Микоризные грибы участвуют и в защите растений от патогенных организмов. Есть данные о том, что гибель чистых культур сосны вызывается отсутствием в почвах микоризных грибов и развитием вследствие этого патогенного гриба Trametes radiciperta. Предоставляя столь многочисленные преимущества растениям, грибы получают от них углеводы, которые из-за отсутствия хлорофилла не способны синтезировать сами.
Большинство наших растений – деревьев, кустарников и трав – образуют микоризу, например, у цветковых растений она не встречается только у семейства крестоцветных и осоковых. Широко распространенная микотрофность древесных пород является основой существования лесов практически во всех зонах умеренного климата. Она облигатна (обязательна) для представителей семейств сосновых, березовых и буковых. Интенсивность формирования микоризы зависит прежде всего от содержания в почве доступных для растений форм азота, фосфора, калия и степени освещенности местообитаний. Максимальное ее развитие наблюдается при низких концентрациях одного из названных элементов. Большинство собираемых нами в лесу съедобных грибов относятся именно к микоризообразователям-симбиотрофам. Это все трубчатые грибы – белые, подосиновики, подберезовики, маслята, моховики, дубовики, польский и желчный грибы; пластинчатые – сыроежки, грузди, рыжики, рядовки, зонтики, а также ядовитые, несъедобные и малоизвестные в пищевом отношении грибы – мухоморы, паутинники. Микоризу образуют некоторые гастеромицеты и аскомицеты: трюфели, строчки, ложнодождевики. Говоря о возможности отнесения того или иного вида гриба к микоризообразователям, следует отметить, что большая их часть либо мобильна в отношении типа питания, либо может образовывать плодовые тела в различных по экологической обстановке условиях. Так, например, обычные в переувлажненных местообитаниях (заболоченные сосняки, березняки, ельники) микоризные грибы – свинушка тонкая, желчный гриб и некоторые другие виды – визуально могут быть отнесены к деревообитающим, так как их плодовые тела встречаются на пнях и кочках. Однако при этом связи между корнями живых деревьев и мицелием грибов не теряются.
Существует немало грибов, способных образовывать плодовые тела как при участии в микоризе, так и без связи с корнями деревьев. В неблагоприятных для деревьев условиях, например, в сфагновых сосняках и березняках, в искусственных лесных насаждениях (лесозащитные полосы в степях, полупустынях) такие грибы, как свинушка тонкая и лаковица, ведут себя как истинные микоризообразователи. А в лесных сообществах с благоприятными для деревьев условиями они меняют тип питания на сапротрофный (питание мертвым органическим веществом растений).
Заключение
Многообразные формы взаимоотношения грибов с растениями обусловили и их использование в биотехнологиях. Сегодня микоризация растений, способствующая повышению продуктивности и устойчивости к патогенным организмам — широко распространенный стандартный прием в сельском и лесном хозяйствах. Ферменты древоразрушающих грибов, утилизирующие лигнин, стали основой технологических процессов в древообрабатывающей промышленности многих стран. Контролирующие эти ферменты гены клонированы и перенесены в технологически удобные микроорганизмы (например, дрожжи). Нашли практическое применение даже фитопатогенные грибы. Споры некоторых видов накапливают в ферментерах и используют в качестве микогербицидов для опрыскивания засоренных посевов сельскохозяйственных культур. Несомненно, по мере изучения молекулярных механизмов фитопаразитизма горизонты практического применения грибов будут расширяться.
Список использованных источников
Катенин А. Е., Микориза растений Северо-Востока Европейской части СССР, Л., 1972.
Контримавичус В.Л. // Журн. общ. биологии. 1982. Т.423. Вып.3.
Мухин В.А., Степанова Н.Т. // Экология. 1976. №6. С.42—45.
Katharina Markmann, Gábor Giczey, Martin Parniske. // PLoS Biology, 2008. 6(3): e68.
Селиванов И.А. "Микосимбиотрофизм как форма консортивных связей в растительном покрове Советского Союза". М., 1981