Клас "Двостулкові молюски"
Зміст
Вступ
Розділ 1. Морфологічні, фізіологічні та екологічні особливості двостулкових
1.1 Загальна характеристика зовнішньої будови
1.2 Загальна характеристика внутрішньої будови молюсків
Розділ 2. Систематика класу
Розділ 3. Аналіз наукової та науково-популярної літератури, присвяченої двостулковим молюскам
Висновки
Список використаної літератури
Додатки
Вступ
Тваринний світ досить різноманітний. Тварини, а особливо безхребетні, зустрічаються майже у всіх біосферних оболонках.
В даний час посилено освоювання різних екосистем, значна частина досліджень припадає на водні екосистеми. Світ безхребетних дуже різноманітний і багаточисленний. Безхребетні тварини є однієї зі складові біосфери. Вони поширені у всіх середовищах існування: вони є й у ґрунті, і у воді, і в повітрі. Багато безхребетні можуть бути показниками якості навколишнього середовища.
Досить цікавим представниками фауни безхребетних є двостулкові молюски. У залежності від глибини водойми і її чистоти залежить кількість видів цих тварин. У прибережній, мулистій і більш чистій воді кількість видів більше, ніж у глибоких і забруднених місцях. Двостулкові молюски можна вважати біоіндикаторами водного середовища. Ці безхребетні тварини мешкають лише в чистих водоймах..
Мета: ознайомитися з біологічними особливостями представників класу Двостулкові молюски, їх значенням в природі, житті людини, з'ясувати сучасні напрямки їх вивчення.
Завдання:
1. ознайомитися з морфологічними, фізіологічними та екологічними особливостями двостулкових, систематикою класу;
2. встановити значення двостулкових молюсків;
3. провести аналіз наукової та науково-популярної літератури, присвяченої молюскам за останні 5 років;
4. з'ясувати сучасні напрямки їх вивчення.
Розділ 1. Морфологічні, фізіологічні та екологічні особливості двостулкових
1.1 Загальна характеристика зовнішньої будови
Двостулкові — виключно водяні тварини, що мешкають у морях, океанах та прісних водоймах. Розміри їх тіла — від кількох міліметрів до 1,5 метра (Tridacna gigas). Вони ведуть придонний спосіб життя і трапляються на різних глибинах. Більшість із них —повільно повзаючі форми, деякі нерухомо прикріплюються до субстрату. До цього класу належить близько 20000 видів. У прісних водоймах України їх налічують близько 150 видів, у Чорному та Азовському морях — понад 100 видів.
Двостулкові — це білатеральносиметричні молюски з двостулковою черепашкою, яка вкриває тіло з боків. Тіло їх здебільшого видовжене, більш-менш сплющене з боків і складається з тулуба, що заповнює верхню частину черепашки, і ноги, яка міститься на черевній стороні. Характерна особливість двостулкових — редукція голови. На передньому кінці тулуба розташований рот, на задньому — анальний отвір (рис. 1).
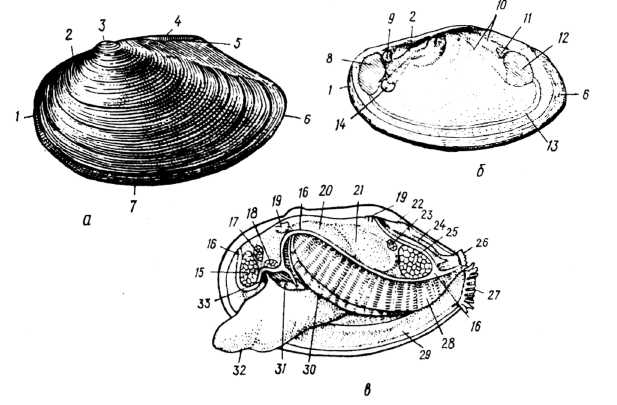
Рис. 1. Зовнішня будова беззубки (Anodonta cygnea): а - зовнішній вигляд черепашки; б — внутрішня поверхня правої стулки; в — вигляд тіла молюска зліва, черепашку видалено; 1 — передній край черепашки; 2, 5 — спинний (замковий край); 3 — маківка; 4 — зовнішній лігамент; 6 — задній край; 7 — черевний край; 8 — відбиток переднього м'яза-замикача; 9 — відбиток переднього ретрактора ноги; 10 — сліди росту та переміщення м'язів; 11 — відбиток заднього ретрактора ноги; 12 — відбиток заднього м'яза-замикача; 13 —мантійна лінія; 14 — протрактор ноги; 15 —передній м'яз-замикач; 16 — лінія, по якій відрізано лівий мантійний листок; 17 — передній ретрактор ноги; 18 — протрактор ноги; 19 — леватор (м'яз, що піднімає) ноги; 20 — кеберів орган; 21 — перикардій; 22 — спинний мантійний отвір; 23 — задній ретрактор ноги; 24 — спинний мантійний канал; 25 — задній м'яз-замикач; 26 — вивідний (анальний) сифон; 27 — ввідний (дихальний) сифон; 28 — ліва зовнішня напівзябра; 29 — мантія; 30 — ліва внутрішня напівзябра; 31 — ротова лопать; 32 — нога; 33 — рот
Ліва і права стулки черепашки рухомо з'єднані на спинній стороні еластичною зв'язкою — лігаментом, утвореним органічною речовиною — конхіоліном. Завдяки еластичності лігамента стулки при розслабленні м'язів-замикачів автоматично відкриваються. Крім того, у більшості двостулкових обидві стулки з'єднані між собою за допомогою замка, що складається із зубоподібних відростків внутрішньої поверхні спинного краю однієї стулки, які входять у заглибини протилежної. Деякі види, наприклад прісноводна беззубка (Anodonta), не мають замка, і стулки з'єднуються лише за допомогою лігамента.
Черепашки двостулкових мають різноманітну форму. У більшості обидві стулки однакові, проте є види, в яких вони різні. В устриці, наприклад, ліва стулка, якою вона прикріплюється до субстрату, значно більша й опукліша — у ній міститься все тіло м'якуна, тоді як права є лише покришкою. У деяких форм, наприклад у тередо деревоточця, або «корабельного черва» (Teredo) черепашка редукована: вона прикриває лише 1/20 довжини тіла.

Рис. 2. Схема будови черепашки та мантії двостулкових молюсків: 1, 2 — внутрішній війчастий та зовнішній епітелій мантії; 3 — екстрапаліальна порожнина; 4, 5, 6 — перламутровий, призматичний та роговий (периостракум) шари черепашки; 7, 8, 9 — зовнішня, середня та внутрішня лопаті мантії; 10 — клітини сполучної тканини; 11 — м'язи сполучнотканинного шару мантії
Закривається черепашка м 'язами-замикачами, яких буває два або один (за рахунок злиття двох м'язів або редукції одного з них). Вони мають вигляд товстих мускульних пучків, що йдуть упоперек тіла від однієї стулки до іншої (рис. 1, б, в). У місцях прикріплення м'язів до стулок на них утворюються відбитки, за якими можна зробити висновок про кількість та розташування м'язів-замикачів.
Під черепашкою розташована мантія, яка звішується з боків тіла у вигляді двох великих мантійних складок. Між мантією та черепашкою є дуже вузька екстрапаліальна порожнина (рис. 2), а між мантією і тілом залишається досить велика мантійна порожнина, в якій міститься мантійний комплекс органів: нога, зябра, осфрадії, ротові лопаті, ротовий, анальний, парні видільні та статеві отвори. На спинній стороні тіла обидві складки мантії зростаються. Коротенькими мантійними м'язами вільний край мантії прикріплюється до нижнього краю стулки черепашки, утворюючи на її внутрішній поверхні більш-менш глибокий слід — мантійну лінію (рис. 1, б). Край мантії, розташований нижче лінії прикріплення мантійних м'язів, потовщений і поділений вздовж на три лопаті — зовнішню, середню та внутрішню.
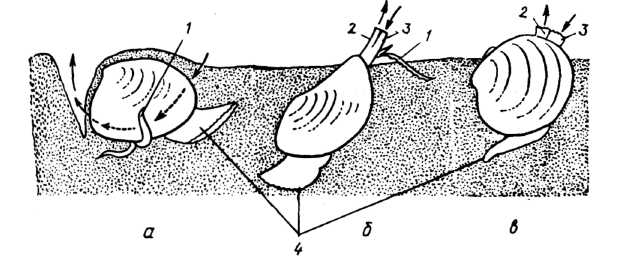
Рис. 3. Двостулкові молюски, які мешкають на поверхні дна та неглибоко закопуються в ґрунт: а, б — горіховидна нукула та куспідарія (надряд Protobranchia); в — циприна (надряд Autobranchia); 1 — ротові лопаті та придатки; 2 — вивідний сифон; 3 — ввідний сифон; 4 — нога. Стрілками позначено рух води
Краї мантійних складок у деяких місцях зростаються, залишаючи кілька отворів (2— 4), через які мантійна порожнина сполучається з оточуючим середовищем. Через такі отвори відбувається висування ноги, надходження води та їжі, вихід води, екскрементів, екскретів та статевих продуктів. Ввідний та вивідний отвори розташовані здебільшого на кінцях сифонів — порожнистих трубчастих виростів мантії (рис. 1, в). Сифони добре розвинені у форм, що зариваються в мул або пісок, у деяких видів їх довжина навіть перевищує довжину тіла (рис. 3, 4). Сифони можуть висуватися назовні і втягуватися всередину черепашки за допомогою особливих м'язів. У прісноводних перлівницевих (родина Unionidae) сифони короткі, а в кулькових (родина Sphaeriidae) їхня довжина може становити 0,3— 0,5 довжини тіла. По краю мантії можуть розташовуватись щупальця, очі, залози тощо.

Рис. 4. Двостулкові молюски, які глибоко закопуються в ґрунт: а — мія; б — скробікулярія; в — теліна; г — донакс (надряд Autobranchia); 1 — ввідний сифон; 2 — вивідний сифон; 3 — нога. Стрілками позначено рух води
Мантія — це багатошарове утворення (рис. 2). Зовні вона вкрита мантійним епітелієм, під яким міститься базальна мембрана, а під нею — потужний шар сполучної тканини (паренхіми), яка пронизана тонкими м'язовими волоконцями. Клітини сполучної тканини лежать пухко, між ними киї питаються великі міжклітинні простори, а також лакуни, заповнені гемолімфою. Внутрішня поверхня мантії вистелена війчастим епітелієм. У сполучній тканині, поблизу від епітеліальних шарів, залягають численні залозисті клітини.
Встановлено, що всі шари черепашки утворюються виділеннями залозистого епітелію зовнішньої лопаті мантії, причому конхіоліновий шар виділяється її внутрішньою поверхнею, а призматичний — зовнішньою. На решті зовнішньої поверхні всієї мантії виділяються речовини, що використовуються на побудову перламутрового шару черепашки.
Епітелій мантії відіграє важливу роль в адсорбції йонів кальцію із зовнішнього середовища шляхом піноцитозу. Крім того, кальцій переноситься до мантії гемолімфою з органів травлення. Застосуванням міченого Са45 було встановлено, що слиз, який суцільним шаром вкриває зябра, інтенсивно адсорбує кальцій із води. Шматочки слизу, відриваючись від зябер, разом з їжею потрапляють до органів травлення, де кальцій звільнюється і переноситься гемолімфою до мантії. Секрет залозистого епітелію мантії виділяється в екстрапаліальну порожнину. Він містить органічні речовини, що входять до складу матрикса (органічної основи) черепашки, а також значну кількість кальцію, який відкладається на цьому матриксі у вигляді СаСО>3>.
Черепашка росте разом з твариною протягом усього її життя, причому вона наростає по вільному краю. Верхівки стулок є найстарішими частинами черепашки, до яких у процесі росту додаються все нові й нові ділянки. Завдяки цьому можна відрізнити щорічний приріст та визначити вік тварин, якщо умови росту в різні сезони року різні. Узимку вповільнення росту помітне на черепашці у вигляді згущення ліній наростання, по яких і визначають вік тварин.
Із секреторною діяльністю мантійного епітелію деяких двостулкових пов'язане утворення перлин. Формування перлів — це захисна реакція організму на потрапляння в нього стороннього тіла: піщинки, уламку черепашки, частинок органічної речовини, паразита. Клітини мантії починають огортати чужорідне тіло зовнішнім шаром мантії з утворенням навколо нього епітеліального, так званого перлинного мішечка. Епітелій мішечка продовжує нормально функціонувати, виділяючи всередину спочатку трохи периостракума, потім призматичний шар, і кінець-кінцем —перламутровий шар, тобто в тій самій послідовності, що й при рості черепашки. Так утворюється вільна перлина. Найцінніші перлини здатні утворювати морські перлівниці (Pinctada margaritifera) та європейська річкова перлова скойка (Margaritifera margaritifera).
У мантійній порожнині двостулкових міститься орган руху — нога. Залежно від способу життя двостулкових нога може мати різну форму. У деяких найпримітивніших представників (надряд Ргоtobranchia) нога має плоску повзаючу підошву. Здебільшого ж вона клиноподібної форми, дуже сплюснута з боків і загострена на кінці (рис. 3, 4). Нога слугує для заривання в ґрунт і повільного повзання. У молюсків, що ведуть сидячий спосіб життя, нога значно редукується і може зовсім зникнути, як в устриць. У багатьох двостулкових на нижній поверхні ноги в особливій заглибині є так звана бісусна залоза, яка виділяє тягучу органічну речовину, що твердне у воді, перетворюючись на міцні нитки — бісус, за допомогою якого молюск прикріплюється до субстрату. Така бісусна залоза є, наприклад, у мідії (Mytilus), дрейсени (Dreissena) тощо.
Покриви двостулкових утворені шкірним епітелієм із великою кількістю слизових клітин та сполучною тканиною з окремими м'язовими волокнами й лакунами, заповненими гемолімфою. Епітелій у більшій частині війчастий (епітелій ноги, внутрішньої стінки мантії, зябер, ротових лопатей).
Мускулатура двостулкових поділяється на дві групи м'язів: одна — пов'язана з ногою, друга — з мантією та черепашкою (рис. 5). Нога має здебільшого дві (у Nuculidae — чотири) пари ретракторів, які прикріплюються до черепашки й при скороченні втягують ногу (рис. 1, в). Крім того, у нозі є безліч м'язових волокон, які розходяться у різних напрямах і утворюють майже справжній шкірно-м'язовий мішок, що спирається на кров'яний синус усередині ноги. Перед висуванням ноги в її синус нагнітається гемолімфа, яка при втягуванні ноги із синуса перекачується в центральний венозний синус, і об'єм ноги різко скорочується. Про м'язи мантії та черепашки вже говорилося.

Рис. 5. Мускулатура Bivalvia: 1 — передній м'яз-замикач; 2, 4 — ретрактори ноги; 3 — леватор (підіймач) ноги; 5 — янус; 6 — задній м'яз-замикач; 7, 8 — кільцеві та поздовжні м'язи сифонів; 9 протрактор ноги; 10 — рот
1.2 Загальна характеристика внутрішньої будови молюсків
Вторинна порожнина тіла (целом) у двостулкових складається з дуже невеличкого перикардія та порожнин гонад. Решта проміжків між внутрішніми органами заповнені сполучною тканиною — паренхімою, багатою на лакуни.
Травна система починається на передньому кінці тіла ротовим отвором, оточеним з боків двома парами ротових лопатей (ротові щупальця). З редукцією голови в двостулкових пов'язана відсутність у травному тракті глотки, радули, щелеп і слинних залоз.
Рот веде в короткий стравохід (ектодермальна передня кишка), вистелений миготливим епітелієм. Він відкривається в мішкоподібний шлунок, який належить до ентодермальної середньої кишки. Шлунок оточує парна печінкова залоза, яка складається з безлічі дрібних часток, що відкриваються кількома (їх може бути дві, три або багато) протоками в шлунок (рис. 6, а).
У задню частину шлунка відкривається сліпе мішкоподібне вип'ячування, у порожнині якого утворюється склоподібно-прозора драглиста паличка — кришталевий стовпчик (рис. 7). Це неклітинний гіаліновий стрижень, до складу якого входять мукопротеїни та ферменти, що розщеплюють вуглеводи (амілаза, глікогеназа тощо). Вільний кінець кришталевого стовпчика впирається в так званий гастричний (шлунковий) щит — невеличку прозору пластинку ущільненого секрету, яка міститься на передній стінці шлунка.

Рис. 6. Схема внутрішньої будови беззубки: а — поздовжній розріз через тіло, вигляд зліва; б — поперечний розріз та схема кровоносної системи; 1 — рот; 2 — мішок кришталевого стовпчика; З — стравохід; 4 — передній м'яз-замикач; 5 — печінка; 6 — шлунок; 7 — протока печінки; 8 — передня аорта; 9 — перикардій; 10 — шлуночок серця; 11 — передсердя; 12 — задня аорта; 13 — спинний мантійний отвір; 14 — задній ретрактор ноги; 15 — нирка; 16 — задня кишка; 17 — спинний мантійний канал; 18 — анальний отвір; 19 — вивідний сифон; 20 — задній м'яз-замикач; 21 — ввідний сифон; 22 — напівзябра; 23 — мантійний листок; 24 — середня кишка; 25 — гонада; 26 — нога; 27 — мантійна вена; 28 — виносні зяброві судини; 29 — венозна система нирок; 30 — нутрощева та ножна вени; 31 — судини від кеберового органа; 32 — лігамент; 33 — зяброва вена; 34 — зяброва артерія; 35 — верхня камера мантійної порожнини; 36 — приносні зяброві судини; 37 — стулка черепашки; 38 — нижня камера мантійної порожнини

Рис. 7. Цикл травлення в шлунку та травній залозі (печінці) двостулкових молюсків: а — шлунок перед прийняттям їжі; б — надходження їжі до шлунка та початок травлення і сортування; в — сортування харчових частинок у шлунку; г — внутрішньоклітинне травлення дрібних частинок у печінці та видалення великих частинок у кишку; д — шлунок порожній, підготовка до нового циклу; 1 — мішок кришталевого стовпчика; 2 — кришталевий стовпчик; 3 — гастричний щит; 4 - стравохід; 5 — печінка; 6 — фрагменти травних клітин з ферментами; 7 — великий тифлозоль; 8 — початок середньої кишки; 9, 10 — дрібні та великі харчові частинки
Більша частина внутрішньої поверхні шлунка має борозенчасті зони, вкриті війками і розділені вузькими провідними жолобками. Це сортувальні зони, де відбувається сортування харчових частинок.
Від шлунка відходить середня кишка, стінки якої утворюють дві довгі внутрішні складки — тифлозолі; часто тифлозолі вдаються в порожнину шлунка. Стінки середньої кишки також вистелені війчастим епітелієм. Середня кишка спускається від шлунка до основи ноги, утворює в ній кілька петель і прямує до спинної сторони тулуба, де переходить у задню кишку. Задня кишка звичайно пронизує шлуночок серця і закінчується порошицею на дні вивідного сифона (рис. 6, а).
Живляться двостулкові рослинним детритом та дрібними організмами, що завжди є в мулі, або придонними планктонними організмами, які молюски пасивно відфільтровують із води, що проходить через їх мантійну порожнину. Усе це зліплюється слизом у грудочки і прямує до харчових жолобків на нижніх краях зябер, а по них —до ротових лопатей. Ротові лопаті також вкриті війками і мають борозенки, по яких харчові грудочки просуваються до ротового отвору. Неїстівні частинки, так звані псевдофекалії, викидаються назовні.
Лише деякі морські форми є хижаками (переважно з надряду Septibranchia). Для молюсків-деревоточців їжею є, крім того, деревина. Отже, більшість двостулкових живиться дисперсними частинками, які збираються поверхнею зябер і ротових лопатей.
Їжа надходить до шлунка із стравоходу у вигляді стрічкоподібного слизового шнура з харчовими частинками. Завдяки биттю війок мішка, в якому міститься кришталевий стовпчик, він обертається навколо своєї осі й працює як коловерт, витягуючи слизовий шнур із стравоходу. Водночас при обертанні стовпчика, який треться об поверхню гастричного щита, утворюється суспензія із частинок їжі та рідини з ферментами, які вивільнюються при розчиненні кінчика кришталевого стовпчика в слабокислому середовищі шлунка. Далі ця суспензія потрапляє до сортувальних зон, де частинки діляться за розміром та вагою. Дрібні, легкі частинки переносяться до печінкових проток і втягуються в них. У печінці вони фагоцитуються і перетравлюються печінковими клітинами. Більші частинки "їжі із шлунка прямують в середню кишку і згодом викидаються з екскрементами. Фагоцитоз відбувається також і в кишечнику. В епітелії кишечника є амебоїдні клітини, здатні виповзати в просвіт середньої кишки, поглинати харчові частинки і потім повертатися до стінки кишки. Вуглеводи перетравлюються позаклітинно, у порожнині шлунка за допомогою ферментів кришталевого стовпчика, а білки й жири — переважно внутрішньоклітинно в печінці й середній кишці.
У хижих Septibranchia в шлунку немає кришталевого стовпчика та сортувальних зон; їх шлунок має добре розвинені м'язові стінки, а його епітелій вистелений хітиноїдною кутикулою. їжа (переважно дрібні ракоподібні) в такому шлунку розчавлюється й перетирається, а перетравлюється в печінці.
Видільна система Bivalvia складається з пари нирок, або боянусових органів. Вони лежать під перикардієм і мають вигляд мішкоподібних трубок, кожна з яких одним кінцем відкривається в перикардій, а другим — у мантійну порожнину біля основи ноги (рис. 6, а). У різних двостулкових форма нирок буває різною: найчастіше нирки складені вздовж, тому набувають V-подібної форми, у деяких вони S-подібно вигнуті. Отвір нирки в перикардій має клапан — невеличку складку, що перешкоджає зворотній течії рідини. Далі йде лійка — короткий тонкостінний канадець, вистелений миготливим епітелієм. За нею міститься залозистий відділ, внутрішня поверхня якого вистелена залозистим епітелієм і утворює численні складки, які вдаються в порожнину нирки. У них є густа мережа венозних лакун, проходячи якими гемолімфа залишає значну кількість продуктів дисиміляції. Залозистий відділ переходить у тонкостінний канадець, через який екскрети виводяться в мантійну порожнину.
Видільну функцію виконує також так званий кеберів орган, що є розростанням передньої стінки перикардія (рис. 1, в). Залозисті клітини кеберового органа вилучають із гемолімфи, яка омиває перикардій, продукти метаболізму і виділяють їх у порожнину останнього, звідки через нирки виводять їх назовні.
Кровоносна система в двостулкових незамкнена, складається із серця, кровоносних судин та системи лакун і синусів. Серце міститься на спинній стороні тіла і має здебільшого один шлуночок та два бічних передсердя (рис. 6). Воно оточене навколосерцевою сумкою —перикардієм, який утворений целомічним (перитонеальним) епітелієм. Кожне передсердя сполучається із шлуночком отвором, перекритим складчастим клапаном. Завдяки цьому при скороченні передсердь гемолімфа прямує лише від передсердь до шлуночка. Крізь шлуночок серця в більшості двостулкових проходить задня кишка, але порожнини їх не сполучаються. Від шлуночка до переднього і заднього кінців тіла відходять передня та задня аорти, які поділяються на артерії, що галузяться й врешті-решт відкриваються в лакуни.
Циркуляція гемолімфи забезпечується роботою серця й м'язів тіла. Насичена киснем гемолімфа з передсердь надходить у шлуночок серця, звідти виштовхується в судини, далі — в лакуни, де віддає кисень тканинам і насичується вуглекислим газом. З тканин венозна гемолімфа збирається в центральний венозний синус під перикардієм. Потім потрапляє до видільної системи, де звільнюється від продуктів обміну, а звідти переганяється в зябра. Зябровий апарат має складну систему судин і лакун, де гемолімфа збагачується киснем. Із зябер гемолімфа надходить у зяброві вени, які відкриваються в передсердя (рис. 6, б). Крім того, значна частина гемолімфи насичується киснем у мантії та ротових лопатях, які відіграють неабияку роль у газообміні; з цих органів по мантійних венах гемолімфа надходить також у зяброві вени.
Як уже згадувалось, у більшості двостулкових крізь шлуночок серця проходить задня кишка, проте в деяких форм серце лежить над кишкою або під нею. Це пояснюється тим, що під час ембріонального розвитку парний мезодермальний зачаток, з якого утворюються перикардій, серце й нирки, спочатку міститься обабіч кишки, а потім обидві його половини з'єднуються над і під кишкою, охоплюючи її у вигляді муфти, але в деяких форм таке злиття відбувається тільки над або під кишкою.
Гемолімфа двостулкових містить багато клітин, серед яких найбільше поширені різні типи амебоцитів; у деяких видів є й еритроцити. Гемолімфа може бути безбарвною (наприклад, у Anodonta), або червоною через наявність червоного пігменту гемоглобіну, який міститься в плазмі чи еритроцитах. Гемоглобін виконує функцію запасання кисню, який віддає в періоди, коли в оточуючому середовищі його мало (наприклад, при зариванні молюска в мул).
Гемолімфа двостулкових виконує різноманітні функції. Вона забезпечує сталість іонного складу та осмотичного тиску внутрішнього середовища організму; постачає тканинам кисень і поживні речовини; видаляє продукти обміну та вуглекислий газ. Крім того, амебоцити, зосереджені в лакунах нирок, накопичують у цитоплазмі кристалики екскретів і виносять їх у порожнину нирок. Захисна функція гемолімфи зумовлена тим, що амебоцитам притаманні фагоцитарні властивості. Так, при запальних явищах ці клітини енергійно фагоцитують мертві клітини, бактерій, а при паразитуванні личинок деяких трематод навколо них утворюється капсула з цих клітин. Нарешті, при пораненні клітинні елементи гемолімфи аглютинують, що припиняє кровотечу й призводить до загоєння рани. Гемолімфа двостулкових, як і інших молюсків, виконує ще функцію гідроскелета при рухах ноги. Висування ноги відбувається при розслабленні її м'язів, і надходженні до лакун гемолімфи, а втягування — при скороченні мускулатури і відтіканні гемолімфи в черевний венозний синус.
Органами дихання двостулкових є пара зябер, які розташовані в мантійній порожнині обабіч ноги. У примітивних форм (надряд Protobranchia) —це типові двопірчасті ктенідії, які складаються з осі та двох рядів трикутних зябрових пелюсток, що звисають у порожнину мантії (рис. 8, а). У більшості двостулкових (надряд Autobranchia) у зв'язку з переходом до живлення шляхом фільтрації води, зябра перетворилися на органи фільтрації і набули вигляду великих пластин, що займають більшу частину мантійної порожнини. У більшості Autobranchia, у тому числі й у прісноводних перлівниць, кожна із зябер складається з двох пластинчастих півзябер: зовнішньої, оберненої до мантії, і внутрішньої, оберненої до ноги. Півзябра складаються з численних тонких трубочок — зябрових ниток (філаментів), які утворюють низхідне (внутрішнє) та висхідне (зовнішнє) коліна. Філаменти з'єднуються між собою короткими поперечними сполучнотканинними перемичками, а висхідні та низхідні коліна кожного філамента сполучаються численними септами (рис. 8, в). Отже, кожна півзябра є двошаровою пластинкою, продірявленою дрібними порами, які ведуть до системи щілин у внутрішньозябровій порожнині. Проте в деяких молюсків (наприклад, у мідій) сусідні зяброві філгменти з'єднуються один із одним лише війками, тому зяброві пластинки легко розтріпуються на окремі нитки (рис. 8, б). Зяброві пластинки зверху і зсередини вкриті війчастим епітелієм, між епітеліальними клітинами містяться залозисті клітини, які виділяють слиз.

Рис. 8. Будова зябер представників трьох надрядів двостулкових (схематичні поперечні зрізи): а — Protobranchia; б—в — Autobranchia; г — Septibranchia; 1 — мантія; 2 — внутрішній листок ктенідія; 3 — зовнішній листок ктенідія; 4 — вісь ктенідія; 5 — лігамент; 6 — черепашка; 7 — тулуб; 8 — мантійна порожнина; 9 — нога; 10 — зовнішня зяброва нитка; 11 — внутрішня зяброва нитка; 12 — внутрішня напівзябра; 13 — зовнішня напівзябра; 14 — пора в зябровій септі; 15 — зяброва
Зябра зміцнюються сітчастим опорним «скелетом» — у кожному філаменті під епітелієм є потовщення сполучної тканини, тобто тонкі хітиноїдні палички.
Над кожною півзяброю від переднього кінця тіла до заднього тягнуться надзяброві канали, пов'язані з вивідним сифоном.
Узгоджена дія війок миготливого епітелію зябер, ротових лопатей, мантії й покривів тіла створює різницю гідростатичних тисків у надзябровій та підзябровій порожнинах, а також в останніх та зовнішньому середовищі. Внаслідок цього вода надходить через ввідний сифон або через широкий педальний отвір у мантійну порожнину, омиває зябра, через пори потрапляє до каналів, розташованих всередині зябер; із них, віддавши кисень і збагатившись вуглекислим газом, вода потрапляє в надзяброві канали і звідти викидається назовні через вивідний сифон (рис. 9).
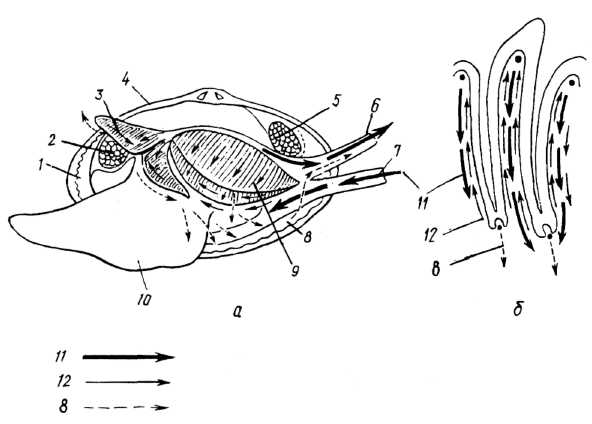
Рис. 9. Фільтрація води справжніми пластинчастозябровими (надряд Auto branchia) для дихання та живлення: а — загальна схема; б — струми води на зябрах мідії; 1 — мантія; 2 — передній м'яз-замикач; 3 — ротові лопаті; 4 — черепашка; 5 — задній м'яз замикач; 6 — вивідний сифон; 7 — ввідний сифон; 8 — шляхи псевдофекалій (неїстівних частинок); 9 — зябра; 10 — нога; 11 — напрям основних токів води, які надходять у мантійну порожнину та виходять з неї; 12 — фільтраційні та сортувальні струми на зябрах та ротових лопатях
У деяких двостулкових (підклас Septibranchia) ктенідії атрофуються, і в мантійній порожнині утворюється мускульна і іеретинка, яка поділяє її на дві ділянки — нижню та верхню, в якій і здійснюється газообмін (рис. 8, г).
Нервова система двостулкових розкидано-вузлового типу, вона складається з трьох пар нервових гангліїв: головних (цереброплевральних), ножних (педальних) і тулубних (вісце-ропарієтальних), з'єднаних між собою комісурами й конективами. Цереброплевральні ганглії лежать на передньому кінці тіла, здебільшого по боках від ротового отвору, обидва ганглії з'єднуються над стравоходом церебральною комісурою (рис. 10). Від них відходять нерви до ротових лопатей, передніх м'язів-замикачів та передньої частини мантії. У нозі іалягає пара педальних гангліїв, які з'єднуються з цереброплевральними за допомогою двох довгих конектив. Ганглії зближені між собою і зв'язані короткою комісурою; педальні ганглії іннервують ногу. Під заднім м'язом-замикачем залягає пара тісно зближених вісцеропарієтальних гангліїв. Вони іннервують задню частину мантії, задні м'язи-замикачі, зябра, осфрадії та нутрощі. Довгими конективами вони з'єднані з цереброплевральними гангліями, а в деяких видів — із педальними.

Рис. 10. Нервова система Anodonta (вигляд з лівої сторони): 1 — цереброплевропедальна конектива; 2 — церебральна комісура; З — нерв переднього м'яза іамикача; 4 — цереброплевральний ганглій; 5 — нерв ротової лопаті; 6 — цереброплевровісцеральна конектива; 7 — нирка; 8 — задній ретрактор ноги; 9 — нерв заднього м'яза-замикача; 10 — вісцеропаріетальний ганглій; 11 — нерви сифонів; 12 — мантійний нерв; 13 — педальний ганглій; 14 — нога
Відсутність голови, спрощення ротового апарату і малорухливий спосіб життя двостулкових зумовили слабкий розвиток у них органів чуття. Органів чуття, пов'язаних в інших молюсків з головою (очі, головні щупальця), у них немає.
Проте досить часто буває, що органи зору вторинно виникають або по всьому краю мантії (гребінець — Pecten), або по краю сифонів (серцевидка — Cardium), і навіть на зябрах (арка — Area). Такі вторинні очі мають різну будову — від очних ямок до таких відносно складно побудованих очей з кришталиком та сітківкою, як мантійні очі гребінців або тридакн ( рис. 11).
Органи дотику представлені чутливими клітинами, розкиданими по всій поверхні тіла — найбільше їх на ротових лопатях, нозі, краях сифонів та мантії.
Органи хімічного чуття — осфрадії та пов'язані з ними чутливі валки — є утвореннями, вкритими чутливими нервовими клітинами. їх функція — контроль якості води, яка надходить до мантійної порожнини. Вони є в різних місцях — на нозі, складках мантії, біля зябер, сифонів.
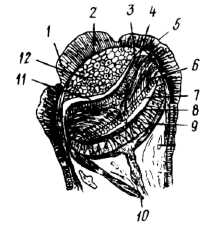
Рис. 11. Розріз мантійного ока Pecten islandicus: 1 — рогівка; 2 — сполучнотканинний шар; 3 — очна септа; 4 — дистальний шар сітківки; 5 — зовнішній епітелій ока; 6 — зорові клітини; 7 — шар світлочутливих паличок; 8 — тапетум; 9 — пігментний шар; 10 — оптичний нерв; 11 — лакунарний простір; 12 — кришталик
Органи рівноваги — статоцисти— у більшості двостулкових містяться в нозі, іноді — на спинній стороні тіла. Здебільшого їх два. Це пухирці, стінки яких складаються з чутливого епітелію, а всередині, у рідині, що їх заповнює, міститься один масивний статоліт або численні дрібні піщинки — статоконії. Переважна більшість двостулкових роздільностатеві, лише деякі групи — гермафродити, наприклад прісноводні кулькові (родина Sphaeriidae). Деяким двостулковим властива зміна статі протягом життя (морські перлові скойки). Гонади парні й містяться в основі ноги. Вони мають вигляд гроноподібних, розгалужених органів; порожнина їх є целомічним утворенням. При сильному розвитку гонади її вирости можуть заходити в тканини ноги (наприклад, у беззубки, рис. 6), або мантії (у мідії). У більшості двостулкових від гонад відходять коротенькі статеві протоки, що відкриваються в мантійну порожнину статевими отворами, які лежать поблизу від отворів нирок. Лише в деяких форм (надряд Protobranchia, а також роди Pecten, Ostrea) гонади не мають власних проток і відкриваються у нирки. Копулятивних органів немає, запліднення яєць переважно зовнішнє.
Більшість двостулкових відкладає яйця у воду, де й здійснюється запліднення. Проте багато прісноводних форм (родина Unionidae) відкладають яйця в зябра, де відбувається запліднення й розвиток до виходу личинки. У деяких видів проявляється турбота про нащадків. Самиці Cardium elegantulum відкладають яйця в дві виводкові сумки, утворені на стінках мантії (рис. 12, д); тут проходить розвиток зародка, без планктонної личинки. У дельфінозубої горіховидки (Nucula delphinodonta) яйця розвиваються в зовнішній шкірястій сумці, яка прикріплена до черепашки і сполучається з мантійною порожниною (рис. 12, б). Деякі двостулкові є живородними (наприклад, прісноводні Sphaeriidae та Pisidiidae).
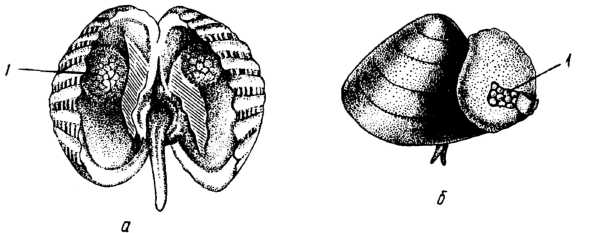
Рис. 12. Виводкові сумки двостулкових
У результаті спірального дробіння яйцеклітини утворюється личинка трохофорного типу (рис. 13, а). У багатьох нидів трохофора має зачаток ноги й первинну черепашку, яка спочатку закладається у вигляді однієї пластинки на спинній стороні. Згодом трохофора перетворюється на велігера — личинку, яка має вкритий війками диск — парус (велум), диостулкову черепашку, зачатки внутрішніх органів (рис. 13, в) За допомогою цих личинок, що ведуть планктонний спосіб життя і переносяться течіями, малорухливі молюски розселюються. У прісноводних уніонід з яєць, відкладених у зяброві порожнини самиць, виходять своєрідні личинки — глохідії (рис. 14), які деякий час паразитують на рибі.
Двостулкові молюски відіграють важливу роль у морських та прісноводних біоценозах. Як дорослі м'якуни, так і їхні личинки є важливими ланками в ланцюгах живлення. Дорослих двостулкових споживають морські зірки, хижі черевоногі та головоногі молюски, риби, птахи, ссавці. Личинками двостулкових живляться всі планктонні фільтруючі організми. Двостулкові є найефективнішими фільтраторами, які очищають воду.
Практичне значення двостулкових зумовлене тим, що багато їх видів людина здавна споживає як висококалорійну та смачну їжу (устриці, мідії, морські гребінці тощо); крім того, черепашки деяких молюсків використовують для виготовлення прикрас і ґудзиків, для інкрустаційних робіт. Перлові скойки дають цінні перла. Проте деякі види молюсків приносять шкоду людині, наприклад дрейсена, якою обростають днища суден і тим уповільнюється їх хід; вона оселяється в турбінних решітках електростанцій, у трубах водопроводів тощо, перешкоджаючи їх роботі.

Рис. 13. Метаморфоз мідії: а — трохофора; б — велігер; в — повзаюча личинка; г — доросла мідія; 1 — рот; 2 — прототрох; 3 — тім'яна пластинка; 4 — телотрох; 5 — педальний ганглій; 6 — ретрактор прототроха; 7 — парус; 8 — черепашка; 9 — вхід до мантійної порожнини; 10 — печінка; 11 — кишечник; 12 — анальний отвір; 13 — нога; 14 — вивідний сифон; 75 — ввідний сифон; 16 — фестончастий край мантії; 17 — нитки бісусу; 18 — передній кінець черепашки

Рис. 14. Глохідій Anodonta: 1 — черепашка; 2 — личинкова бісусна залоза; 3 — крайовий зубець; 4 — пучки чутливих щетинок; 5 — зубчики на крайовому зубці
Розділ 2. Систематика класу
В основі системи класу Двостулкові лежить будова черепашки, особливо її замка, та зябер. Останнім часом до діагностичних ознак відносять і деякі риси внутрішньої будови, зокрема будову шлунка. Клас Bivalvia поділяється на три надряди — Первиннозяброві (Protobranchia), Пластинчастозяброві (Autobranchia) та Перетинчастозяброві (Septibranchia):

НАДРЯД ПЕРВИННОЗЯБРОВІ (PROTOBRANCHIA)
Первиннозяброві — виключно морські форми, поширені по всьому Світовому океану. У закритих морях із низькою солоністю, наприклад в Азовському, Аральському, Каспійському, їх немає. Вони живуть практично на всіх глибинах — від літоралі до глибоководних западин (глибше 8 км), причому на великих глибинах це одна з основних груп молюсків.
Цей надряд об'єднує молюсків, які мають примітивні риси будови. Замок черепашки складається з однакових загострених зубів, розташованих перпендикулярно замковому краю, у деяких видів зуби редуковані; зябра мають вигляд двопірчастих ктенідіїв. Шлунок первиннозябрових має відносно просту будову з однією-трьома протоками печінки; тифлозолі кишки не вдаються в його порожнину. Нога з широкою плоскою повзальною підошвою. При втягуванні в черепашку підошва згортається вздовж по середній лінії. Нирки та статеві залози відкриваються в мантійну порожнину єдиною порою. Ротові лопаті здебільшого добре розвинені й складаються з пари листочків та довгого щупальцеподібного придатка, за допомогою якого молюск збирає детрит. Ці молюски ніколи не утворюють бісус і не прикріплюються до субстрату. За способом живлення вони — збирачі детриту.

Рис. 15. Надряд Protobranchia: будова тонкої горіховидки (Leionucula tenuis): а — права стулка зі спинної сторони; б — вигляд тіла збоку, праву стулку та листок мантії видалено; 1 — гребінчасті замкові зуби; 2 — м'язи ноги; 3 — вирости краю мантії між зубами; 4 — нутрощевий мішок; 5 — передній м'яз замикач; б — ділянка правого листка мантії; 7 — рот; 8 — ротова лопать; 9 — нога; 10 — підошва ноги; 11 — лівий листок мантії; 12 — придаток ротової лопаті; 13 — правий ктенідій; 14 — задній м'яз-замикач
Надряд Protobranchia об'єднує два ряди: Солеміїди (Solemyida) та Нукуліди (Nuculida); останній — найбільш багатий видами, поширеними переважно в морях північної півкулі. Тут дуже часто трапляються представники родини Nuculidae, наприклад горіховидки (Nucula). У цих молюсків замок складається з великої кількості гострих зубів, що нагадують зубці гребінки. У прибережній смузі північних морів Європи мешкає тонка горіховидка (Leionucula tenuis, рис. 15).
У північних морях поширені також представники родини Nuculanidae, серед яких численними є види родів Yoldia, Portlandia, Nuculana; останні утворюють щільні поселення і тому відіграють неабияку роль у бентосних біоценозах.
Значно менший ряд Solemyida включає види, що живуть у товщі мулу без зв'язку з відкритою водою —дуже рідкісний випадок серед молюсків. Наприклад, Solemya borealis, довжина черепашки якої досягає 5 см, має великі зябра і редукований кишечник; цих молюсків ще недостатньо вивчено.
НАДРЯД ПЛАСТИНЧАСТОЗЯБРОВІ (AUTOBRANCHIA)
До цього надряду належить більшість двостулкових. Вони заселяють усі моря та океани, а також прісні водойми, тряпляються на різних глибинах: від урізу води до найглибших западин.
Для Autobranchia характерні черепашки із замком, зуби якого розташовані по радіусах від верхівки, і лише вторинно вони можуть займати інше положення або зовсім зникати. Зябра в них збільшені, з дуже витягнутими листками (філаментами); кожен із філаментів напрямлений від нерухомої осі ктенідія вентрально (низхідне коліно), потім після перегину —дорзально (висхідне коліно). У порожнину шлунка з кишки вдається великий тифлозоль. Печінка відкривається в шлунок численними отворами. Нога клиноподібна, позбавлена плоскої підошви, у молодих особин завжди з бісусом, іноді редукується.
На відміну від первиннозябрових, які збирають харчові частинки придатками ротових лопатей з поверхні ґрунту, основна маса пластинчастозябрових живиться шляхом фільтрації води та відціджування з неї харчових частинок. їхні зябра перетворені на фільтр-сито з війчастим водорухаючим епітелієм.
Ця група двостулкових надзвичайно різноманітна за розмірами, формою черепашки, способом життя; серед них є й повзаючі, й плаваючі, й стрибаючі, й прикріплені форми.
До надряду Autobranchia відносять сім — дев'ять (за даними різних авторів) рядів та більше 120 родин. Наведемо найважливіші з них.
Ряд Уніоніди (Unionida). До цього ряду належать великі прісноводні молюски. Для них характерні більш-менш добре виражений перламутровий шар черепашки, наявність розвиненої сокироподібної ноги та зовнішнього лігамента черепашки. Краї мантії не зрощені, сифони дуже короткі. Замкові зуби здебільшого поділяються на дві групи: горбкоподібні центральні, які розташовані біля маківки черепашки, та пластинчасті бічні, витягнуті вздовж задньоспинного краю черепашки. Іноді замкових зубів немає, наприклад у беззубки (Anodonta).
Яйця уніоніди виношують усередині зябрових порожнин материнських особин. Ці порожнини відкрають роль інкубатора, де з яєць виходять личинки — глохідії. Глохідій має двостулкову черепашку з одним м'язом-зами-качем. На черевному краю кожної стулки є гострий зубець, у середній частині тіла міститься личинкова бісусна залоза. Молюск виштовхує глохідіїв через вивідний сифон у воду,коли повз нього пропливає риба. Личинка прикріплюється до зябер чи плавців риби за допомогою клейкої нитки бісуса та краєвих зубців черепашки. Занурюючись потім у тканини риби, глохідій оточується цистою і веде деякий час паразитичний спосіб життя; через один-два місяці сформований молюск залишає рибу й переходить до самостійного життя. За допомогою риб уніоніди розселюються, проникаючи у верхів'я річок.
Серед уніонід широко відома родина Прісноводних перлових скойок (Margaritiferidae), поширена в Євразії та Північній Америці. Саме ці молюски здатні утворювати перлини, які здавна цінували як прикраси. На півночі Європи відома європейська річкова, або перлова скойка (Margaritifera margaritifera), з довжиною черепашки до 12 см (рис. 16). Оскільки ці молюски живуть лише в дуже чистих річках із швидкою течією, то через забруднення європейських річок стічними водами вони майже зникли. У річках України перлових скойок немає, в Росії вони залишились тільки в річках Кольського півострова, проте популяції їх дуже нечисленні.
У річках України найпоширенішими є представники родини перлівницевих (Unionidae), яка представлена видами родів беззубок (Anodonta), перлівниць (Unio) та красіан (Crassiana). Черепашки беззубки переважно тонкостінні, з тонким перламутровим шаром. В Україні найпоширеніший вид — беззубка лебедина (A. cygnea), завдовжки до 20 см, яка трапляється переважно в річках із повільною течією та замуленим піщаним дном. Перлівниці мають товсту черепашку з добре розвиненим перламутровим шаром. Особливо товстостінна черепашка в товстої скойки (Crassiana crassa), саме її черепашки використовують для виготовлення перламутрових прикрас та ґудзиків.
Ряд Мітиліди (Mytilida). Це виключно морські двостулкові, переважно прикріплені бісусом до субстрату. У більшості з них спостерігається редукція і навіть втрата одного із м'язів-замикачів.
До цього ряду належать важливі промислові молюски: їстівні устриці та мідії (підряд Mytilenina), а також Справжні перлові скойки (підряд Pteriina), які утворюють цінні перли.
Устриці (родини Ostreidae та Crassostreidae) мають нерівну, ребристу черепашку з неоднаковими стулками: ліва (нижня) стулка опукліша, глибша, з більш виступаючою маківкою. Молюск цементується лівою стулкою до субстрату, набуваючи його рельєфу і повторюючи його нерівності. М'яз-замикач один, займає середину стулки. Мантія відкрита, не утворює сифонів; вода входить через передній край черепашки та виходить через черевний та задній краї. У дорослих форм ноги немає, хоча в молоді вона є (рис. 42). Доижина черепашки 8—12 см, але велетенська устриця (dassostrea gigas) може досягати 38 см.
Відомо близько 50 видів устриць, які майже всі тепловодні. Живуть вони переважно на кам'янистих ґрунтах на глибинах від 1 до 50—70 м як поодинці, так і великими скупченнями, утворюючи берегові поселення та банки, де щільність їх така висока, що окремі особини зростаються разом, утворюючи великі зрощення.
Устриць вважають смачним делікатесним продуктом, їх здавна виловлювали у великих кількостях. Один із основних промислових видів — Ostrea edulis — мешкає біля берегів Європи, у тому числі в Середземному та Чорному морях. Деякі вчені вважають, що чорноморська устриця — це окремий вид О. taurica.
З давніх часів людина використовувала в їжу також мідії (родина Mytilidae). Вони мають гладеньку черепашку, маківка якої дуже зсунута до звуженого переднього краю, і добре розвинену бісусну залозу. Задній м'яз-замикач значно більший, ніж передній. Довжина черепашки може досягати 10 см. Вони населяють літораль і глибини до 80 м, утворюючи щільні поселення.
Найбільш відомим та поширеним видом є їстівна мідія (Mytilus edulis), яка мешкає вздовж Атлантичного узбережжя Європи, біля берегів Ісландії, південної частини Гренландії, берегів Північної Америки, у Баренцевому, Білому, Балтійському, а також далекосхідних морях. У Чорному морі мешкає близький вид — чорноморська мідія (М. galloprovincialis).
Представники підряду Pteriina мають лише один м'яз-замикач, ногу із добре розвиненою бісусною залозою і могутній перламутровий шар. Ці молюски здатні утворювати досить великі перли. Найкращі та найцінніші перли продукують види роду Pinctada та Pteria. Саме їх називають справжніми перловими скойками. У них велика черепашка здебільшого округлої форми, з прямим замковим краєм, витягнутим ззаду у вухоподібний виріст.
Найбільша з перлових скойок — Pinctada margaritifera (рис. 18, а) — досягає 30 см в діаметрі черепашки та маси 10 кг, хоча такі великі екземпляри трапляються рідко. Цей вид живе в Тихому та Індійському океанах, утворює щільні поселення — банки.
Основні промисли морських перлів зосереджені в Перській затоці, біля острова Шри-Ланка, у Червоному морі, біля берегів Австралії, Японії, уздовж узбережжя Венесуели, Панами, Мексики та в деяких інших місцях.
Близькою до перлових скойок є родина пін (Pinnidae), які мають велику, до 30 см, клиноподібної форми черепашку без замкових зубів, із радіальними ребрами та зігнутими лусками (рис. 18, б). Піни мешкають на невеликих глибинах у Середземному морі, Атлантичному та Індійському океанах, вони закопуються в ґрунт звуженим переднім кінцем і закріплюються бісусом, а заднім піднімаються над поверхнею дна.
Ряд Пектиніди (Pectinida). З цього ряду найбільш відомими є морські гребінці (родини Pectinidae та Propeamussiidae), поширені майже в усіх морях та океанах на різних глибинах, включаючи найглибші западини. Особливо численні й різноманітні вони на прибережних мілководдях субтропічної та помірної зон Світового океану.
Гребінці мають округлу черепашку з прямим замковим краєм, який видається спереду та ззаду у вигляді виступів — пушок (рис. 19, а). Верхня (ліва) стулка плоскіша, нижня (права) — опукліша. Поверхня черепашки має радіальні або концентричні ребра, часто з шипами або лусками. У мілководних гребінців (Pecten, Chlamys та ін.) черепашка переважно велика, товстостінна; у глибоководних (Amussium, Propeamussium, Delectopecten) стулки крихкі, напівпрозорі.
Нога гребінців слабо розвинена, рудиментарна і має вигляд пальцеподібного вироста. Молодь прикріплюється до субстрату бісусом, а дорослі втрачають цю здатність, хоча відомі й винятки. З усіх двостулкових молюсків гребінці найбільш рухливі. Вони можуть плавати та підстрибувати, періодично хлопаючи стулками і виштовхуючи з-під них воду (рис. 19, б). Цьому сприяє особлива будова єдиного м'яза-замикача та мантії, краї якої звисають з-під черепашки. Край мантії облямовують численні щупальця (органи дотику) та очі, які сприяють орієнтації тварини при плаванні.
У Баренцевому, Білому й далекосхідних морях, біля берегів Скандинавії та Атлантичного узбережжя Північної Америки на глибинах до 100 м мешкає досить великий (діаметр черепашки 8 см) ісландський гребінець (Chlamys islandicus). Його м'ясо дуже смачне, тому він є об'єктом промислу. У далекосхідних морях є ще й приморський гребінець (Patmopecten yessoensis). У Чорному морі трапляється лише один вид — чорноморський гребінець (Flexopecten ponticus), який має яскраву черепашку діаметром 5 см з відтінками жовтого, оранжевого, рожевого, червоного кольорів.
Ряд Люциніди (Lucinida). Це дуже різноманітна й багата видами група, вона включає близько 30 родин; її представники мешкають як у морях, так і в прісних водоймах.
Серед морських форм найбільш відомими є представники родини Astartidae, які дуже поширені в морях північної півкулі і складають там звичайний компонент донних біоценозів, наприклад зубчаста астарта (Astarte crenata) та північна астарта (A. borealis).
До цього самого ряду належать прісноводні молюски родин горошинкових (Pisididae) та кулькових (Sphaeriidae). Це дрібні молюски (найменші види до 2 мм), найбільші серед них види роду Sphaerium, наприклад кулька рогова (S. rivico1а, рис. 20) досягає 2,5 см. Кульки живородні: їх яйця розвиваються у виводкових камерах, розташованих на внутрішніх напівзябрах; із камер виходить повністю сформована молодь.
Кулькові поширені в прісних водоймах усіх материків, крім Антарктиди. При пересиханні водойм вони можуть навіть до шести місяців перебувати без води, зарившись у мул.
Деякі види роду горошинки (Pisidium) опанували незвичайний для двостулкових тип біотопу: вони живуть у заболочених ґрунтах, і їм для дихання та живлення вистачає тієї незначної кількості води, що є між частинками ґрунту.
Серед невеликих (до 5 см), але дуже гарних морських люцинід слід назвати представників родини Donacidae, які
мають витягнуту округло-трикутну черепашку зі зсунутою до заднього кінця маківкою. Для них характерні яскраві кольорові промені, що йдуть радіально від маківки до черевного краю, а також плями на внутрішній поверхні черепашки. Види роду Donax звуть через це морськими метеликами. Ці молюски добре закопуються в ґрунт і навіть пересуваються в його товщі за допомогою великої ноги.
Ряд Венериди (Venerida). Це найбільший за об'ємом ряд двостулкових, що об'єднує близько 40 родин. Представники ряду живуть як у морях, так і в прісних водоймах.
Найзнаменитішою та найпопулярнішою родиною є тридакніди (Tridacnidae), що об'єднує найбільших двостулкових молюсків. Серед них найбільш відомі тридакни (рід Tridacnа). Велетенська тридакна (Т. gigas) досягає майже 1,5 м у довжину і маси 250 кг, причому основна маса припадає на черепашку.
Тридакни — мешканці тропічних мілководь. Вони трапляються в Індійському та Тихому океанах у зонах коралових рифів, де нерухомо лежать на ґрунті спинною стороною, при цьому черевна сторона обернена догори (рис. 21). Нога, яка виділяє бісус, міститься на спинній стороні, і бісус виходить із черепашки біля її маківки. Відповідно зсунуті й сифони: ввідний (спинний) змістився в передньочеревний відділ, а вивідний напрямлений догори і лежить посередині черевного краю. Краї мантії зрослися по всій довжині, крім місць виходу сифонів та бісусу. У потовщеному краї мантії живе безліч одноклітинних джгутикових — зооксантел (Symbiodi-nium microadriaticum, ряд Dinoflagellida). Тридакни мають ряд пристосувань для покращення умов існування симбіонтів. По краях мантії у Т. сгосеа та Т. elongata виявлено так звані гіалінові органи. Це глечикоподібні утворення розміромблизько 1 мм, заповнені прозорими клітинами. Навколо таких органів розташовані скупчення зооксантел. Припускають, що гіалінові органи концентрують світло, яке падає на мантію, і розсіюють його в прилеглих тканинах, покращуючи умови фотосинтезу водоростей. Крім того, краї мантії тридакн мають дуже яскраве забарвлення.
Вважають, що рясна пігментація краю мантії тридакни також має адаптивне значення: захист хлорофілу симбіонтів від надмірної дії сонячного проміння в денні часи. Скорочуючи та розправляючи пігментні клітини мантії, молюски можуть ефективно регулювати рівень сонячної радіації, яка досягає симбіонтів. Тридакни дістають від симбіонтів продукти їх фотосинтезу (глюкозу, глюкозофосфат, ліпіди), а симбіонти споживають продукти азотистого обміну хазяїна (аміак, солі амонію), амінокислоти, вуглекислий газ.
Родина серцевидок, або кардіїд (Cardiidae) — це величезна група мілководних форм, що мешкають переважно в теплих морях. їхня черепашка з радіальними ребрами нагадує серце (рис. 22, а). У Чорному морі поширена їстівна серцевидка Ламарка (Cerastoderma lamarckii) та кілька інших видів.
Заслуговує на увагу родина Glossidae. У Середземному морі та Атлантичному океані живе молюск, що називається морське серце (Glossus humanus). Його тонкостінна черепашка довжиною до 5 см має опуклі, сильно закручені маківки (рис. 22, б).
Велика родина венерид (Veneridae) поширена в помірних і тропічних морях на піщаних ґрунтах мілководдя. Черепашки венерид дуже гарні, різноманітних розмірів та форм. Багато з них є об'єктами промислу. У Чорному морі вздовж берегів мешкає кілька видів венерид. Один із найбільш поширених видів —венуспівник (Chamelea gallina).
У Чорному морі часто трапляється представник родини морські черенки (Solenidae) — звичайний морський черенок (Solen vagkia) з довгою, до 12 см, черепашкою майже прямокутної форми (рис. 22, в). Вони живуть на глибинах до 10 м і можуть дуже швидко закопуватись у ґрунт, виставляючи назовні пару сифонів, а також стрибати реактивним способом за допомогою струменя води, що викидається з сифонів.
У прісних водоймах дуже поширені сидячі молюски родини дрейсеніди (Dreissenidae). Найпоширенішим у водоймах Європи видом є річкова дрейсена (Dreissena polymorpha), трикутна черепашка якої має довжину 4— 5 см і смугастий малюнок (рис. 23). Цей вид проникає в лимани Чорного моря, а також опріснені ділянки Азовського моря. На півдні України трапляється ще й бугська дрейсена (D. bugensis), яка в наш час активно розселюється. Річкова дрейсена бісусом прикріплюється до субстрату і утворює величезні скупчення, обростаючи каміння, сваї, різні гідротехнічні споруди, а також водостоки, труби, по яких вода тече до турбін, захисні ґрати тощо. Це призводить до великих затрат на очищення зазначених споруд.
До ряду венерид належать також дві родини, представники яких пристосувалися до життя в ходах, проточених ними в скелях, вапняках, деревині. Представники родини свердлунових (Pholadidae) свердлять тверді породи за допомогою своєї черепашки. Черепашка фоладід позбавлена лігамента, що різко підвищує взаємну рухливість стулок. Особливе розташування м'язів-замикачів призводить до того, що стулки черепашки поперемінно розсуваються то в передній, то в задній частині. Завдяки таким рухам стулок, озброєних ребрами з шипами та горбками, відбувається свердління субстрату. У Чорному морі поширений свердлуй звичайний (Pholas dactylus).
Представники родини деревоточців (Teredinidae) пристосувалися до свердління деревини. До цієї родини належать два роди: Bankia й Teredo; вони мають видовжене червоподібне тіло, тому їх називають «корабельними червами». Напередньому кінці тіла міститься дуже маленька (1/30 — 1/40 частина загальної довжини тіла) черепашка, озброєна гострими гребенями (рис. 24). При розсуванні стулок черепашки гострі зубці, що є на ребрах стулок, здирають шар деревини.
На задньому кінці тіла містяться два тонких сифони, а також пара вапнякових пластинок — палеток, які захищають тіло молюска, закриваючи вхідний отвір у хід. Стінки ходу молюск покриває зсередини тонким шаром вапнякових відкладів. Тередові використовують деревину не тільки як сховище, а й як їжу. Дрібні частинки деревини перетравлюються внутрішньоклітинно амебоцитами печінки. Крім деревини, в їжу використовуються планктонні організми, які втягуються через ввідний сифон.
Деревоточці пошкоджують деревину. Особливо велику небезпеку вони становлять для дерев'яних споруд причалів, а також для дерев'яних суден. У Чорному морі відомі три види деревоточців: Teredo navalis, T. pedicellatus та Т. utriculus; перші два види проникли в Азовське море.
НАДРЯД ПЕРЕТИНЧАСТОЗЯБРОВІ (SEPTIBRANCHIA)
Перетинчастозяброві — це морські, переважно глибоководні молюски. Вони трапляються лише в морях з океанічною солоністю, тому в Чорному морі їх немає. Цей надряд включає чотири ряди та кілька родин.
У перетинчастозябрових зябра маленькі, редуковані або перетворені на мускулясту перетинку (септу), яка проходить уздовж черепашки і поділяє мантійну порожнину на верхній та нижній відділи. Зяброву септу пронизують кілька отворів, через які сполучаються верхня та нижня камери мантійної порожнини. Черепашка — з редукованим замком. Шлунок повністю вкритий хітиноїдною вистилкою, яка закриває навіть сортувальне поле. Тифлозолі розвинені слабо. Печінка складається з невеликої кількості часток та відкривається в шлунок двома отворами. Нога клиноподібна, з поздовжньою борозною, іноді частково редукована. За способом живлення це хижаки.
Найчисленнішою родиною цього надряд у є родина Cuspidariidae (ряд Cuspidariida), що мають витягнену у вигляді трубки задню частину черепашки, в якій залягають сифони (рис. 25). їхні зябра редуковані. Водообмін у зябровій порожнині здійснюється шляхом почергового скорочення та розслаблення зябрової септи. Різке скорочення зябрової септи забезпечує різке втягування води через ввідний сифон —«вдих». Потім вода переганяється через отвори з нижньої камери у верхню, отвори замикаються, вода викидається через анальний сифон — відбувається «видих». Ці молюски — хижаки, різкими «вдихами» вони затягують у мантійну порожнину дрібних планктонних тварин. Хітиноїдна вистилка шлунка та його м'язові стінки сприяють розчавлюванню та перетиранню здобичі —переважно дрібних ракоподібних. Функцію дихання виконує внутрішня поверхня мантійної порожнини, яка пронизана кровоносними судинами.
Розділ 3. Аналіз наукової та науково-популярної літератури, присвяченої двостулковим молюскам
З метою з`ясування сучасного стану наукових досліджень двостулкових молюсків ознайомилися з такими виданнями: Вестник зоологии, Наука и жизнь, Світ науки, Химия и жизнь, Природа, Юный натуралист, Гидробиологический журнал.
Кількість статей, присвячених двостулковим молюскам розподілена таким чином (таблиця 1).
|
Назва журналу |
Рік видання |
Номери, в яких надруковано матеріали |
|
Вестник зоологии |
2000 |
1 – 2, 3 |
|
2001 |
1, 3, 4 |
|
|
2002 |
1, 5, 6 |
|
|
2004 |
2, 3 |
|
|
Гидробиологический журнал |
2003 |
2, 3, 4, 6 |
|
2004 |
1, 3 |
|
|
2005 |
2, 4 |
|
|
Природа |
1990 |
6 |
|
2000 |
12 |
|
|
Химия и жизнь |
1988 |
6, 11 |
|
Юный натуралист |
2003 |
5, 6, |
Таким чином за останні 5 років було опубліковано 28 статей про двостулкових молюсків.
Статті, які надруковані в журналах присвячені наступним питанням:
значення двостулкових молюсків (молюски займають важливе місце в природних водних системах та відіграють значну роль в живленні риб, спостерігається в деяких випадках, з одного боку і негативна дія молюсків, як наприклад, мідійне обростання гідрологічних споруд, але з іншого боку таким чином мідії стають складовою природного біофільтра);
морфологія, екологія та систематика (розглядається питання порового апарату раковин молюсків, порівняльний каріологічний аналіз двох видів роду Unio, зміни морфологічних ознак раковин, особливості екології і біології розмноження молюсків, особливості екології та морфології, особливості хоботного апарату цестоди, морфологія та особливості біології молюсків Камчатки та Північних Курил);
зоогеографія (дослідження донних безхребетних низового Дунаю);
екологічна фізіологія та біохімія (поліплоїдія і анеуплоїдія у молюсків, особливості накопичення іонів важких металів в організмі двостулкових молюсків, дія деяких амфіфільних речовин і сумішей препаратів на морських молюсків, вплив фенолів на вміст глюкози в організмі перлівниці, вплив умов існування на зв`язування важких металів в організмі молюска);
методи дослідження (ймовірні напрямки еволюції каріотипів європейських представників молюсків в світі сучасних уявлень про філогенію родини, новий вид фауни молюсків та його діагностика і можливі шляхи інтродукції, дослідження видового складу прісноводних молюсків та стратегія їх охорони, молюски як седиментатори радіоактивних речовин, сезонний ріс деяких прісноводних двостулкових молюсків).
Висновки
Молюски - тварини з м'яким несегментованим тілом, що поділяється на три відділи - голову, тулуб, ногу. Тіло вкрито шкірястою мантією, яка властива лише молюскам. Між мантією і внутрішніми органами знаходиться мантійна порожнина, яка з'єднується з навколишнім середовищем ввідним та вивідним сифонами. Мантія виділяє черепашку, яка виконує 3 функції: захисну; опорну, як зовнішній скелет, до якого прикріплюються м'які частини тіла; рухому.
Кожен рік черепашка збільшується: світла смужка утворюється влітку, темна - взимку. На ній можна побачити річні кільця, за якими ми можемо визначити скільки років живе молюск. Порожнина тіла змішана, утворена залишками первинної порожнини і сильноредукованим целомом. У первинній порожнині, виповненій паренхімою, розташовані внутрішні органи. Залишки целому представлені навколосерцевою сумкою і порожниною статевих органів.
Кровоносна система незамкнена. В них з'являється серце, яке може бути 2-х, 3-х камерним і навіть 5-ти камерним. Кров може бути червоною, рожевою в залежності від концентрації гемоглобіну та блакитною. Нервова система складається із 3-5 пар добре розвинених гангліїв, які або розташовані у різних частинах тіла і з'єднані між собою нервовими тяжами, або зливаються в єдиний, утворюючи праобраз головного мозку. В залежності від розвитку нервової системи розвиваються органи чуття. У них є очі, органи дотику, хімічного чуття (осфрадії), рівноваги /статоцисти/. В них з'являється орган руху - нога. У більшості молюсків розвинені гладенькі м'язи, тому рухи їх повільні. Лише у і головоногих, які активно і швидко рухаються, м'язи поперечносмугасті.
Органами дихання є або зябра, - розташовані у мантійній порожнині, або легені. Функції легень виконують стінки мантійної порожнини, пронизані кровоносними судинами. Органи виділення - нирки являють собою видозмінені метанефридії. Одним кінцем нирка відкривається в навколосерцеву порожнину (целом), іншим - у мантійну порожнину.
Двостулкові молюски поділяється на 3 надряди — Первиннозяброві (Protobranchia), Пластинчастозяброві (Autobranchia) та Перетинчастозяброві (Septibranchia).
Досить багато написано статей та проведено досліджень щодо особливостей будови, фізіологічних функцій, поширення, пристосування до умов існування Двостулкових молюсків, але для вчених ще існує досить широке поле діяльності в даному напрямки досліджень.
Список використаної літератури
Бондарёва А. Лечат ли... молюски? // Комсом. знамя. – 1988. - № 3. – С.49 -57.
Галина М.С., Петров С.А. Молюски в загрязненном море. // Химия и жизнь. – 1988. - № 6. – С. 52 – 53.
Давидов А.І. Молюски-друзі, молюски – вороги. – К.: Знання, 1975. – 48 с.
Дунаева Ю.А. Трофейное оружие…. моллюсков.// Биология. – 2003. - № 45. – С. 14 – 15.
Зверев И.Д. Книга для чтения по зоологии. — М.: Просвещение, 1971.
Жизнь животных: В 6-ти томах. – М., 1985.
Киричок Е. Природные фильтры (Двустворчатые моллюски). // Юный натуралист. – 2003. - № 6.
Книга для читання по зоології.: Посібник для вчителів. – К., 1985.
Кораго А.А. Жемчуг. // Природа. – 1990. - № 6. – С. 38 – 43.
Конюшин А.В. Пресноводные двустворчатые молюски или Происхождение фауны Австралии. // Природа. – 2000. - № 12. – С. 28 – 36.
Коштовності узбережжя: (Черепашки). //Юний натураліст. – 2003. - № 5. – С. 9.
Молюски // Книга для чтения по зоологи. – М., 1986. – С. 36 – 49.
Незмен Л.П. Речная жемчужница: (Двустворчатый моллюск). // Юный натуралист. – 1991. – № 3. – С. 18 – 19.
Петухов С.А. металлические терки моллюсков. // Химия и жизнь. – 1988. - № 11. – С. 42 – 45.
Рыбка хвостиком махнула: о колониях моллюсков. // Юный натуралист. – 1999. - № 7. – С. 23 – 24.
Сергеев Б.Ф. Кто охотник? (Самый крупный двустворчатый моллюск – триданка). // Юный натуралист. – 1998. - № 10. – С. 12.
Савчук М.П. Зоологія безхребетних. – К., 1965. – 503 с.
Согур Л.М. Зоологія. Курс лекцій. – К.: Фітосоціоцентр, 2004. – 308 с.
Тип Молюски. // Биология./ Под ред. В.А.Глумовой. – Ижевск, 1995. – С. 94 – 98.
Фауна України: В 40 томах. Редк. Топачевський В.А. та ін. Т.29. Молюски. – К., 1985.
Федяков В.В., Наумов А.Д. Двустворчатые молюски Арктики. // Природа. – 1987. - № 3. – С. 49 – 57.
Шабатура М.Н., Матяш Н.Ю., Мотузний В.О. Зоологія. 7 кл. — К.: Генеза, 1997.
Щербак Г.Й. Зоологія безхребетних. Кн. 1-3. – К.: Либідь, 1995.
Яновиг Л.М. Розмноження перлівницевих в умовах Центрального Полісся України. Спец.030008 – зоологія. Автореф. дис. канд. біол. наук. – К., 1998. – 16 с.
Додатки
Особливості будови різних представників класу Двостулкові
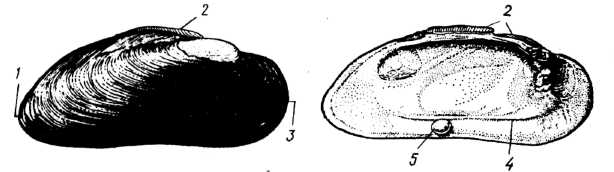
Рис. 16. Ряд Unionida: європейська річкова перлова скойка (Margaritifera margaritifera) з перлиною: 1 — задній кінець черепашки; 2 — зовнішній лігамент; 3 — передній кінець черепашки; 4 — мантійна лінія; 5 — перлина

Рис. 17. Ряд Mytilida: чорноморська устриця (Ostrea taurica): а - повзаюча личинка; б — молоді устриці, що приросли стулками до субстрату; в — черепашка дорослого молюска; 1 — черепашка; 2 — нога; 3 — лігамент; 4 — нижня (ліва) стулка, що приростає до субстрату; 5 — верхня (права) стулка з морськими жолудями на поверхні

Рис. 18. Ряд Mytilida: а — перлова скойка Pinctada margaritifera; б — Pinna sp.
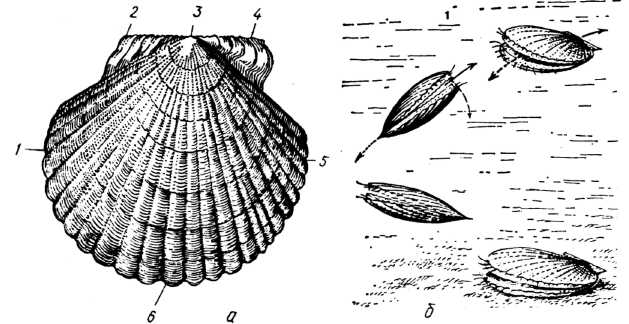
Рис. 19. Ряд Pectinida: приморський гребінець (Patinopecten yessoensis): а - права стулка ззовні; б — послідовні рухи гребінця; 1 — задній край черепашки; 2 — заднє вушко; 3 — маківка; 4 — переднє вушко; 5, 6 — передній та черевний краї черепашки

Рис. 20. Ряд Lucinida: а — горошинка (Pisidium amnicum); б — кулька рогова (Sphaerium rivicola)

Рис. 21. Ряд Venerida: тридакна (Тгіdacna сгосеа) — тіло орієнтовано так, як воно розташоване у ґрунті (черевною стороною догори)


Рис. 22. Ряд Venerida: a — їстівна серцевидка (Cerastoderma edule); б — морське серце (Glossus humanus); в — морський черенок (Solen vagina), стадії занурення в грунт; 1 — нога; 2 — вивідний сифон; 3 — ввідний сифон
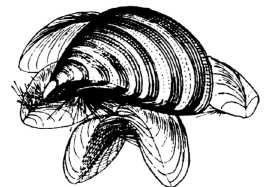
Рис. 23. Ряд Venenda: гроно прісноводних дрейсен (Dreissenа polymorpha), скріплених нитками бісусу
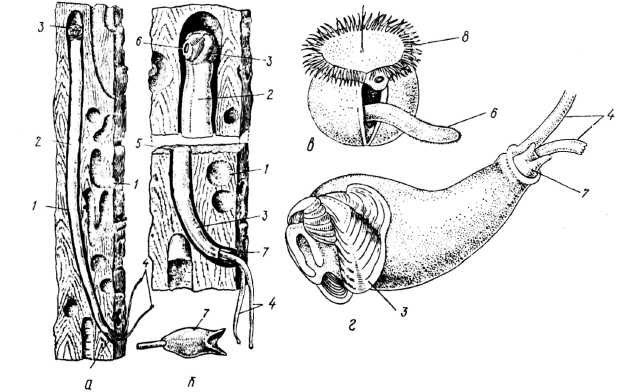
Рис. 24. Ряд Venerida: корабельний черв Teredo: а, б — дорослий молюск у проточених ним ходах; в — личинка велігер; г — молода стадія; 1 — ходи, які просвердлив тередо; 2 — тіло молюска; З — черепашка; 4 — сифони; 5 — вапнякова висилка ходів; 6 — нога; 7 — палетка; 8 — парус
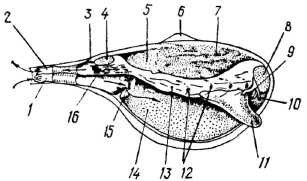
Рис. 25. Схема будови перетинчасто зябрового молюска Cuspidaria: 1 — ввідний сифон; 2 — вивідний сифон; 3 — анальний отвір; 4 — задній м'яз-замикач; 5 — верхня (надзяброва) камера мантійної порожнини; б — маківка; 7 — печінка; 8 — передній м'яз-замикач; 9 — рот; 10 — ротові лопаті; 11 — нога; 12 — пори в зябровій септі; 13 — зяброва септа; 14 — нижня (підзяброва) камера мантійної порожнини; 75 — перетинка; 16 — сифональний ретрактор