Кислотноосновное равновесие в крови пловцов при стандартной физической работе
Федеральное агентство по образованию
Пензенский государственный педагогический университет
им. В.Г. Белинского
Факультет
Естественно-географический
Кафедра
Биохимии
Дипломная работа
кислотно-основное равновесие в крови пловцов при стандартной физической работе
Студент
__________________ Миронов И.А
Руководитель
_________________ Петрушева О.П.
К защите допустить.
Протокол № от «____» ___________2008г.
Зав. кафедрой
__________________ Генгин М.Т.
Пенза, 2009г.
СОДЕРЖАНИЕ
ВВЕДЕНИЕ
ГЛАВА 1. ОБЗОР ЛИТЕРАТУРЫ
1.1. Понятие о кислотно-основном равновесии
1.2. Буферные системы организма
1.3. Клинико-биохимические показатели кислотно-основного равновесия организма
1.4. Нарушения кислотно-основного равновесия организма
1.5. Показатели кислотно-основного равновесия и уровня лактата в крови при физической нагрузке.
ГЛАВА 2. МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
2.1. Материалы исследования
2.2.Методы исследования
2.2.1. Метод определения парциального давления углекислого газа в капиллярной крови пловцов
2.2.2 Метод определения рН в капиллярной крови пловцов
2.2.3 Метод определения концентрации гидрокарбонатных ионов в капиллярной крови пловцов
2.2.4 Метод определения величины ВЕ в капиллярной крови пловцов
2.2.5. Метод определения концентрации лактата в капиллярной крови пловцов
ГЛАВА 3. РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
3.1. Показатели лактата в капиллярной крови пловцов до, во время физической нагрузки и в периоде раннего восстановления.
3.4. Показатели ВЕ в капиллярной крови пловцов до, во время физической нагрузки и в периоде раннего восстановления
ГЛАВА 4. ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
ВЫВОДЫ
Список литературы:
ВВЕДЕНИЕ
Одним из факторов, лимитирующим физическую работоспособность, является нарушение кислотно-основного состояния крови спортсменов. При выполнении мышечной работы наблюдается кислородный дефицит, что ведет к накоплению кислых метаболитов в организме. Все это способствует снижению рН в крови и тканях. Сдвиг кислотно-щелочного равновесия вызывает угнетение активности ферментов окислительно-восстановительных реакций, окисления углеводов и липидов, кроме того, меняется чувствительность клеточных рецепторов к медиаторам и гормонам и тд [19,42].
Организм всегда стремится к удержанию рН в строгих пределах. В поддержании кислотно-основного равновесия принимают участие буферные системы крови; внутриклеточный метаболизм, в процессе которого образуется и используется ион водорода; легкие, удаляющие углекислый газ; почки, реабсорбирующие бикарбонат и выделяющие ион водорода в форме NH4+ и H>3>PO>4. >В процессе систематических тренировок происходит накопление буферных резервов крови, совершенствуются механизмы легочной и почечной компенсации, что позволяет организму спортсмена более длительное удерживать рН на нормальном уровне при выполнении физической работы [1,4,6,7].
Целью данной работы было изучение механизмов адаптации кислотно-основного равновесия крови пловцов при физической работе.
При выполнении работы были поставлены следующие задачи:
Изучить показатели кислотно-основного состояния крови пловцов до физической работы.
Изучить показатели кислотно-основного состояния крови пловцов во время физической работы.
Изучить показатели кислотно-основного состояния крови пловцов в периоде раннего восстановления.
Научная новизна и практическая ценность работы.
Изучены показатели кислотно-основного состояния крови пловцов при стандартной работе, направленной на оценку гликолитической емкости. Полученные результаты могут быть основой для разработки методов профилактики и коррекции нарушений кислотно-основного равновесия организма, организации тренировочного процесса, направленного на увеличение буферных резервов крови спортсменов.
Апробация работы: Материалы дипломной работы доложены на научно-практической конференции студентов ПГПУ им. В.Г. Белинского.
ГЛАВА 1. ОБЗОР ЛИТЕРАТУРЫ
1.1 Понятие о кислотно-основном равновесии
Одним из важнейших условий жизнедеятельности организма является постоянство концентрации водородных ионов во внеклеточном пространстве и в клетках - кислотно-основное состояние (КОС) или кислотно-основное равновесие (КОР). Иными словами, КОР - относительное постоянство реакции внутренней среды организма, количественно характеризующееся концентрацией водородных ионов (протонов)[9,24,37]
Согласно определению Бренстеда, кислотами следует считать молекулы или ионы, способные отдавать ионы водорода (протоны), а основаниями - соединения, принимающие протоны. Таким образом, кислоты являются донорами ионов водорода, а основания - их акцепторами. Бикарбонатный анион (НСО>3>-), образующийся при диссоциации слабой угольной кислоты (Н>2>СО>3>), в физиологических условиях действует только как основание. Ранее основаниями назывались только щелочи типа NaОН, в связи, с чем и возник неточный термин "кислотно-щелочное равновесие". Однако, концентрации щелочей в биологических жидкостях ничтожно малы по сравнению с концентрацией анионов слабых кислот и физиологического значения не имеют, поэтому более правильно говорить о кислотно-основном равновесии (КОР)[18,29,41].
Концентрации водородных (Н+) и гидроксильных (ОН-) ионов в воде, степень диссоциации которой незначительна, одинаковы и составляют 10-7 моль/л. Для упрощения выражения этих концентраций введено понятие рН, которое соответствует отрицательному десятичному логарифму (- lg) концентрации водородных ионов. Так, для электрически нейтральной воды рН составляет 7,0, тогда как кислые растворы имеют рН менее 7,0, а щелочные - выше 7,0. Средняя нормальная концентрация водородных ионов в плазме крови составляет 40 нмоль/л, что соответствует рН 7,4[25,27]
От соотношения концентраций ионов водорода и ионов ОН- зависят интенсивность окислительно-восстановительных реакций, процессы расщепления и синтеза белков, окисления углеводов и липидов, чувствительность рецепторов к медиаторам и гормонам, проницаемость клеточных мембран и многое другое[21,23].
Большая часть ионов водорода образуется в результате распада углеводов, жиров, белков.
Другим важным продуктом обмена веществ является углекислый газ, который оказывает влияние на концентрацию ионов водорода. Углекислый газ называется кислотой, хотя к нему и не присоединен ион Н+, поскольку СО>2 >вступает в реакцию с водой с образованием углекислоты – Н>2>СО>3>. последняя диссоциирует с образованием протонов и является ведущим фактором закисления крови:
СО>2> + Н2О ↔ Н+ + НСО>3>- [32,33]
При постоянной температуре давление газа над поверхностью жидкости определяется количеством нерастворенного газа. Поскольку воздух представляет собой смесь газов, давление каждого газа пропорционально его молекулярной концентрации. Оно называется парциальным давлением.
При растворении углекислого газа в воде происходят два важных события: небольшое количество углекислого газа превращается в углекислоту – количество образуемой углекислоты прямо пропорционально количеству углекислого газа в растворе и, следовательно, парциальному давлению СО>2> – рСО>2>. Углекислота является слабой кислотой и лишь в незначительной степени диссоциирует на ионы водорода и бикарбоната.
Это при водит к тому, что повышение давления СО>2> в альвеолах увеличивает давление СО>2> в легочных капиллярах, а также количество углекислоты во внеклеточной жидкости и, наоборот.
Третьим фактором образования водородных ионов является соотношение окисленного и восстановленного гемоглобина:
2 Fe2+
+ 1/2O>2 >+ 2H+
2Fe3+ + H>2>O
[2,41,43,]
Fe2+
+ 1/2O>2 >+ 2H+
2Fe3+ + H>2>O
[2,41,43,]
1.2 Буферные системы организма
Постоянство рН внутренней среды организма обусловлено совместным действием буферных систем крови и тканей и ряда физиологических механизмов (деятельность легких и выделительная функция почек).
Буферными свойствами, т.е. способностью противодействовать изменениям рН раствора при внесении в него кислот или оснований, обладают смеси, состоящие из слабой кислоты и её соли с сильным с основанием или слабого основания с солью сильной кислоты. Буферная система в организме представляет собой сопряженную кислотно-основную пару, состоящую из донора и акцептора водородных ионов (протонов)[8,14].
Функционирование буферной пары описывается уравнением Гендерсона-Хассельбалха, которое связывает значение рН с константой диссоциации любой кислоты (К>А>):
>РН = рК>А> + lg [акцептор протонов]/[донор протонов] [28,30]
Важнейшими буферными системами крови являются: бикарбонатная, гемоглобиновая, белковая и фосфатная.
Бикарбонатная буферная система - достаточно мощная система внеклеточной жидкости и крови. На долю бикарбонатного буфера приходится 10% всей буферной ёмкости крови. Эта бикарбонатная система является сопряженной кислотно-основной парой, состоящей из молекулы угольной кислоты (Н>2>СО>3>), выполняющей роль донора протона, и бикарбонат-аниона (НСО>3>-) - основания, выполняющего роль акцептора протона. Пользуясь уравнением Гендерсона-Хассельбалха для данной буферной системы величину рН можно выразить через константу диссоциации угольной кислоты (рК Н>2>СО>3>) и десятичный логарифм соотношения концентраций ионов НСО3 - и недиссоциированных молекул Н>2>СО>3> : рН = рК Н>2>СО>3> + lg ([НСО>3>-]/[Н>2>СО>3>])[15,17,35].
При нормальном значении рН крови (7,4) концентрация ионов бикарбоната НСО>3>- в плазме примерно в 20 раз превышает концентрацию Н>2>СО>3> (или растворенного углекислого газа СО>2>).
Механизм действия данной системы заключается в том, что при выделении в кровь относительно больших количеств кислых продуктов водородные ионы (Н+) взаимодействуют с ионами бикарбоната (НСО>3>-) c образованием слабодиссоциирующей угольной кислоты Н>2>СО>3>. Снижение концентрации последней достигается ускоренным выведением СО>2> через легкие в результате их гипервентиляции. Когда же в крови увеличивается количество оснований, то они, взаимодействуя со слабой угольной кислотой, образуют ионы бикарбоната и воду; при этом заметных сдвигов в величине рН не происходит. К тому же, в сохранении нормального соотношения между компонентами бикарбонатной буферной системы участвуют физиологические механизмы регуляции КОР: происходит задержка в плазме крови некоторого количества СО>2> в результате уменьшения вентиляции (гиповентиляции) лёгких.
Второе место по значению после бикарбонатной буферной системы занимает буферная система гемоглобин-оксигемоглобин. Последняя играет важную роль в регуляции соотношения гемоглобин (слабое основание) - оксигемоглобин (слабая кислота), а также в преобразовании растворенной угольной кислоты в углекислый газ и выведении его через легкие. Функции этой системы зависят от концентрации гемоглобина в крови и от поступления достаточных количеств кислорода: при анемии и гипоксии её мощность резко снижается [13,39].
В эритроцитах имеется механизм сбережения оснований для организма, известный как эффект Амбурже. Он состоит в том, что образующийся в тканях углекислый газ превращается в эритроцитах в угольную кислоту (Н>2>СО>3>). В свою очередь Н>2>СО>3> диссоциирует на ион Н+ и анион НСО>3>- под влиянием фермента карбоангидразы (угольной ангидразы) эритроцитов. Ион водорода при этом захватывается буферными системами внутри клетки (гемоглобин, фосфаты), а анион бикарбоната возвращается в плазму крови, обмениваясь на анион хлора, поступающий в эритроцит (согласно равновесию Доннана). В эритроцитах анион хлора связывается с катионом калия. В лёгких образующийся оксигемоглобин связывает значительную часть калия, в результате чего анион хлора вытесняется за пределы эритроцита и связывается с катионом натрия, освобожденным при удалении углекислоты. В итоге происходит активное образование и задержка в организме аниона НСО>3>- (основания) и удаление угольной кислоты. [8,40,44]
Следует подчеркнуть, что степень связывания кислорода с гемоглобином существенно зависит от сдвигов рН плазмы крови: при сдвиге его в кислую сторону (ацидоз) сродство гемоглобина к кислороду снижается и соответственно уменьшается насыщение гемоглобина кислородом; при сдвиге рН в щелочную сторону (алкалоз) имеет место обратная зависимость: сродство гемоглобина к кислороду и насыщение его кислородом возрастают. Эта закономерность называется эффектом Бора.
Белковая буферная система имеет меньшее значение для поддержания КОР в плазме крови, чем другие буферные системы. Белки обладают буферными свойствами благодаря наличию кислотно-основных групп в молекуле белка: белок-Н+ (кислота, донор протонов) и белок - (сопряженное основание, акцептор протонов). Данная буферная система плазмы крови эффективна в области рН 7,2-7,4.
Фосфатная буферная система представляет собой сопряженную кислотно-основную пару, состоящую из однозамещенного и двузамещенного ортофосфата (NaН>2>РО>4> : Na>2>HPO>4>), в которой первый компонент является донором ионов Н+ (кислота), а второй - акцептором протонов (основание). Буферное действие фосфатной системы основано на возможности связывания водородных и гидроксильных ионов. Органические фосфаты также обладают буферными свойствами, но мощность их слабее, чем неорганического фосфатного буфера. Функционирование этой буферной системы тесно связано с участием почек в регуляции КОР. Ионы водорода активно секретируются в мочу канальцевым эпителием, причем этот процесс восстанавливает физиологические соотношения в фосфатной буферной системе и обеспечивает преобладание двузамещенного натрия в крови, оттекающей от почек. Выводимые таким путем избытки водородных ионов составляют титруемую кислотность мочи. Анионы сильных кислот выводятся вместе с катионом NH>4>+, который образуется из аммиака и водорода в почках. Этот процесс называется аммониогенезом и также направлен на удаление избытка ионов водорода. Почечная регуляция КОР, таким образом, включает образование и удаление ионов аммония, секрецию ионов водорода, а также экономию аниона бикарбоната (анионы бикарбоната из первичной мочи почти полностью абсорбируются в почечных канальцах)[10,13,20].
1.3 Клинико-биохимические показатели кислотно-основного равновесия организма
КОР оценивается на основании величины рН, парциального давления углекислого газа (рСО>2>), концентрации истинных (актуальных) стандартных бикарбонатов крови (SB), концентрации буферных оснований – ВВ,(от англ. Buffer base), избытка оснований в цельной крови – ВЕ (от англ. Bases excess).
рН крови (показатель концентрации водородных ионов). Границы этого показателя, совместимые с жизнью, следующие: от 6,8 до 7,8. В норме рН крови колеблется в узких пределах: от 7,35 до 7,45; рН ниже 7,35 указывает на значительное преобладание кислых продуктов обмена веществ (ацидоз); рН выше 7,45 свидетельствует о чрезмерном накоплении оснований (алкалоз). В других биологических жидкостях и в клетках рН может иметь иные значения: к примеру, в эритроцитах рН в норме составляет 7,19±0,02. Показатель водородных ионов отражает лишь общее направление сдвигов в КОР крови, тогда как тип этих расстройств (дыхательный или метаболический) можно оценить только на основании комплекса нижеследующих показателей.
Парциальное напряжение углекислого газа в крови (рСО>2>) составляет в норме в среднем 40 мм. рт. ст. (5,3 кПа). Повышение этого параметра свидетельствует о дыхательном ацидозе, связанном с угнетением дыхательной функции лёгких; это часто сочетается с компенсаторным подъемом бикарбонатов. Снижение рСО>2> (и нередко одновременное компенсаторное падение бикарбонатов) указывает на наличие дыхательного алкалоза, отмечающегося чаще всего при гипервентиляции легких.
Буферные основания (ВВ) - суммарный показатель содержания ионов бикарбоната и анионов белка и гемоглобина. Физиологические колебания данного показателя - от 31,8 до 65,0 ммоль/л. Его определение выявляет степень сдвига КОР за счёт метаболических изменений в тканях.
Basis excess (BE) - параметр, указывающий на избыток оснований (положительные значения) или их дефицит (отрицательные значения). В норме пределы колебаний этого показателя следующие: от 3,26±0,4 до -0,98±0,2 ммоль/л. Увеличение BE свидетельствует о развитии метаболического алкалоза, а уменьшение до отрицательных значений (вплоть до -20 ммоль/л) - о наличии метаболического ацидоза.
На практике ВЕ определяется по номограмме Сиггор-Андерсена.
Стандартные бикарбонаты (SВ) - концентрация бикарбонатов в плазме крови, уравновешенная при рСО>2> , равном 40 мм рт. ст., и при парциальном напряжении кислорода в крови, обеспечивающем полную насыщаемость гемоглобина кислородом.
Истинные или актуальные бикарбонаты (АВ) - показатель, соответствующий концентрации бикарбонатов в крови, взятой без соприкосновения с воздухом при температуре 38°С. У здоровых лиц содержание актуальных бикарбонатов и стандартных бикарбонатов (SB) примерно идентично и находится в пределах от 18,5 до 26,0 ммоль/л.
Определение концентрации как истинных, так и стандартных бикарбонатов, производиться с помощью номограмм, построенных по уравнению Гендельсона-Гассельбаха, наилучшая из которых является Сиггор-Андерсона[3,15,27].
1.4 Нарушения кислотно-основного равновесия организма
Несостоятельность компенсаторных механизмов организма в предотвращении сдвигов концентрации водородных ионов приводит к различным нарушениям КОР. В зависимости от механизмов развития этих нарушений различают дыхательный ацидоз или алкалоз и метаболический ацидоз или алкалоз.
Дыхательный ацидоз возникает вследствие гиповентиляции легких (при бронхиальной астме, пневмонии, при нарушениях кровообращения с застоем в малом круге, отёке лёгких, эмфиземе, ателектазе легких, угнетении дыхательного центра под влиянием ряда токсинов и препаратов типа морфина и т.п.). В результате наблюдается гиперкапния, т.е. повышение рСО>2> артериальной крови; при этом увеличивается содержание Н>2>СО>3> в плазме крови, что, в свою очередь, приводит к компенсаторному нарастанию ионов бикарбоната (НСО>3>-) в плазме. Одновременно со снижением рН крови при дыхательном ацидозе повышается выведение с мочой свободных и связанных (в форме аммонийных солей) кислот. [11,12]
Дыхательный алкалоз возникает при гипервентиляции лёгких (при вдыхании чистого кислорода, компенсаторной одышке, сопровождающей ряд заболеваний, в том числе нейротоксический синдром, инфекционно-вирусные состояния). При этом вследствие быстрого выведения из организма СО>2> развивается гипокапния, т.е. понижение рСО>2> в артериальной крови (менее 35 мм рт. ст.); снижение содержания угольной кислоты в артериальной крови сопровождается уменьшением бикарбонатов в плазме крови, т.е. снижается щелочной резерв крови, поскольку часть их компенсаторно превращается в угольную кислоту. Хотя этот механизм часто оказывается недостаточным, чтобы компенсировать уменьшение содержания Н>2>СО>3>. При дыхательном алкалозе отмечается снижение кислотности мочи и содержания в ней аммиака. [44,46]
Метаболический ацидоз - наиболее часто встречающаяся форма нарушений КОР, обусловлен накоплением в тканях и крови органических кислот. Он возникает при выполнении физической нагрузки из-за накопления лактата, сахарном диабете (увеличение кетоновых тел - бета-оксимасляной и ацетоуксусной кислот), нарушении питания, голодании, лихорадке, токсических состояниях, почечно-гломерулярной недостаточности, сердечно-сосудистой недостаточности, наследственной и приобретенной формах пиелонефрита, гипоальдостеронизме, адреногенитальном синдроме с потерей соли, заболеваниях желудочно-кишечного тракта, шоковых состояниях. При метаболическом ацидозе кислотность мочи и содержание в ней аммиака увеличены.
Метаболический алкалоз может развиться в результате потери большого количества кислотных эквивалентов (при неукротимой рвоте, желудочно-кишечных расстройствах) и усиленного поступления из желудочно-кишечного тракта веществ, не подвергшихся нейтрализации кислым желудочным соком и обладающих основными свойствами, а также при накоплении подобных агентов в тканях (в частности, при тетании) и в случаях избыточного и бесконтрольного введения щелочных растворов для коррекции метаболического ацидоза. При метаболическом алкалозе повышается содержание бикарбоната (НСО>3>-) в плазме и, следовательно, увеличивается щелочной резерв крови. Как компенсацию метаболического алкалоза следует рассматривать возникающую при этом гиперкапнию в результате снижения возбудимости дыхательного центра в условиях повышенного рН и, соответственно, урежения частоты дыхания. Данный тип нарушения КОР сопровождается снижением кислотности мочи и содержания в ней аммиака.
На практике изолированные формы дыхательных или метаболических нарушений КОР встречаются редко: чаще всего имеют место их сочетания. Так, смешанный ацидоз является результатом изменения как метаболических, так и дыхательных показателей; такие нарушения КОР нередко наблюдаются при бронхолегочной патологии.
Если при различных по характеру сдвигах КОР крови значения рН остаются в пределах нормы, такие изменения КОР можно считать компенсированными; если же величина рН выходит за границы нормы, тогда нарушения КОР могут быть либо частично компенсированными, либо некомпенсированными [38,40].
Таким образом, различные патологические состояния могут приводить к существенным сдвигам в кислотно-основном равновесии крови.
1.5 Показатели кислотно-основного равновесия и уровня лактата в крови при физической нагрузке
Динамические наблюдения за показателями КОС крови у спортсменов в условиях покоя отражают особенности их текущего функционального статуса, что дает возможность объективно оценить и контролировать процессы адаптации организма к мышечной деятельности [26].
Изучение особенностей КОС крови экспериментальных животных с разным уровнем их адаптированности к физическим нагрузкам было осуществлено в условиях предельной по продолжительности мышечной работы (изнуряющая плавательная нагрузка). Результаты исследований показывают прямую зависимость между степенью адаптированности животных к физической активности и величиной изменений внутренней среды организма. Это проявилась в том, что по мере повышения физической работоспособности животного, достигаемой систематической тренировкой плаванием, их способность выполнить предельную мышечную работу при более глубоко ацидотических сдвигах в крови увеличивается [22,35].
В процессе адаптации к мышечной работе происходит расширение диапазона гомеостаза деятельности. Однако способность тренированного организма к предельной мобилизации физиологических функций, а, следовательно, и способность совершать работу в значительно измененных условиях его внутреннего среды наблюдается лишь в тех случаях, когда предъявляемые при этом требования действительно максимальные. Во всех отдельных случаях действует основной защитный механизм саморегуляции – тенденция к меньшему отклонению от физиологического равновесия при наиболее целесообразной взаимокомпенсации функций и высокой эффективности тканевых процессов [27].
Содержание молочной кислоты в крови в норме в состоянии относительного покоя составляет 1-1,5 ммоль/л и существенно возрастает при выполнении интенсивной физической работы. При этом накопление ее в крови совпадает с усиленным образованием в мышцах, которое существенно повышается после напряженной кратковременной нагрузки и может достичь около 30ммоль*кг-1 массы при изнеможении. Количество молочной кислоты больше в венозной крови, чем в артериальной. С увеличением мощности нагрузки содержание её в крови может возрастать: у нетренированного человека до 5-6 ммоль/л, у тренированного – до 20ммоль/л и выше. В аэробной зоне физических нагрузок лактат составляет 2-4ммоль/л, в смешанной 4-10 ммоль/л, в анаэробной – более 10 ммоль/л. Условная граница анаэробного обмена соответствует уровню лактата 4 ммоль/л и обозначается, как порог анаэробного обмена (ПАНО), или лактатный порог (ЛП) [40,45].
Снижение лактата у одного и того же спортсмена при выполнении стандартного работы на разных этапах тренировочного процесса свидетельствует об улучшении тренированности, а повышение – об ухудшении [7,38].
Значительные концентрации молочной кислоты в крови после выполнения максимальной работы свидетельствуют о более высоком уровне тренированности при хорошем спортивном результате или большей метаболической емкости гликолиза, большей устойчивости его ферментативных систем к смещению рН в кислую сторону. Таким образом, изменение концентрации молочной кислоты в крови после выполнения определенной физической нагрузки связано с состоянием тренированности спортсмена.
Накопление молочной кислоты в крови при выполнении физической работы ведет к развитию метаболического ацидоза, который клинически характеризуется снижением рН. В спортивной практике важно определять показатели КОС при стандартной и предельной физической работе с целью оценки адаптивных возможностей организма спортсмена [1,2,3].
В последние годы исследователи уделяют все большее внимание особенностям функционально-метаболических изменений, возникающих в организме при мышечной деятельности в условиях произвольного апноэ. В испытаниях участвовали 106 спортсменов (мужчин) в возрасте 19—23 лет, имеющих I и II разряды по военно-прикладному плаванию (ныряние в длину). Забор крови из пальца руки проводили до и сразу после ныряния на дистанции 40—50 м. Продолжительность преодоления спортсменами этих отрезков под водой колебалась в пределах 38—58 с. Показатели КОС определялись по микрометоду Аструпа на аппаратах датского производства ABL-2 и OSM-2 Hemoximetr фирмы «Radiometr». Оценку кислотно-основного равновесия в организме осуществляли по истинному рН, истинному рСО>2>, концентрации стандартного бикарбоната (SB), избытку оснований (BE), содержанию гемоглобина (Нb). Показатели КОС до выполнения упражнений в нырянии были в пределах физиологических границ. Ныряние в длину вызвало большие сдвиги и показателях КОС крови, что выражалось в понижении показателя pH от 7,40 до 7,12; только у четырех спортсменов он соответствовал пределам физиологической нормы. После нагрузки дефицит оснований BE изменялся от -3 до – 15,8 ммоль/л. Уровень стандартного бикарбоната (SВ) снижался от 20,2 до 12,2 ммоль/л. Пять спортсменов имели показатель SB в пределах нормы. Показатель рСО>2> в крови после ныряния был в пределах нормы у 83 человек (4,7—6,0 кПа), понижен у 23 (до 3,07 кПа). Содержание гемоглобина у 48 ныряльщиков соответствовало норме (2,17 – 2,56 ммоль/л), 44 имели повышенный показатель (до 3,52 ммоль/л), у 14 он был снижен (до 1,39 ммоль/л) [30].
Полученные данные свидетельствуют, что ныряние в длину в условиях соревнований сопровождается весьма значительными сдвигами в кислотно-основном состоянии. Согласно литературным данным, интенсивная физическая работа под водой в условиях произвольного апноэ вызывает существенное накопление лактата в крови, указывающее на включение анаэробных механизмов в энергетическом обеспечении работающих мышц. Неизбежным результатом нейтрализации недоокисленных метаболитов анаэробного обмена является снижение содержания SB в крови. При этом отмечается дефицит оснований (показатель BE приобретает отрицательный знак) и наступает понижение показателя рН [30].
После ныряния, т. е. после работы в зоне субмаксимальной мощности, когда в энергообеспечении нагрузки превалирует анаэробный (гликолитический) путь ресинтеза АТФ, отмечались резко выраженные изменения содержания SB, BE и рН. Содержание дефицита оснований доходило до —15,8 ммоль/л, а величина рН достигала 7,12, что характерно для декомпенсированного метаболического ацидоза (рН<7,20). Очевидно, мощность буферных систем была недостаточна для того, чтобы обеспечить строгое постоянство КОС при интенсивной мышечной работе под водой в состоянии произвольного апноэ.
Согласно современным представлениям мощность буферных систем повышается при улучшении адаптации организма к мышечной деятельности под влиянием тренировки. Так, у малоподготовленных спортсменов снижение рН во время работы умеренной интенсивности более значительно чем у тренированных. Вместе с тем после нагрузок, возрастающих до пределов максимальной мощности, зависимость сдвига рН от уровня тренированности обнаруживается не всегда. Усиленная нейтрализация недоокисленных продуктов во время работы под водой, сопровождающейся острой гипоксией и гиперкапнией, является дополнительным источником продукции СО>2>. Однако выведение его из организма в первые 60-75 с (время, необходимое для забора крови) после ныряния, по-видимому, не затрудняет функцию дыхательного аппарата. Вероятно, этому способствует непроизвольная гипервентиляции, возникающая у спортсменов сразу после прохождения дистанции. У подавляющего большинства ныряльщиков он существенно не изменился. Повышение содержания гемоглобина у 44 спортсменов до 3,52 ммоль/л объясняется адаптационной реакцией крови на комбинированное воздействие гипоксии и гиперкапнии [30,36,38].
На основании полученных результат можно заключить, что интенсивная мышечная работа в условиях комплексная воздействия гипоксии и гиперкапнии (ныряние в длину) приводит к выраженным изменениям КОС крови, характерным для субкомпенсированного и декомпенсированного метаболического ацидоза. Несмотря на это, у подавляющего большинства обследуемых не выявлялось внешних признаков утомления и жалоб на состояние здоровья. Несомненно, это свидетельствовало об адекватности физического напряжения, вызываемого нырянием, функциональному состоянию и резервным возможностям организма спортсменов. Однако значительные биохимические изменения, происходящие в организме ныряльщиков во время соревнований, говорят о необходимости тщательного отбора спортсменов, их тренированности, введения соответствующих интервалов отдыха после ныряния [30,39].
Полученные данные могут быть использованы при планировании профилактических мероприятий по сохранению работоспособности военнослужащих при работе в аналогичных условиях и предупреждению несчастных случаев [30,31].
Таким образом, при выполнении физической работы отмечаются значительные сдвиги в кислотно-основном состоянии организма.
ГЛАВА 2. МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
2.1 Материалы исследования
Объектом исследования служила капиллярная кровь, которая забиралась в гепаринизированный капилляр в объеме 200 мкл. В полученных образцах определяли рН, рСО>2>, уровень НСО>3>-, ВЕ, концентрацию лактата.
Экспериментальную группу составили пловцы с квалификацией: мастера спорта и мастера спорта международного класса. Спортсмены выполняли тест на гликолитическую емкость: 4x50 м в режиме 1'30" - 1'15" - 1'. Показатели кислотно-основного равновесия крови были изучены до, во время физической работы и в периоде раннего восстановления.
2.2 Методы исследования
Показатели кислотно-щелочного равновесия определяли на Анализаторе критических состояний ROSH OMNI S. Анализ занимает всего несколько мин после взятия пробы крови. Величины рН, рСО>2>, стандартные и истинные бикарбонаты, избыток буферных оснований и концентрация лактата определяются одновременно.
2.2.1 Метод определения парциального давления углекислого газа в капиллярной крови пловцов
Парциальное давление углекислого газа в крови определяли с помощью потенциометрических микроэлектродов [26].
2.2.2 Метод определения рН в капиллярной крови пловцов
рН в крови определяли потенциометрическим методом с помощью рН-метра.
2.2.3 Метод определения концентрации гидрокарбонатных ионов в капиллярной крови пловцов
Величину НСО>3>- в крови вычисляли по результатам измерения величин рН и рСО>2>, для чего использовали следующее уравнение:
сНСО>3>- = 0.0307рСО>2>*10 (рН-6,105)> >[ 16,26,]
2.2.4 Метод определения величины ВЕ в капиллярной крови пловцов
Избыток и дефицит буферных в крови оснований определяли с помощью измеренных параметров с использованием следующего уравнения:
BE= (1-0.014ctHb)*((1.43ctHb+7.7)*(pH-7.4)-24.8+cHCO3-)) [3,16,26,34]
2.2.5 Метод определения концентрации лактата в капиллярной крови пловцов
Величину лактата в крови определяли с помощью ферментного электрода с иммобилизованной лактатдегидрогеназой [26,34].
ГЛАВА 3. РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
3.1 Показатели лактата в капиллярной крови пловцов до, во время физической нагрузки и в периоде раннего восстановления
Результаты исследования показывают, что содержание лактата в капиллярной крови пловцов до физической работы находилось в пределах физиологической нормы.
При выполнении тестирующей нагрузки концентрация молочной кислоты в крови спортсменов существенно нарастала, что связано с преобладанием гликолитического пути ресинтеза АТФ.
В периоде раннего восстановления обнаружено возвращение уровня лактата к исходным значениям, так как происходило устранение кислородного дефицита, возникшего при выполнении физической нагрузки (рис. 1).
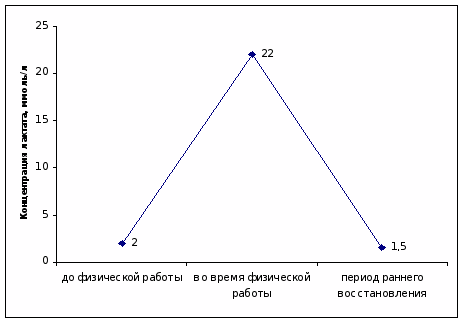
Рисунок 1. Концентрация лактата в капиллярной крови пловцов до, во время физической нагрузки и в периоде раннего восстановления, ммоль/л.
3.2 Показатели парциального давления углекислого газа в капиллярной крови пловцов до, во время физической нагрузки и в периоде раннего восстановления
На дорабочем уровне в капиллярной крови пловцов отмечаются высокие значения парциального давления углекислого газа в силу того, что имеет место гиповентиляция легких.
При выполнении теста усиливается дыхательная функция легких, что выражается в снижении рСО>2.>
В периоде раннего восстановления происходит устранение кислородного дефицита в организме, поэтому незначительная гипервентиляция легких сохраняется (рис. 2).

Рисунок 2. Уровень парциального давления углекислого газа в капиллярной крови пловцов до, во время физической нагрузки и в периоде раннего восстановления, мм рт ст.
3.3 Показатели рН в капиллярной крови пловцов до, во время физической нагрузки и в периоде раннего восстановления
До физической работы в капиллярной крови пловцов обнаруживаются физиологические значения рН.
При физической нагрузке происходит закисление крови и тканей кислыми продуктами обмена, преимущественно лактатом, поэтому отмечается значительное снижение рН.
Однако, после теста рН крови приходит в норму, так как аэробная работа способствует устранению избытка кислот в организме (рис. 3).
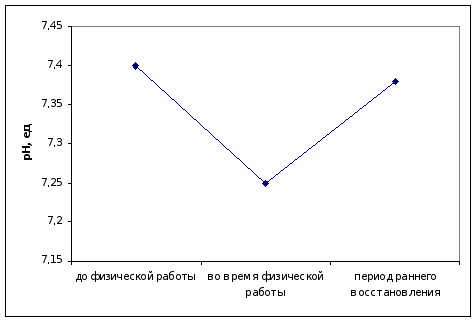
Рисунок 3. Уровень рН в капиллярной крови пловцов до, во время физической нагрузки и в периоде раннего восстановления, ед.
3.4 Показатели ВЕ в капиллярной крови пловцов до, во время физической нагрузки и в периоде раннего восстановления
На дорабочем уровне в капиллярной крови пловцов отмечался избыток буферных оснований – ВЕ, – что указывает на высокое развитие щелочных резервов.
При выполнении физической работы выявлен существенный дефицит ВЕ, так как возникла необходимость в поддержании гоместаза в условиях закисления.
После нагрузки в крови спортсменов происходило устранение недостатка буферных оснований, однако небольшой дефицит ВЕ еще сохранялся (рис. 4).
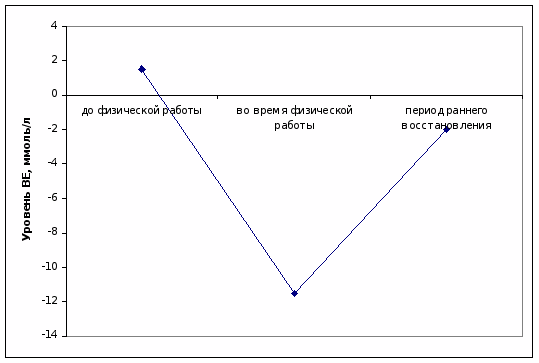
Рисунок 4. Уровень ВЕ в капиллярной крови пловцов до, во время физической нагрузки и в периоде раннего восстановления, ммоль/л.
3.5 Показатели гидрокарбонатных ионов в капиллярной крови пловцов до, во время физической нагрузки и в периоде раннего восстановления
До физической работы в капиллярной крови спортсменов выявлено незначительное повышение содержания гидрокарбонатных ионов по сравнению с физиологической нормой, что также указывает на высокий уровень развития щелочных резервов организма.
При выполнении теста в крови пловцов обнаруживается снижение уровня НСО>3>-. Вероятно, это связано с вовлечением в компенсацию ацидоза карбонатных буферных систем крови.
В периоде раннего восстановления обнаруживается возвращение содержания НСО>3>- к исходным значениям (рис. 5).

Рисунок 5. Уровень гидрокарбонатных ионов в капиллярной крови пловцов до, во время физической нагрузки и в периоде раннего восстановления, ммоль/л.
Таким образом, при выполнении физической нагрузки в анаэробных условиях происходят значительные сдвиги показателей кислотно-основного равновесия в крови спортсменов.
ГЛАВА 4. ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
В практике спорта большое значение имеет жесткий контроль за параметрами кислотно-основного равновесия крови. Нарушения КОС ведут к значительным изменениям во всех видах обмена веществ. Физическая работа, особенно анаэробного характера, всегда сопровождается нарастанием кислородного дефицита в организме. В таких условиях большое значение имеет гликолитический путь ресинтеза АТФ, конечным продуктом которого является молочная кислота. Лактат выбрасывается из мышц в кровь, вызывая сдвиг рН в кислую сторону. Это, в свою очередь, может способствовать снижению активности ферментов энергообразования, нарушению проведения гормонального сигнала, преобладанию катаболических процессов. В результате возникает угроза срыва процессов адаптации к физической работе [7,32].
При систематическом занятии спортом в организме отмечаются существенные биохимические изменения, что позволяет им развивать положительные компенсаторные реакции при выполнении физической работы. Такие сдвиги происходят и в кислотно-основном равновесии тканей и крови [7].
Регулярные тренировки способствуют повышению буферных резервов организма, что является необходимым условием для поддержания рН при закислении. При недостаточном развитии щелочной емкости крови, механизмов почечной и легочной компенсации может происходить более раннее прекращение работы, связанное, прежде всего, со снижением интенсивности энергообразующих процессов. Поэтому является актуальным исследование механизмов адаптации кислотно-основного равновесия организма спортсменов при физической нагрузке [32].
Результаты исследования показали, что до физической работы уровень лактата в капиллярной крови спортсменов соответствует физиологической норме. Нарушений со стороны кислотно-основного состояния не выявлено. Следует отметить имеющуюся гиповентиляцию легких, что выражается в высоких значениях парциального давления углекислого газа. На фоне этого, вероятно, происходит нарастание буферных резервов крови, так как отмечается избыток ВЕ и гидрокарбонатных ионов. Возможно, на дорабочем уровне у спортсменов отмечается респираторный ацидоз, компенсированный метаболическим алкалозом. Такое исходное состояние должно позволить лучше переносить предстоящее закисление организма при выполнении физической работы.
При выполнении тестирующей нагрузки в капиллярной крови спортсменов обнаружено существенное увеличение уровня лактата, что указывает на высокую гликолитическую емкость организма, выражающуюся в достаточных запасах гликогена в мышцах и печени, а также в адекватной работе ферментов анаэробных реакций.
Однако, накопление молочной кислоты привело к значительному закислению крови, т.е. при выполнении физической нагрузки развился метаболический лактат-ацидоз. На этом фоне наблюдаются компенсаторные реакции со стороны буферных резервов крови, что выражается в снижении уровня ВЕ и концентрации гидрокарбонатных ионов. Кроме того, при выполнении теста в капиллярной крови пловцов отмечается снижение парциального давления углекислого газа. Это указывает на подключение легочного механизма компенсации нарушений КОС, выражающееся в гипервентиляции.
Таким образом, при выполнении физической работы в организме спортсменов развивается метаболический ацидоз, который частично компенсируется респираторным алкалозом и буферными резервами крови, что позволяет более длительное время поддерживать механизмы энергообразования и сохранять работоспособность. Кроме того, на высокий уровень развития щелочных резервов крови указывает высокое соотношение концентрации лактата к уровню ВЕ. Оценка адекватности компенсаторного ответа организма на метаболический ацидоз произведена при сопоставлении значений лактата и рН, т.е . на фоне высоких значений молочной кислоты не происходит существенного падения рН. В связи с этим следует отметить высокий уровень тренированности пловцов.
В периоде раннего восстановления анаэробная физическая работа сменялась на аэробную. На таком фоне в организме спортсменов происходило устранение кислородного дефицита, возникшего в условиях выполнения физической нагрузки. В капиллярной крови пловцов обнаружено возвращение уровня лактата к физиологическим нормам. При активном потреблении кислорода происходит полное сгорание молочной кислоты и других продуктов обмена до воды и углекислого газа с целью восполнения запасов АТФ. Кроме того, имеющаяся легочная гипервентиляция позволяет восполнять кислородный дефицит и выводить излишки углекислого газа, образующегося при активном окислении кислых метаболитов.
Таким образом, в аэробных условиях в периоде раннего восстановления происходит устранение метаболического ацидоза, развившегося при тестировании. В результате в норму приходят такие показатели кислотно-основного равновесия, как рН, концентрация гидрокарбонатных ионов, уровень ВЕ, хотя небольшой дефицит буферных оснований имеет место.
Необходимо отметить, что возвращение показателей кислотно-основного равновесия крови пловцов к физиологическим нормам происходило довольно быстро, что указывает на высокий уровень тренированности спортсменов. Такой фон облегчает дальнейшее восстановление организма и подготавливает к следующей нагрузке. Устранение кислых метаболитов способствует созданию адекватных условий для синтетических процессов, которые являются основой для закрепления результатов тренировок, повышения квалификации и достижения высоких спортивных результатов[32,33]. Напротив, недостаточное удаление лактата из крови спортсменов в периоде раннего восстановления может приводить к дополнительному напряжению буферных систем организма, что, в свою очередь, вызывает срыв адаптационных реакций с угрозой развития утомления [11,27,39,40]. В результате спортсмен может не восстановиться к следующей тренировке.
Таким образом, при выполнении физической работы анаэробного характера в организме отмечаются существенные компенсаторные сдвиги в КОР, которые позволяют длительное время сохранять работоспособность. Однако, следует очень внимательно относиться к биохимическим изменениям к периоде раннего восстановления с целью недопущения развития перетренированности и утомления, что может отрицательно сказаться на функциональном состоянии организма. Полученные результаты позволяют своевременно диагностировать нарушения КОС в организме, что облегчает дальнейшую коррекцию подобных сдвигов. Кроме того, полученные данные могут быть использованы для построения тренировочного процесса с целью развития буферных резервов крови.
ВЫВОДЫ
В организме пловцов до физической работы отмечается высокий уровень развития буферных резервов крови и механизмов легочной компенсации, что определяет готовность к выполнению физической работы.
При физической нагрузке в организме пловцов развивается метаболический лактат-ацидоз, который частично компенсируется буферными основаниями крови и респираторным алкалозом.
В периоде раннего восстановления наблюдается интенсивный процесс окисления лактата и быстрое возвращение к норме показателей буферной системы крови.
Список литературы:
Агапов Ю.Я. Кислотно-щелочной баланс, М. изд. Медицина :1983
Березов Т.Т., Коровкин Б.Ф. Биологическая химия. – М: «Медицина», 2004
Биохимические методы исследования в клинике. Под ред. Меньшикова В.В. Элиста: АПП. Джангар:1998,.
Введение в клиническую биохимию. Под ред. И.И. Иванова. Л.: 1969
Владимиров Г.Е. и Пантелеева Н.С. Функциональная биохимия. Изд. Ленинградского университета, 1965 ССР, 1953
Биохимия заболеваний мышц. Под ред. Фердмена Д.Л. Изд. А.Н.Украинской.
Волков Н.И., Несен Э.Н., Осипенко А.А., Корсун С.Н. Биохимия мышечной деятельности. – Киев: Олимпийская литература, 2000
Гомеостаз. Под ред. Горизонтова М.:1976
Гортейши Я. Основы клинической биохимии. Прага, 1967,
Граевская Н.Д. В сб: Современная система и методы врачебного контроля в спорте. – Малаховка МОГИФК, 1997,
Давиденко Д. Н. Функциональные резервы адаптации организма спортсмена /Д.Н. Давиденко, А.С. Мозжухин. – Л., 1995
Дементьева И.И. Лабораторная диагностика и клиническая оценка нарушений гомеостаза у больных в критическом состоянии при хирургических вмешательствах и в отделении интенсивной терапии. – М: ЗАО «Рош-Москва», 2007
Калинин В.М. Проблема гомеостаза в спорте: кислотно-основное состояние крови при адаптации к мышечной деятельности. Кемерово, 2009
Капланский С.Я. Кислотно-щелочное равновесие в организме и его регуляция М. – л.: 1940
Клинико-лабораторные аналитические технологии и оборудование. под ред. И.И. Меньшикова, М.:2007г
Клиническая биохимия. под ред. В.А.Ткачука, М.ГЕОТАР-МЕД. 2004
Крохалов А.А. Водный и электролитный обмен. М:1972
Лазарис Я.А. и Серебровская И.А. Нарушение кислотно-щелочного гомеостаза Л.:1973
Маршалл В.Дж. Клиническая биохимия. Пер. с англ., М.-СПб.: ©Бином»- «Невский диалект», 1999,
Меерсон Ф. З. Адаптация к стрессорным ситуациям и физическим нагрузкам //Ф.З. Меерсон, М.Г. Пшенникова. – М.: Медицина, 1988.
Методы практической биохимии. М.:1978,
Озолина Е.В. Ключевые аспекты адаптации организма инвалидов к физическим нагрузкам / Озолина Е.В., Дмитриев В.С. // Открытый мир : Науч.-практ. семинар по адапт. двигат. активности. - М., б. г. - С. 58-72
Робинсон Дт. Р. Основы регуляции кислотно-щелочного равновесия, пер. с англ. М:1969
Руководство по клинической реаниматологии. Под ред. Т.М.Дербянина, с 73, М.: 1974
Рут Г. Кислотно-щелочное состояние и электролитный баланс, пер. с англ. М.:1978
Эндрю Сейнт Джон. Экспресс-диагностика. Roche Diagnostics, Маннхайм.
Справочник по функциональной диагностике. Под ред. И.А.Кассирского, М.:1970
Физиология дыхания. Под ред. Л.Л. Шиха,. Л.:1973 с 256
Хашен Р., Шейх Д. Очерки по патологической биохимии. М.: 1981,
Чернец М.И., Потапов А.В. Влияние ныряния в длину с задержкой дыхания на кислотно-основное состояние крови // Военно-медицинский журнал. – 1998. - № 8. – с. 53-54.
Ютикова О.С., Бурлакова А.Ю. Использование биохимических критериев контроля соревновательной деятельности пловцов в процессе многолетней подготовки. //Успехи современного естествознания 2008 №9,
Яковлев Н.Н. Биохимия спорта М.: ФиС,1974
Яковлев Н.Н. Очерки по биохимии спорта. Изд. физкультура и спорт», 1953
Antony P. F. Turner .Biosensor; Fundamentals and Applications. 1997.
Antonutto, G. and Prampero, P. "The Concept of Lactate Threshold"// Journal of Sports Medicine and Physical Fitness, March 1995.vol. 35,
Arthur PG, West TG, Brill RW, Schulte PM, and Hochachka PW. Recovery metabolism in tuna white muscle: rapid and parallel changes of lactate and phosphocreatine after exercise.// Can J Zool -1992-№70: р1230–1239,
Baggott J., PhD, Dennis Sh.E., MS. Medical Biochemistry. 2003.
Brooks GA. Intra- and extra-cellular lactate shuttles. // Medicine and Science in Sports and Exercise (2000)№32, р 790-799
Hochachka P.W. and Somero GN. Biochemical Adaptation—Mechanism and Process in Physiological Evolution. New York: Oxford University Press, 2001.
Hochachka P.W., C. L. Beatty, Y. Burelle, M. E. Trump, D. C. McKenzie аnd G. O. Matheson The Lactate Paradox in Human High-Altitude. //Physiological Performance News in Physiological Sciences, Vol. 17, No. 3, 122-126, June 2002
Hood and R. L. Tannen: Protection of acid-base balance by pH regulation acid production //New England Journal of Medicine. (1998) №32 р343
C. C. W. Hsia: Respiratory function of hemoglobin. //New England Journal of Medicine (1998) №338, р 239.
Jones JH. Optimization of the mammalian respiratory system: symmorphosis versus single species adaptation. //Comp Biochem Physiol B Biochem Mol Biol №120B-1998: 125–138,
Kleshnev I.V., Suhova O.A., Petriaev A.V. Dynamics of structure of special endurance of a high class swimmers. // Sport Science ’99 in Europe. 4th Annual Congress of European College of Sport Science. Rome University Institute of Motor Sciences, Rome, July 1999,
Lundby C, Saltin B, and van Hall G. The ‘lactate paradox’, evidence for a transient change in the course of acclimatization to severe hypoxia in lowlanders.// Acta Physiol Scand 2000 №170: р 265–269.
Riley R.L., Dutton R.E. Regulation of respiration and blood gases. Ann.N.Y. Acad., 1963, Vol 109,