Состояние естественной резистентности и иммунологической реактивности у новорожденных телят при колибактериозе
Государственный Аграрный Университет Молдовы
На правах рукописи
ЖOCAH Николай Степанович
УДК 619:619. 98-092:636.2-053.2
СОСТОЯНИЕ ЕСТЕСТВЕННОЙ РЕЗИСТЕНТНОСТИ И ИММУНОЛОГИЧЕСКОЙ РЕАКТИВНОСТИ У НОВОРОЖДЕННЫХ ТЕЛЯТ ПРИ КОЛИБАКТЕРИОЗЕ
16.00.03 – ветеринарная микробиология и вирусология
Диссертация
на соискание ученной степени доктора ветеринарных наук
Научный консультант – Скутару Иван Гаврилович, доктор-хабилитат ветеринарных наук, профессор
Кишинев 1998
Содержание:
Список условных сокращений……………………………6
ВВЕДЕНИЕ 7
РАЗДЕЛ I ОБЗОР ЛИТЕРАТУРЫ 17
1.1. Краткие сведения о колибактериозе телят 17
1.2. Специфическая иммунологическая реактивность телят в постнатальный период 43
1.3. Неспецифическая иммунологическая реактивность новорожденных телят 53
1.4. Получение и применение лактоглобулинов 57
заключение 64
РАЗДЕЛ 2 РЕЗУЛЬТАТЫ СОБСТВЕННЫХ ИССЛЕДОВАНИЙ 66
2.1. Материалы и методы исследований. 66
2.2. Изучение специфической иммунологической реактивности телят в постнатальный период. 89
2.2.1. Уровень иммуноглобулинов в сыворотке крови телят до приема молозива и через 24ч после рождения и взаимосвязь между концентрацией иммуноглобулинов и проявлением синдрома нарушения пищеварения. 90
2.2.2. Количественная характеристика абсорбции колостральных иммуноглобулинов новорожденными телятами. 94
2.2.3. Уровень колостральных иммуноглобулинов в сыворотке крови новорожденных телят через 6, 24 и 48 ч после рождения. 100
2.2.4. Количественное определение колостральных иммуноглобулинов в сыворотке крови и в фецес неонатальных телят.. 102
2.2.5. Концентрация иммуноглобулинов в сыворотке крови и секрете молочной железы коров. 103
2.2.6. Содержание иммуноглобулинов в сыворотке крови коров и новорожденных телят. 105
2.2.7. Уровень антител против О-антигенов Е. coli в нормальном коровьем молозиве, молоке и сыворотке крови новорожденных телят.. 110
2.2.8. Уровень антител против К-антигенов Е. coli в нормальном коровьем молозиве, молоке и в сыворотке крови новорожденных телят. 120
2.2.9. Протективный механизм молозива при колибактериозе новорожденных телят. 131
2.2.10. Факторы, влияющие на выживание новорожденных телят. 134
2.2.11. Клиническая оценка эффективности вакцинации коров для профилактики колибактериоза телят. 137
ВЫВОДЫ 145
РАЗДЕЛ 3 ИЗУЧЕНИЕ НЕСПЕЦИФИЧЕСКОЙ ИММУНОЛОГИЧЕСКОЙ РЕАКТИВНОСТИ НОВОРОЖДЕННЫХ ТЕЛяТ 147
3.1. Биохимический анализ крови новорожденных телят. 147
3.2. Коррекция кислотно-основного баланса при диареи неонатальных телят. 154
3.3. Изменение фагоцитарныхных показателей у новорожденных телят в зависимости от клинического статуса. 158
3.4. Бактерицидная активность сыворотки крови новорожденных телят. 171
3.5. Комплементарная активность сыворотки крови новорожденных телят 178
3.6. Лизоцимная активность сыворотки крови. 184
3.7. Пропердиновая активность сыворотки крови. 189
ВЫВОДЫ 193
РАЗДЕЛ 4 ЭФФЕКТИВНОСТЬ ЛЕКАРСТВЕННЫХ ПРЕПАРАТОВ ПРИ КОЛИБАКТЕРИОЗЕ НОВОРОЖДЕННЫХ ТЕЛЯТ 194
4.1. Эффективность применения колостральной сыворотки и лактоглобулина для коррекции иммунодефицита неонатальных телят. 194
4.2. Эффективность лактоглобулина и ретинола применяемых с целью коррекции иммунодефицита новорожденных телят. 198
4.3. Применение хлорпромазина и Т-активина для лечения телят, больных колибактериозом. 203
ВЫВОДЫ 209
5. Обсуждение результатов исследований. 210
6. ВЫВОДЫ. 222
7. РЕКОМЕНДАЦИИ ПРОИЗВОДСТВУ. 228
С ПИСОК
условных сокращений
ПИСОК
условных сокращений
агт – агглютинационный тест
ант – антиглобулиновый тест
атф – аденозинтрифосфат
днк – дезоксирибонуклеиновая кислота
мпа – мясо-пептонный агар
мпб – мясо-пептонный бульон
мппб – мясо-пептонный печоночный бульон
ра – реакция аглютинации
рнк – рибонуклеиновая кислота
рн – отрицательный логарифм концентрации водородных ионов
сда – синдром дефицита антител
tl – термолабильная фракция E. coli
ts – термостабильная фракция E. coli
фэк-м – фотоэлектрокаллориметр
цамф – циклический 3’5’ - аденозинмонофосфат
В ВЕДЕНИЕ
ВЕДЕНИЕ
Успешное развитие животноводства во многом зависит от направленного выращивания молодняка, сочетающего высокую продуктивность с устойчивостью организма к заболеваниям.
Результаты многочисленных исследований состояния естественной резистентности организма сельскохозяйственных животных свидетельствует о том, что защитные силы являются динамичным показателем, и определяется как генетическими особенностями организма, так и воздействием различных факторов окружающей среды. Это обстоятельство позволяет направленно влиять на формирование и проявление защитных сил организма. Обеспечение животным благоприятных условий содержания, максимально отвечающих биологическим особенностям организма, сложившимся в процессе эволюционного развития, способствует более быстрому формированию и лучшему проявлению его защитных сил. Вместе с тем, неблагоприятное воздействие окружающей среды приводит к ослаблению устойчивости организма, защитные силы его проявляются недостаточно, что усиливает опасность возникновения и распространения инфекционных заболеваний. Следовательно, инфекционные болезни могут возникнуть только в результате нарушения нормальной реактивности, ослабления защитных свойств организма.
На фоне нарушения нормальной реактивности и ослабления защитных свойств организма возникают массовые желудочно-кишечные заболевания новорожденных животных, особенно телят. Среди них, колибактериоз телят относится к числу наиболее широко распространенных инфекционных болезней молодняка и регистрируется во всех развитых странах мира, в том числе в хозяйствах Республики Молдова.
Вследствие этого, актуальность изучения данного заболевания стала не меньшей, а наоборот возросла. Это связано с резистентностью энтеротоксических штаммов Е. coli к большинству доступных химиотерапевтических средств. Данная резистентность имеет внехромосомальный характер, она детерминируется плазмидами (R) и в связи с этим постепенно охватывает все большее число штаммов энтеротоксических и определяется, как «заразная комплексная резистентность к антибиотикам». Кроме того, продуцируемые эшерихиями экзо- эндо- и энтеротоксин способны воздействовать самостоятельно, что затрудняет использование средств специфической защиты и профилактики. Очень сложное многогранное и далеко не изученное воздействие возбудителя на иммунную систему организма хозяина приводящее к крайне низкой его сопротивляемости.
Биологическая активность энтеротоксинов проявляется в виде нарушения равновесия между процессами секреции электролитов в кишечнике. Колонизация энтеротоксических штаммов Е. coli посредством специфических фимбрий в энтероциты, дает возможность проникновения в них молекул энтеротоксинов, которые нарушают состояние равновесия между объемами электролитов, перемещаемых из кровеносных сосудов в кишечник и в обратном направлении, это же равновесие является результатом роста секреции жидкостей эпителием кишечных крипт и торможения их резорбции клетками эпителия кишечных ворсинок.
Таким образом, вследствие продуцирования кишечной палочкой (Е. coli) энтеротоксина происходит стимуляция аденилаткиназы энтероцитов, что вызывает трансформацию АТФ в циклический 3’,5’-аденозинмонофосфат (АМФ). Одновременно изменяется функция энтероцитов, которые вместо абсорбции питательных веществ, элиминируют свободную воду и минеральные вещества из клетки в кишечник, что приводит к резкому обезвоживанию и деструктивным изменениям ворсинок эпителия [E. Salajka, 1980]. Следовательно, колонизация слизистой кишечника патогенными Е. coli имеет основное значение для развития болезни. Смертельные исходы в ходе острой формы колибактериоза у новорожденных телят являются следствием существенных нарушений электролитного равновесия и дегидратации.
Актуальность темы. Снижение заболеваемости и предупреждение гибели народившегося молодняка является одной из главных задач, стоящих перед ветеринарной наукой и практикой.
Одной из главных причин, тормозящих полное сохранение нарождающегося молодняка — массовые желудочно-кишечные заболевания новорожденных животных, особенно телят. Эти заболевания имеют широкое распространение, наносят значительный экономический ущерб и вызывают большой отход телят, который составляет в странах мира от 3,1 до 31,0 %, а в бывшем СССР средний отход среди заболевших телят за ряд лет составил 18-21 % [Г. В. Гнатенко, 1968; Л. К. Волынец, 1975; А. В. Драгомир, Е. А. Драгомир, A.Ф. Карышева, Ф. В. Спатарь, 1985; А. А. Гутковский, 1989; Ю. С. Кабанков, С. Ф. Армашу, 1991].
Среди заболеваний новорожденных телят колибактериоз занимает одно из ведущих мест. Он относится к числу острых заболеваний, чаще всего протекающее с признаками диареи, интоксикации, септицемии, расстройства сердечно-сосудистой и центральной нервной системы. В возникновении, развитии и исходе заболевания реактивность организма играет первостепенную роль [С. Н. Преображенский, Н. И. Блинов, Н. В. Ершова, Г. В. Вышинский, 1974; И. М. Архангельский, 1976; В. С. Бузлама, С. М. Сулейманов, В. Н. Долгополов, Н. Н. Коновалов, М. Н. Рецкий, И. С. Толкачев, 1978; П. А. Емельяненко, 1979; Э. С. Коган, 1981; В. Я. Мозгис, 1982; В. М. Асламов, 1983; М. А. Костына, 1984; R. Morar, 1985; А. Н. Голиков, 1989; И. М. Карпуть, А. Г. Ульянов, 1990; Е. А. Маринин, Т. Т. Ворошилова, 1993; П. П. Игнатьев, Г. Ч. Бондаренко, 1994; T. E. Besser, C. C. Gay, 1994; A. S. Sheikh, P. S. Bradford, S. C. James, B. Patricia, B. F. Thomas, H. Richard, D. Gregore, D. R. Lin, W. Bert, 1995; Д. Н. Масюк, 1997].
Защитные силы животных являются основными показателями, обеспечивающими устойчивость организма новорожденных телят к факторам внешней среды и их тестирование позволяет определять иммунный статус.
Неблагоприятное воздействие окружающей среды приводит к ослаблению устойчивости организма и усиливает опасность возникновения и распространения различных заболеваний, в том числе инфекционных. В связи с этим академик А. А. Богомолов отмечал, что инфекционные болезни могут возникнуть только в результате нарушений нормальной реактивности, ослабления защитных свойств организма [С. И. Плященко, В. Т. Сидоров, А. Ф. Трофимов, 1990].
Несмотря на то, что в профилактике колибактериоза телят достигнуты определенные успехи, однако эта проблема еще далеко от своего полного решения. Лекарственные препараты, используемые с этой целью, не вызывают коррекцию иммунного статуса. Кроме того, реализация существующих способов профилактики колибактериоза телят затруднена из за отсутствия экспресс-индикации К-антител в колостральной сыворотке коров.
Изучение иммунологической реактивности организма новорожденных телят, таким образом, приобретает актуальное значение для понимания патогенеза заболевания, для рациональной патогенетической терапии.
Связь работы с научными программами. Тема диссертационной работы была частью проблемы ветеринарной медицины Республики Молдова «Разработка методов и средств профилактики и борьбы с болезнями животных».
Цель работы. Цель работы состоит в экспериментальном и теоретическом обосновании неспецифической и специфической реактивности новорожденных телят при колибактериозе, разработке иммунологически обоснованных рациональных способов коррекции иммунного статуса и ингибирования циклического аденозинмонофосфата (сАМР) посредством «анти-секреторного фактора».
Для реализации данной цели мы поставили следующие задачи:
Определить уровень антител против О- и К-антигенов Е. coli в нормальном коровьем молозиве, молоке и в сыворотке крови новорожденных телят.
Изучить количественную характеристику абсорбции колостральных иммуноглобулинов классов G, М и А новорожденными телятами.
Выявить роль кислотно-основного баланса в течение инфекционного процесса при колибактериоза телят.
Установить величину взаимосвязи между показателями неспецифической реактивности; (фагоцитарная реакция, бактерицидная, лизоцимная, комплементарная и пропердиновая активность) у здоровых, больных и павших новорожденных телят.
Разработать способы коррекции иммунного статуса новорожденных телят и ингибирования циклического аденозинмонофосфата (сАМР) посредством «антисекреторного фактора».
Научная новизна полученных результатов.
Новизна работы заключается в том, что впервые к изучению заболевания колибактериоза телят проявлен новый, комплексный подход. В результате комплексно проведенных исследований изучена специфическая и неспецифическая реактивность организма новорожденных телят.
Впервые изучен уровень антител против К-антигенов в нормальном коровьем молозиве, молоке и в сыворотке крови новорожденных телят.
Доказано, что клинический статус новорожденных телят находится в прямой зависимости от сывороточной концентрации иммунных глобулинов и их катаболической эффективности, увеличение уровня иммуноглобулинов М и А в сыворотке крови клинически здоровых животных в среднем в 5,7 и 3,4 раза, по сравнению с павшими указывает на их защитную функцию. Выявлено нарушение кислотно-щелочного равновесия при диареи неонатальных телят. Доказано, что сопутствующими ацидозу были отклонения в уровнях сывороточных хлоридов натрия, кальция и калия. Применение электролитного раствора корректировало указанные аномалии.
Установлена взаимосвязь между фагоцитарными показателями и бактерицидной активностью сыворотки крови, между фагоцитарной активностью и уровнем пропердина, между уровнем пропердина и лизоцимной активностью, между уровнем комплемента и фагоцитарной активностью у здоровых, больных и павших телят.
Обоснованы и испытаны лекарственные препараты, корректирующие иммунный статус новорожденных телят и ингибирующие циклический аденозинмонофосфат (сАМР) посредством «анти-секреторного фактора».
Практическая значимость полученных результатов. На основании результатов исследований определен уровень антител против О- и К-антигенов Е. coli. Изучена количественная характеристика абсорбции колостральных иммуноглобулинов новорожденными телятами.
Материалы дают информацию о содержании иммуноглобулинов G, M и А в сыворотке крови и в фекалиях телят больных колибактериозом, а также об экспресс методах для определения иммунного статуса новорожденных телят.
Для коррекции иммунного статуса испытан лактоглобулин совместно с ретинолом. Испытан хлорпромазин совместно с Т-активином для ингибирования циклического аденозинмонофосфата.
Результаты исследований позволяют дать теоретически обоснованные рекомендации по коррекции специфической реактивности организма новорожденных телят, для понимания патогенеза заболевания и для рациональной патогенетической терапии, так как в возникновении, развитии и исходе заболевания реактивность организма играет первостепенную роль.
Основные результаты исследований используются в учебной и научно-исследовательской работе и в практической деятельности ветеринарных специалистов хозяйств Республики Молдова.
Для ветеринарной наук и практики предложены:
Методические рекомендации по применению колостральной сыворотки и лактоглобулина для коррекции иммунодефицита неонатальных телят. Одобрены научно-техническим советом Министерства сельского хозяйства и продовольствия Республики Молдова. Изданы: Молдагроинформреклама, Кишинев, 1992.
Методические рекомендации – Экспресс-метод индикации антиген-агглютининов против К-антигена Е. coli в колостральной сыворотке коров и в сыворотке крови новорожденных телят. Одобрены Научно-техническим советом Министерства сельского хозяйства и продовольствия республики Молдова, Кишинев, 1994.
Разработаны способы:
Коррекция иммунодефицита неонатальных телят. (Brevet de invenţie, n 345), Republica Moldova.
Лечение телят больных колибактериозом (Brevet de invenţie, nr. 409), Republica Moldova.
Результаты исследований включены в тексты лекций и реализуются на лабораторно-практических занятиях со студентами ветеринарного и зооинженерного факультетов Государственного аграрного университета Республики Молдова.
Личный вклад соискателя. Автором самостоятельно выполнен весь объем экспериментальных исследований. Биохимический анализ крови проводили на венгерской лабораторной установке «Контифло».
Апробация результатов диссертации. Результаты научных исследований доложены и одобрены на состоявшемся в октябре 1972 г. в г. Москве Пленуме отделения ветеринарии ВАСХНИЛ по проблеме лечения и профилактики болезней молодняка сельскохозяйственных животных; на научной конференции профессорско-преподавательского состава Одесского сельскохозяйственного института (Одесса, 1974); на научной конференции по вопросам повышения продуктивности сельскохозяйственных животных в условиях Северного Казахстана (Целиноград, 1976), на Республиканской научной конференций профессорско-преподавательского состава и аспирантов Кишиневского сельскохозяйственного института им. М. В. Фрунзе (Кишинев 1977); на 3-х конференциях профессорско-преподавательского состава научно-исследовательской конференции Кишиневского сельскохозяйственного института (Кишинев, 1979, 1980, 1982); на Республиканской научно-производственной конференции (Кишинев, 1981); на Республиканской научно-практической конференции «Технические проблемы продовольственной программы» (Кишинев, 1984); на Всесоюзной научно-производственной конференции «Вопросы интенсификации и научно-обоснованного ветеринарного обслуживания промышленного животноводства» (Кишинев, 1987); на заседаниях Ученого совета ветеринарного факультета Государственного аграрного университета Республики Молдова. (1988-1998 гг.).
Публикации. По материалам диссертации опубликовано 37 работ в трудах ВАСХНИЛ, межвузовских сборниках и республиканских изданиях, в том числе 2 учебника (учебные пособия), 1 монография, 29 научных статей, 2 патента, 1 рацпредложение, 2 методические рекомендации.
Структура и объем диссертации. Диссертация изложена на 229 страницах машинописного текста и состоит из введения, обзора литературы, результатов собственных исследований, обсуждения полученных результатов, выводов, рекомендаций производству, списка литературы и приложения. Работа содержит 88 таблиц и 11 рисунков. Список литературы включает 410 источников, в том числе 212 иностранных.
РАЗДЕЛ I ОБЗОР ЛИТЕРАТУРЫ
1.1. Краткие сведения о колибактериозе телят
1.1.1. Историческая справка. Учение о роли в патологии новорожденных телят кишечных палочек (Bact. сoli, позже названной Eschеrichia coli, 1939), обнаруженных Обихом [C. О. Jensen, 1897], а позже Эшерихом [Th. Escherich, 1885] прошло длительную историю становления.
Морские свинки и кролики после заражения кишечной палочкой погибали через несколько часов, а иногда через 2 дня, при наличии сильного поноса и повышенной температуры [Th. Escherich, 1885]. Болезнетворное действие микроба проявлялось как при интраперитонеальном, так и при подкожном заражении.
Датский ученый C. O. Jensen, 1897 первый обратил внимание на кишечную палочку как на возбудителя поносов у новорожденных животных. Его исследования позволили предположить об аналогичной причине диспепсических расстройств у детей.
По данным многочисленных исследователей среди массовых желудочно-кишечных заболеваний новорожденных телят значительный удельный вес занимают инфекционные заболевания, из которых наибольшее распространение имеет коли-инфекция [П. Алтухов, 1904; М. К. Апалев, 1908; Th. Smith, M. L. Orcutt, 1925; Ф. Гутира, И. Марек, Р. Маннингер, И. Мочи, 1961; Т. П. Руденко, 1963; О. С. Андреева, 1966; Д. Келмен, 1967; Е. П. Чиркова, 1968; Я. Е. Коляков, С. С. Гительсон, Л. С. Каврук, 1970; Л. Н. Головнев, 1970; S. Craven, 1970; C. C. Gaу, 1971; H. Feу, 1972; T. Gossling, K. A. Mckaу, 1974; H. W. Moon, 1974; S. D. Acres, C. J. Laing, O. M. Radostis, 1975; А. В. Голиков, А. С. Вовк, А. Т. Марчук, В. Т. Ширяева, 1976; J. Tyler, 1976; Л. К. Волынец, 1978; Г. Пашкявичус, 1978; Я. Е. Коляков, 1978; Ю. В. Алексеев, 1979; J. E. C. Bellamу, S. D. Acres, 1979; J. Hutchinoson, 1979; H. W. Smith, M. B. Huggins, 1979; E. Salajka, 1980; М. А. Сидоров, 1981; H. Feу, 1981; Дж. Х. Б. Рой, 1982; C. C. Gaу, S. M. Parish, T. C. McGuire, 1982; В. А. Аликаев, В. В. Матюшин, 1983; Г. В. Гнатенко, Л. Т. Тупица, 1984; М. А. Сидоров, 1984; В. А. Гушул, Л. А. Афанасьев, Т. П. Мантрова, 1984; M. E. Amstutz, 1985; J. A. Morris, W. J. Sojka, 1985; В. Г. Зароза, 1985; И. И. Фельдман, 1985; Я. Антал, Р. Благо, Я. Булла, 1986; В. И. Девликамов, М. А. Сидоров, 1986; L. Okerman, 1987; M. M. Levine, 1987; К. С. Ливанов, 1988; Г. Л. Дворкин, 1989; З. М. Бедоева, 1990; В. Г. Зароза, 1991; В. А. Ушкалов, 1992; Г. К. Волков, В. Д. Баранников, 1997; ].
1.1.2. Возбудитель болезни. Изучение энтеропатогенной роли кишечной палочки для человека и животных начинается сразу же после открытия ее Обихом Цит. по Jensen C. O., (1897), позже Эшерихом [Th. Escherich, 1885] и, особенно Иенсеном [C. O. Jensen, 1897].
В 1897 году C. O. Jensen, 1897 отметил, что в органах и кишечнике телят, павших от кровавого поноса, находится большое количество микробов подобных тем, которые были открыты Эшерихом. Иенсен установил, что достаточно новорожденному теленку дать вместе с молоком 5-6 мл бульонной культуры Е. coli, выделенной из трупов телят, павших от поноса, чтобы вызвать смертельную болезнь.
Возможность эндогенного возникновения колибациллеза у телят путем скармливания им креолина, пиоктанина и формалина, вызывавших у телят острый энтерит и последующее проникновение Е. coli в общий кроваток экспериментально подтвердил C. O. Jensen, 1905. Из органов павших в этих случаях животных выделяли бактерии кишечной палочки, способные вызвать заболевание при пероральной даче их здоровым телятам без введения раздражающих веществ. На этом основании Иенсен считал, что дизентерию, «белый понос», может вызвать любая кишечная палочка, если по каким либо причинам резко снижается резистентность организма новорожденного. Иенсен первый приготовил вначале моно валентную сыворотку а затем поливалентную гипериммунную колисыворотку и с успехом ее применил для профилактики «белого поноса».
Мнение Иенсена разделяли и русские исследователи [П. Алтухов, 1904; М. К. Апалев, 1908].
Иенсен указывал, что гибель новорожденных телят, может быть как в результате септической формы колибациллеза, так и в результате тяжелого гастроэнтерита с явлениями интоксикации, без проникновения возбудителя в кровяное русло. Автор не обнаружил различия в культурально-биохимических свойствах у штаммов, выделенных от поносящих телят, и штаммов, выделанных от здоровых животных того же возраста.
Количественные и качественные соотношение разных серогрупп Е. coli в содержимом кишечника мало чем различаются у больных и здоровых телят [W. J. Sojka, 1971].
В настоящее время для дифференциации энтеропатогенных штаммов и непатогенных используют классический тест на изолированных петлях кишечника кролика, поросенка, теленка [E. Salajka, 1980].
В последнее время опубликовано значительное количество работ, в которых подробно дается характеристика купьтурально-морфологических и биохимических свойств Е. coli выделенных от новорожденных телят, как больных, так и здоровых [C. O. Jensen, 1897; О. С. Андреева, 1966; Е. П. Чиркова, 1968; А. Ф. Пилуй, В. А. Ленькова, В. В. Медведьев, 1974; L. L. Myers, P. A. Guinee, 1976; Я. Е. Коляков, 1978; Л. К. Волынец, 1978; J. Hutchinson, 1979; М. А. Сидоров, 1980; S. Tzipori, 1981; H. Brade, C. Galanos, 1983; И. И. Тарасов, 1984; Б. М. Анохин, 1985; А. В. Драгомир., Е. А. Драгомир., А. Ф. Карышева, Ф. В. Спатарь,1985; И. И. Фельдман, 1985; В. Тилга, Н. Кампус, 1986; У. Риихикоски, 1986; H. W. Ewing, 1986; М. А. Сидоров, 1987; Н. А. Соколова, О. А. Полякова, Н. И. Евглевская, 1987; К. С. Ливанов, 1988; I. Blanco, E. A. Gonzalez, S. Garcia, 1988; Н. Н. Жосан, 1989; А. А. Гутковский, 1989; B. H. Janke, D. H. Francis, J. E. Collins, 1990; В. Г. Зароза, 1991; Ю. С. Кабанков, С. Ф. Армашу, 1991; G. Galiero, M. Palladino, O. Lai, C. G. Goffredi, 1995; А. В. Куликовский, А. Н. Панин, В. В. Соснина, 1997].
Согласно последним международным классификациям кишечных бактерий (9-ое издание определителя бактерий Берги) род Escherichia представлен одним видом Escherichia coli, который в свою очередь делится на серо-ферментативные типы.
Е. coli представляет собой бактерию или кокобактерию размером 1-4 мкм/0,4-0,6 мкм. Под влиянием внешних факторов эшерихии трансформируются в нитевидные формы длиной до 8-10 мкм. Спор не образуют. Перитрихи, но некоторые штаммы не имеют жгутиков (атрихи). Под воздействием антибиотиков и других факторов образуются гранулярные формы (L-форма), представленные сферическими элементами различной величины (5-20 мкм). Большинство серотипов, изолированных от здоровых животных не образуют капсулу. Изолированные серотипы от больных животных могут быть как капсулообразующими (серотипы 08, 09 и 0101), так и не образующими капсулу.
Красятся всеми анилиновыми красками. Грам-негативные.
Е. coli обладают ресничками (пили), которые представляют рецепторы, посредством которых происходит абсорбция на бактериальной клетке РНК-содержащих фагов, а также осуществляется проникновение РНК фага в микробную клетку. В процессе конъюгации через реснички (пили), выполняющие роль коньюгационного мостика (канала) происходит передача ДНК от клетки донора к клетке реципиенту. Фимбрии (реснички, пили) обладают адгезивными свойствами с помощью которых эшерихии прикрепляются на эпителиальных клетках пищеварительного тракта.
Способность Е. coli образовывать фимбриальные адгезины определяют в капельной РА с моноспецифическими антиадгезивными сыворотками К99, F41, Аtt25, 987Р, К88ав, К88ас, К88аd [О.А. Полякова, Н. И. Евглевская, Н. А. Соколова, 1986; Ю. А. Маллахов, О. А. Тугаринов, М. К. Пирожков, Т. И. Исхакова, 1993].
Е. coli аэроб или факультативный анаэроб. Оптимальная температура роста 37,5° С, рост и размножение бактерий может происходить в пределах 15-45° С, оптимум рН 7,0, но культивировать возможно и при значительных изменениях рН среды.
Культивируют, как на обычных (МПБ, МПА), так и на дифференцнально-днагностических средах (Эндо, Левина). В МПБ рост эшерихий характеризуется равномерным помутнением среды без осадка или с образованием незначительного осадка легко разбивающегося при встряхивании. В старых культурах образуется иногда пристеночное кольцо и очень редко пленка. Форму «R» бактерий продуцируют культуры образующие, осадок, а МПБ остается почти прозрачным.
На МПА форма «S» бактерий образуют округлые колонии, слегка выпуклые, непрозрачные с ровными краями. Форма «R» представлена сухими колониями, сплющенными, плотно прилегающими к среде с неровными краями. Размер колоний варьирует в пределах 2-10 мм.
На среде Эндо кишечная палочка растет в виде малиново-красных колоний с металлическим блеском или без него, а на среде Левина (агар с эозином и метиленовой синькой) в виде темно-фиолетовых или черных колоний.
Кишечная палочка не требовательна к питательной среде и способна размножаться даже в изотоническом растворе хлорида натрия [Я. Е. Коляков, С. С. Гительсон, Л. С. Каврук, 1990].
1.1.3. Биохимические свойства. Одним из ведущих дифференциальных признаков кишечной палочки, отличающих ее от бактерий других родов семейства Enterobacteracea является сбраживание лактозы с образованием кислота и газа [И. В. Голубева, 1985].
Кишечная палочка способна ферментировать многие сахара и многоатомные спирты, глюкозу, маннит, дульцит, сахарозу, арабинозу и др., но, как правило, не изменяет адонита и инозита. При сбраживании углеводов образуется пируват, превращающийся в молочную, уксусную и муравьиную кислоты. Глюкоза, лактоза, маннит сбраживаются с образованием кислоты и газа. Сахароза и дульцит ферментируются непостоянно.
Кишечная палочка не усваивает цитраты, не разжижает желатину, выделяет индол и не образует сероводород, дает положительную реакцию с метилротом и отрицательную Фогес-Проскауэра (не образует ацетилметилкарбинол), редуцирует нитраты в нитриты, непостоянно декарбоксилирует лизин и орнитин. Она не обладает фенилаланиндезаминазой и цитохромоксидазой, являющейся окислительным ферментом. Не растет на средах с цианистым калием [H. Fey, 1972; А. А. Гутковский, Г. Л. Дворкин, 1989; В. Г. Зароза, 1991; Ю. С. Кабанков, С. Ф. Армашу, 1991].
1.1.4. Антигенная структура. Дифференциация патогенных Е. coli от непатогенных стало возможным благодаря детальному изучению антигенной структуры бактерий рода Escherihia [Th. Smith, M. L. Orcutt, 1925; H. Knipschildt, 1945; F. Kaufman, 1966; H. Fey, 1972; G. Orskov, F. Orskov, 1984; А. А. Гутковский, Г. Л. Дворкин, 1989; Ю. С. Кабанков, С. Ф. Армашу, 1991].
В 1948 году была опубликована первая классификация Кауфмана-Книпшильдта-Вэлна, которая послужила основанием к международной классификации. [H. Fey, 1972].
Е. coli имеет сложную антигенную структуру. Различают соматический О-антиген, поверхностный (оболочечный) К-антиген и жгутиковый Н-антиген [H. Knipschildt, 1945; F. Kaufman, 1966; H. Fey, 1972; G. Orskov, F. Orskov, 1984].
В настоящее время по О-антигену эшерихии подразделяются на 180 серогрупп, по К-антигену на 104 и по Н-антигену на 56 серогрупп. Установлено 18 фимбриальных адгезинов (пили-антигены) [В.Г.Зароза, 1988, 1991].
Соматический О-антиген Е. coli термостабильный, резистентный к нагреванию при 100° С в течении 2,5 ч. Он представляет собой полисахариднопротеинолипидный комплекс.
К-антиген имеет полисахаридную природу и состоит из трех компонентов, обозначенных буквами L, В, А. Для типизации культуры по О-антигену ее прогревают при температуре от 100 до 121°С. L- и В-антигены обладают сходной термостабильностью. Антигенная способность штамма, обладающего В-антигеном не полностью разрушается при нагревании при 100° С в течение 2,5 ч. Более строгая дифференцнация между L- и В-антигенами происходит, когда культуру нагревают при 121°С. А-антиген сохраняет агглютинабильные и антигенные свойства после нагревания культуры при 100° С, но эти свойства утрачиваются при 121° С, через 2,5 ч. [Я. Е. Коляков, С. С. Гительсон, Л. С. Каврук, 1970; H. Fey, 1972; А. А. Гутковский, Г. Л. Дворкин, 1989; В. Г. Зароза, 1991]
Штамм Е. coli может обладать более чем одним типом К-антигена [G. Orskov, F. Orskov, 1984].
Жгутиковый Н-антиген термолабильный, обладает антигенными и агглютиногенными свойствами при нагревании до 60°С, эти свойства утрачиваются при 100° С. Химический состав антигена-Н белковой природы [Th. Smith, M. L. Orcutt, 1925].
Адгезивные антигены Е. coli имеют первостепенное, значение, поскольку они позволяют микроорганизмам конкурировать с бактериями коменсалами при паразитировании в организме животных.
Известно 7 типов специфических адгезивных антигенов: К99, К88, 987Р, СFА1, СFА2 и F7, Аtt25 [В. Г. Зароза, 1988; А. Н. Головко, 1997].
Адгезивный антиген К99 впервые описан в 1975 г. он характерен для кишечных палочек, энтеропатогенных для телят и ягнят, по некоторым сообщениям и для поросят, жеребят, козлят [F. Orskov, G. Orskov, H. W. Smith, W. J. Sojka, 1975; S. Tzipori, 1981].
В адгезивном антигене К99 в зависимости от иммунофоретической активности его фракций различают анионный и катионный компоненты. Первый содержится только в серотипах Е. coli О9, О101, а второй в серотипах Е. coli О8, О9, О20, О64, О101. Серотипы Е. coli с анионным компонентом вызывает агглютинацию эритроцитов овец, морских свинок и лошадей, а с катионным компонентом только эритроцитов лошадей.
При заболевании телят диареей Е. coli К99 обычно заселяют медиальный и каудальный сегменты тонкого кишечника. Энтеротоксигенные Е. coli с этим антигеном обычно вызывают диарею у 1-2-дневных телят и 1-4-дневных ягнят [H. W. Moon, S. C. Whipp, S. M. Skartvedt, 1976].
Адгезивный антиген К88 впервые описан в 1961г. По иммунофоретической подвижности адгезивный антиген К88 имеет три варианта: К88ав, К88ас и К88ад. [C. J. Murray, 1987].
Е. coli с адгезивным антигеном К88 вызывает в 71% случаев диарею у новорожденных поросят. [J. A. Morris, W. J. Sojka, 1985].
Адгезивные антигены 987Р и F41 впервые описаны в 1977г. и в настоящее время признаны энтеротоксигенными для поросят [B. Nagy, Gy. Nagy, 1982]. Характерная особенность культур Е. coli с адгезивпым антигеном 987Р отсутствие способности агглютинировать эритроциты. Этот антиген не сочетается с К88, обладает более выраженными, чем К88 колонизирующими и адгезивными свойствами и встречается иногда у телят. Адгезивный антиген F41 в отдельных случаях у Е. coli может сочетаться с адгезивным антигеном К99 [J. A. Morris, W. J. Sojka, 1985].
Адгезивные антигены СFА1, СFА2, и F7 впервые описаны в 1975 (СFА1, СFА2) и в 1980 F7 .Все эти антигены характерны для кишечных палочек, энтеротоксигенных для человека. Данные типы адгезивных антигенов термолабильны, обладают выраженной специфичностью, агглютинируют у человека эритроциты крови группы А [S. Tzipori, 1981].
Адгезивные антигены Е. coli закодированы в плазмидах, которые относятся к молекулам ДНК (независимо от хромосомы бактериальной клетки) и обладают способностью к репликации. При скрещивании бактерий-доноров с другими реципиентами плазмиды легко им передаются т.е. обладают трансмиссивностью [М. А. Сидоров, М. А. Соколова, 1982; J. A. Morris, W. J. Sojka, 1985].
Взаимосвязь адгезивных и соматических антигенов. При диареи телят энтеротоксигенные Е. coli с адгезивным антигеном К99 в 47 % случаев относятся к О-группе 101, а в остальных случаях к О-группам 64, 8, 9, 20. При холероподобных заболеваниях детей у энтеротоксигенных Е. coli адгезивный антиген СFА1 коррелирует с соматическим антигеном 78, а антиген СFА2 с О-группами 6 и 8 [B. Nagy, Gy. Nagy, 1982]. При диареи поросят энтеротоксигенные Е. coli с адгезивным антигенами К88, 987Р, К99 присутствуют у Е. coli О-групп 9, 20, 141, 101, 149, причем Е. coli О101 чаще всего синтезируют антиген К99 [M. Awad-Masalmeh, H. W. Moon, P. L. Runnels, R. A. Schneider, 1982].
1.1.5. Токсины. Несмотря на многочисленные исследования в области изучения биологии кишечной палочки, все еще недостаточно ясно, за счет каких именно свойств патогенных серотипов Е. coli обусловливается их энтеропатогенность и способность, в отличие от непатогенных кишечных палочек, вызывать заболевания у новорожденных телят [Ю. П. Вертиев, 1987].
Поэтому изучение токсических компонентов Е. coli имеет большое значение в расшифровке патогенности кишечной палочки.
Уже вскоре после открытия кишечной палочки отдельные исследователи отличали у данного микроба наличие токсических свойств [P. H. Romer, H. Much, 1906].
На основании наблюдений при заражении культурой кишечной палочки, установлено наличие у данного микроба двух типов токсических веществ: 1) вызывающих токсикоз и 2) обусловливающих энтерит [А. И. Улендеев, В. И. Оленин, О. Г. Тихонова, 1971; И. В. Голубева, 1985; Ю. В. Езепчук, 1985].
Впервые присутствие токсина в фильтратах бульонных культур кишечной палочки было отмечено А. Г. Радзинским [Д. Э. Беленький, 1932]; Его наблюдения в дальнейшем подтвердил Д. М. L. Barber, 1978, который установил, что кишечная палочка продуцирует два токсина экзотоксин, появляющиеся в бульонной культуре уже в течение первых суток роста и эндотоксин, накапливающийся в тех же культурах позднее, в результате аутолиза микробных тел.
Токсичность суточных фильтратов бульонных культур исчезла после прогревания их при 56°С в течение 30 мин [Д. Э. Беленький, 1932]. Прогревание же «старых» фильтратов (из 20-40 суточных культур) при 56°С не приводило к потере их токсичности.
Более детальное изучение экзотоксинов Е. coli проводилось И. В. Голубевой, 1985. Автор исследовала экзотоксины в фильтратах бульонных культур 130 штаммов Е. coli. Ею были установлены различия в антигенных и иммуногенных свойствах экзо- и эндотоксинов. Величина смертельной дозы термолабильного колитоксина зависела от силы токсина каждого штамма, индивидуальной чувствительности животных и способа введения токсина. Наиболее характерные патологоанатомические изменения, вызываемые экзотоксином, автор обнаружила в нервных клетках спинного мозга.
Наличие у Е. coli экзо- и эндотоксинов было подтверждено многочисленными учеными: [В. Л. Елин, 1957; Т. П. Руденко, 1963; О. С. Андреева, 1966; K. Garson, E. Bull, 1970; А. И. Улендеев, 1971; H. Fey, 1971b; C. Wray, J. R. Thomlinson, 1972; H. Fey, 1972; J. W. Boyd, J. R. Baker, A. Leyland, 1974;Л. К. Волынец, 1975; E. T. Anderson, L. S. Young, W. L. Hewitt, 1978; Дж. Х. Б. Рой, 1982; J. Blanu, J. H. Parvu, O. Ivanciu, 1983; W. H. Ewing, 1986; А. А. Гутковский, Г. Л. Дворкин, 1989; Л. Сланина, 1989; В. А. Ушкалов, 1992].
Эндотоксины Е. coli представлены полисахаридо-липидо-протеиновым комплексом, ассоциирующимся с О-антигеном, который интегрирован в кишечную стенку и сому. Это основной токсин общий для патогенных и апатогенных грам-негативных микроорганизмов. Эндотоксиновый комплекс может быть экстрагирован из бактерий в количестве 5-10 % в который входят: полисахариды 45-60 %, липид А 5-15 %, протеин 15-20 % и липиды 10% [Д. Э. Беленький, 1932; C. Wray, J. R. Thomlinson, 1972].
Эндотоксины обладает важным биологическим эффектом в течение инфекционного процесса, вызванного грам-негативными микроорганизмами, иммунотерапия и иммунопрофилактика в данном случае являются чрезвычайно необходимы [C. Wray, J. R. Thomlinson, 1972; I. Kim, D. W. Watson, 1978; D. C. Morrison, R. J. Ulevitch, 1978; E. Neter, 1983]. Но защита против эндотоксина не возможна О-антителами, что достигается защитой против экзотоксинов, продуцируемых грам-позитивными микроорганизмами [H. Fey, 1972].
Антисыворотка против эндотоксинов, полученная при О-антигенной стимуляции обладает нейтрализующим антитоксическим эффектом по отношению к данному штамму [S. Lariviere, R. Lallier, M. Morin, 1979]. Несмотря на этот обнадеживающий факт, эндотоксины грам-негативных бактерий не могут быть нейтрализованы сходным образом как токсины клостридий. В связи с этим антиэндотоксическая сыворотка не обладает терапевтическим и профилактическим действием по сравнению с дифтерийным токсином [H. Fey, 1971а; H. Fey, 1971b;].
Энтеротоксин Е. coli представляет собой токсический комплекс, продуцируемый внеклеточно бактериальной клеткой. Энтеротокоин стимулирует возникновение диареи, вследствие чего нарушает водно-минеральный статус. Основным методом, позволяющим идентифицировать штаммы продуцирующие энтеротоксин является метод «кишечной лигатуры» [И. В. Голубева, 1985; M. Decun, 1986; А. А. Гутковский, Г. Л. Дворкин, 1989]. Энтеротоксигенные эшерихии пролиферируют в кишечник ворсинки и вызывают большое накопление полостной жидкости и диарею.
Энтеротоксины Е. coli состоят из двух фракций:
Фракция термостабильная (ТS) резистентная к нагреванию при 100° С в течение 30 мин.
Фракция термолабильная (TL) резистентная к нагреванию при 60° С в течение 10 мин.
Установлено тесное антигенное сходство между термолабильным энтеротоксином Е. coli и энтеротоксином Vibrio cholerae [W. J. Sojka, 1971; E. Salajka, 1980].
Термостабильный энтеротоксин Е. coli способен диализироваться, резистентный к воздействию кислот, трипсина, не обладает антигенными (иммунными) свойствами. Энтеротоксин Е. coli вызывающий диарею, часто изолируют при энтеритах у телят, и, исключительно, редко при септицемийном колибактериозе [G. Sivaswamy, C. L. Gyles, 1976]. В очаге колибактериоза при энтеритной форме выделяют энтеротоксигенные типы Е. coli из фекалий телят и здоровых коров-матерей в достаточно высокой пропорции [H. W. Smith, 1971; G. Sivaswamy, C. L. Gyles, 1976; M. Decun, 1986].
Энтеротоксины имеют плазмидную природу. Ent+ плазмида контролирует синтез энтеротоксина у патогенных эшерихий. Передача фактора Ent+ при конъюгации происходит независимо или одновременно с факторами Hly+, K88+, соl+ и R [H. W. Smith, 1976; М. А. Сидоров, Т. К. Курашвили, 1978; H. W. Smith, 1978; В. А. Ушкалов, 1994].
Повышение секреторных процессов в кишечнике новорожденных после введения термолабильного энтеротоксина Е. coli происходит вследствие активизации гуанилциклазы в слизистой кишечника, которая вызывает образование внутриклеточного циклического гуанозин 3,5-монофосфата [W. Gaastra, F. K. de Graaf, 1982; H. de Jonge, 1984; C. L. Gules and K. K. Baxi, 1987].
Термолабильный энтеротоксин Е. coli обладает антигенными свойствами и в отличие от термостабильного энтеротоксина имеет специфические рецепторпые зоны на слизистой кишечника новорожденных. Этот знтеротоксин подобно холерному вибриону (Vibrio сholerae) стимулирует активность аденилциклазы, которая вызывает накопление внутриклеточного циклического аденозин 3,5-монофосфата, что приводит к нарушению секреции воды и электролитов и развитию диареи в первые 10-18 ч его действия.
В качестве тестов для выявления термостабильного энтеротоксина могут быть использованы проба на инфантильных мышатах и проба с расширением перевязанного сегмента тонкого кишечника поросят и телят [Н. И. Романенкова, В. Е. Ефремов, В. Е. Клеганов, 1963; H. Fey, 1972; S. C. Whipp, H. W. Moon and N. C. Lyon, 1975; P. M. Newsome, M. N. Burgess, R. J. Bywater, C. M. Cowley, N. A. Mullan, 1978; C. M. Wise, A. P. Knight, M. J. Lucas, 1983; J. P. Dube, 1990].
Для обнаружения термолабильного энтеротоксина использует пробу с перевязанным сегментом тонкого кишечника кролика или морской свинки [H. W. Moon and S. C. Whipp, 1971; M. M. Levine, D. R. Nalin, R. B. Hornick, 1978; J. S. Seriwatana, P. Echeverria, D. N. Taylor, 1988].
1.1.6. Клинические симптомы колибактериоза. Большинство авторов характеризует колибактериоз, как инфекционное заболевание протекающее в трех формах: септицемической, энтеритной и энтеротоксемической [Ф. Гутира, И. Марек, Р. Маннингер, И. Мочи, 1961; Г. В. Гнатенко, 1968; Л. Н. Головнев, 1970; Я. Е. Коляков, С. С. Гительсон, Л. С. Каврук, 1970; W. J. Sojka, 1970; H. Fey, 1972; T. Gossling, K. A. McKay and D. A. Barnum, 1974; Л. К. Волынец, 1975; К. Эльце, Х. Мейер, Г. Штейнбах, 1977; J. N. Roy, 1980; E. Salajka, 1980; M. E. Amstutz, 1985; M. Decun, 1986; В. С. Шипилов, В. П. Шишков, В. Г. Зароза, В. П. Карев, Г. Д. Смоленская, 1987; A. K. Gupta and K. K. Baxi, 1987; Ю. Н. Федоров, 1988; D. H. Zeman, J. U. Thomson, D. H. Francis, 1989; А. А. Гутковский, Г. Л. Дворкин, 1989; В. Г. Зароза, 1989; А. С. Ковалев, 1989; B. H. Janke, D. H. Francis, J. E. Collins, 1990; Г. П. Задорожняя, В. Д. Уманец, 1990; Ю. С. Кабанков, С. Ф. Армашу, 1991; В. Г. Зароза, 1991; М. А. Сидоров, 1996].
Колисептицемийная (септическая) форма протекает очень остро и, обычно телята погибают через 3-6 дней после рождения. Новорожденные телята внезапно отказываются от молозива (молока), находятся в состоянии прострации, не способны стоять, глаза запавшие, дыхание и сердцебиение учащенные, возможна диарея, но чаще отсутствует, животные подвергаются быстрой дегидратации, вызывающая гибель животного после коматозного состояния через один два дня после начала заболевания. У отдельных животных развиваются артриты и менингоэнцефалит. Из других признаков отмечают сонливость, парез, опистотонус, атаксию, судороги [T. K. Korhonen, M. V. Valtonen, J. Parkkinen, 1985].
Энтеритная форма характеризуется диареей и основным симптомом дегидратацией варьирующей от слабой до тяжелой. Фецес жидкий, желтого или серо-белого цвета. Телята не погибают быстро, они могут болеть в течение недели без прироста массы или могут даже выздоравливать. Эта форма наиболее часто встречается в Англии [W. J. Sojka, 1971], Ирландии [M. Morrin, S. Lariviere, R. Lallier, 1976], Канаде и США [J. Hadad, C. L. Gyles, 1982], тогда как в Швейцарии преобладает септицемическая форма колибациллеза [H. J. Greene, 1984].
Энтеротоксемическая форма колибациллеза характеризуется коллапсом и чрезмерной прострацией. Течение очень быстрое, не наблюдается диарея, смерть наступает обычно в пределах 6-16ч от начала заболевания, животное теряет мышечный тонус и становится параличным. При данной форме бактериемия не развивается, но массивная пролиферация Е. coli наблюдается в нижней и задней части тонкого кишечника. Штаммы Е. coli мукоидного типа, принадлежащие к О группам 9 и 10 и обладающие капсулярным антигеном А внедряются в мезентериальные лимфатические узлы [С. С. Gay, 1971; С. С. Gay, S. M. Parish, T. C. McGuire 1982].
1.1.7. Патогенез. Основным путем инфицирования телят большинство исследователей считают алиментарный. Не исключается также заражение через носоглотку, внутриутробно и другими путями [Я. Е. Коляков, С. С. Гительсон, Л. С. Каврук, 1970; H. Fey, 1972; B. Tennant, D. Harold and Reina- M. Guerra, 1972; B. Tennant, D. Harold, Reina- M. Guerra, 1975; Дж. Х. Б. Рой, 1982; R. D. Linnabary, D. F. Dean, 1983; У. Риихикоски, 1986; I. Castrucii, F. Frigeri, M. Ferrari, 1987; А. А. Гутковский, Г. Л. Дворкин, 1989; В. Г. 3ароза, 1991].
В результате быстрого размножения бактерий и значительного увеличения их количества в кишечнике происходит быстрый распад микробов, при котором высвобождаются токсические продукты эндотоксины, вызывающие воспаление желудка и кишечника, вследствие чего резко усиливается перистальтика, что также следует рассматривать как рефлекторную защиту организма, стремящегося быстрее вывести из кишечника раздражающий агент [M. Morrin, S. Lariviere, R. Lallier, 1976].
При недостаточной активности защитных механизмов кишечной стенки и, прежде всего фагоцитоза, эшерихии проникают в лимфатическую и кровеносную системы и вызывают септический процесс, который приводит к летальному исходу.
Начавшийся понос вызывает резкое обезвоживание тканей организма. Размножение в крови бактерий и наводнение организма токсическими продуктами их жизнедеятельности и тканевого распада угнетают деятельность центральной нервной системы, что дает к концу заболевания картину теплого коматозного состояния.
Новорожденный теленок является полностью неспособным противостоять инфекционным заболеваниям, и кишечник его стерилен. Поэтому теленку необходимо в течение первых четырех часов жизни выпоить не менее двух литров молозива. С молозивом теленок получает антитела, которые препятствуют размножению бактерий и вирусов, у него формируется пассивный иммунитет. Приобретенная таким образом способность к сопротивлению продолжается в течение 3-4 недель, после чего у теленка в 4-5 недельном возрасте развивается активный иммунитет [Th. Smith, and M.L. Orcutt, 1925; E. F. Logan, and W. J. Penhale, 1971; W. J. Penhale, E. F. Logan, A. Stenhouse, 1971; I. E. Selman, G. H. de la Fuente , E. W. Fisher and A. D. MсEwan, 1971; E. F. Logan, 1974; J. N. Roy, 1980; G. H. Stott A. Pellah, 1982; Т.Е. Besser, 1991; L. J. Perino, 1995].
Синдром дефицита антител (СДА) появляется у подсосных телят, если они получают недостаточное количество молозива или в молозиве отсутствуют гаммаглобулины. Содержание гаммаглобулинов в сыворотке крови в количестве более 500 мг обеспечивает невосприимчивость к колибактериозу. С другой стороны установлено, что 82,4% телят, болевших колибактериозом имели выраженный СДА [R. Kruedener, 1970].
Колибактериоз вызывается небольшим количеством энтеропатогенных штаммов, продуцирующих энтеротоксины, к которым новорожденные животные обладают повышенной чувствительностью [E. M. Kohler, 1971; A. Kircher, 1981].
Энтеротоксины Е. coli стимулируют аденилциклазу в эпителиальных клетках кишечника. Этот энзим действует каталитически при образовании циклического 3’,5’-аденозинмонофосфата (сАМР) из аденозинтрифосфата (АТР). Увеличение количества 3,5 аденозинмонофосфата с (АМР) вызывает изменение транспортировки ионов, ингибируя активную абсорбцию натрия и стимулируя выделение ионов хлора и бикарбоната, вследствие чего происходит накопление жидкости в просвете кишечника и в результате наблюдается сильный понос [E. Salajka, 1980].
Многие исследователи полагает, что дефицит витамина А является главным условием для возникновения колибациллеза у телят [Н. И. Старикова, 1994; Н. И. Старикова, 1994].
Телята, родившиеся от коров с низким содержанием витамина А в молозиве были более подвержены заболеваниям, таким как белый понос, пупочная инфекция, заболевание суставов, по сравнение с телятами, родившимися от коров с относительно высоким содержанием витамина А [F. Blakemore, A. Davies, E. Eylenburg, Т. Moore, K. C. Sellers and A. N. Worden, 1948]. Взаимосвязь между дефицитом витамина А и колибациллезом также установили и другие ученые: [R. A. Willoughby, D. G. Buttler and J. K. Thorton, 1970; H. Fey, 1972; R. G. Hansen, P. H. Phillips, G. W. Rupel, 1976; J. N. Roy, 1980; H. Fey und S. Lindt, 1982; G. J. Pearson, E. F. Logan, 1986]
По некоторым данным [W. J. Sojka, 1970; H. Fey und G. Hunyady, 1972; В. М. Чекишев, В. М. Васильев, А. И. Кабанцев, 1983; R.K. Braun, B.C. Tennant, 1983; A. S. Sheikh P. S. Bradford, S. C. James, B. Patricia, B. F. Thomas, H. Richard, D. George, D. R. Lin, W. Bert, 1995], решающая роль иммуноглобулинового дефицита в патогенезе колисеятицемии у неонатальных телят, принадлежит грам-негативным бактериям.
1.1.8. Специфическая профилактика и терапия. Неуклонное выполнение комплекса зоотехнических, зоогигиенических и организационных мероприятий является незыблемой основой в деле получения, сохранения и выращивания молодняка. Исходя из этого для предупреждения колибактериоза, прежде всего, необходимо создать хорошие условия кормления и содержания стельных коров и новорожденных телят. Изоляция новорожденных телят в специальном изолированном приёмном отделении является важнейшим звеном в цепи эффективных профилактических и противоэпизоотических мероприятий против колибактериоза телят. Необходимость и эффективность таких мероприятий признаны давно, и проведение их рекомендуется многими учеными [H. Fey, 1972; Л. К. Волынец, 1978; А. В. Драгомир, 1982; В. Т. Самохин, М. А. Сидоров, Г. К. Волков, 1983; П. А. Рыдак, 1984;С. И. Джупина, И. И. Фельдман, В. М. Чекишев, 1986; P. Г. Иксамов, М. П. Неустроев, 1987; Г. Ф. Коромыслов, Н. Н. Михайлов, 1987; А. А. Гутковский, Г. Л. Дворкин, 1989; Т. Otoi, Т. Toujou, H. Toujou, M. Hasimoto, 1990; В. Г. 3ароза, 1991; С. И. Плященко, 1991; Г. К. Волков, В. Н. Гущин, В. Д. Баранников, 1996; E. С. Воронин, Д. А. Девришов, Р. В. Петров, В. П. Шишков, 1996; А. Н. Куриленко, 1996].
Раннему выпаиванию молозива придают большое значение [Я. Е. Коляков, С. С. Гительсон, Л. С. Каврук, 1970; A. D. MсEwan, E. W. Fisher, I. E. Selman, 1970; T. C. Guire, N. E. Pfeiffer, J. M. Weikel and R. C. Bartsch, 1976; Я. Е. Коляков, 1978; М. И. Немченко, Т. Д. Гришина, 1983; М. И. Немченко, 1984; В. Г. 3ароза, 1985; R. Morar, 1985; Г. В. Полушин, 1987; E. С. Воронин, Д. А. Девришов, Л. Я. Ставцева, 1989; М. И. Немченко, 1989; E. С. Воронин, Л. Я. Ставцева, Т. Н. Грязнева, 1990; P.E. Howe, 1991; К. Ф. Думбур, В. В. Левенец, О. Н. Ткаченко, А. К. Думбур, 1996]. При хранении молозива химический состав его быстро приближается к составу обычного молока, причем кислотность молозива у одних коров понижается быстро, в то же время как у других постепенно. Снижение кислотности молозива неблагоприятно влияет на организм теленка, в таких случаях отмечается большой процент заболевания и отхода.
Специфическая профилактика основана на пассивной иммунизации новорожденных сывороткой. О положительном применении антиколибактериозной сыворотки сообщали многие исследователями [P. Ehrlich, 1892; C. O. Jensen, 1905; Ф. Гутира, И. Марек, Р. Маннингер, И. Мочи, 1961; С. И. Губкин, А. И. Коган, 1971; A. Dam, 1971; E. F. Logan, and W. J. Penhale, 1971; Я. Е. Коляков, 1978; Y. Danieli, I. Kornitzer, R. Tamarin, 1979; А. Т. Флюстиков, 1982; В. А. Аликаев, В. В. Матюшин, 1983; Е. В. Андреев, М. Д. Бакуменко, А. К. Панасенко, В. И. Тертышник, Л. Т. Лещенко, Р. И. Бессараб, 1983; Г. В. Гнатенко, Л. Т. Тупица, 1984; В. М. Чекишев, Т. В. Бондарь, О. А. Колганова, 1985; Г. В. Полушин, 1987; C. M. Scanlan, 1988; Е. С. Сухарев, 1994; М. Х. Шайхаманов, В. П. Грамолин, Б. М. Авакаяц, 1994; Г. К. Волков, В. Н. Гущин, В. Д. Баранников, 1996; B. C. Солодков, Е. Э. Школьников, Г. Ф. Коломнина, Е. М. Лукьянова, В. Н. Алейников, 1996; С. В. Вальциферова, 1997] .
Противоколибактериозную сыворотку особенно важно вводить телятам, которые были лишены молозива и молодняку, матери которых были переведены из другого хозяйства и поэтому содержат в молозиве антитела, несоответствующие микрофлоре нового помещения [B. C. Шипилов, В. К. Копытин, 1984; C. Staak, 1992].
Исследования [M. M. EI-Nageh, 1967; H. Fey, 1971a; H. Fey und G. Hunyady, 1972; B. Tennant, B.H. Baldwin, 1979; H. Fey und A. Margadant, 1981; М. И. Немченко, Т. Д. Гришина, 1983; Ю. Н. Федоров, И. Д. Фесенко, М. Ю. Горбунова, С. О. Кодыров, И. Ю. Пичкова, 1983; В. М. Чекишев, В. М. Васильев, А. И. Кабанцев, 1983; М. И. Немченко, 1984; В. Т. Сидоров, 1984; B. C. Шипилов, В. П. Шишков, В. Г. Зароза, В. П. Карев, Г. Д. Смоленская, 1987; A. Badiu, T. Nedelcu, 1989; Ю. Н. Федоров, 1996; Ю. Н. Федоров, О. А. Верховский, 1996], показали большое значение гипогаммаглобулинемии, как одного из важных предрасполагающих факторов в возникновении колибактериоза у новорожденных телят. Авторы установили, что у всех телят павших от колисептицемии, наблюдалась гипо- или агамаглобулинемия [A. K. Lascelles, G. H. Me Dowell, 1974; T.C. McGuire, D.S. Adams, 1982].
Наличие К агглютининов в сыворотке молозива, защищающих телят от колисептицемии и других форм колибациллеза обнаружил С. С. Gay, 1971. Им установлено, что 45 телят из 59 кормившихся молозивом погибли вследствие отсутствия К агглютининов в скармливаемом молозиве.
Аналогичные исследования о дефиците К агглютининов в молозиве коров и, следовательно, о развитии колисептицемии у новорожденных телят провели многие ученые [C. Briggs, 1951; H. W. Smith and M. A. Linggood, 1972; F. Orskov, G. Orskov, H. W. Smith and W. J Sojka, 1975; B. Kaijser, S. Ahlstedt, 1977; В. Б. Федотов, 1982; J. Hadad, C.L. Gyles, 1982].
Успехи, достигнутые в изучения антигенной структуры возбудителя колибактериоза телят, позволили ученым предложить активные средства специфической профилактики этого заболевания иммунные сыворотки и лактоглобулины [Ф.Н.Щепетов, 1951; R. Aschaffenburg, S. Bartlett, S. K. Kon, J.M. Roy and D. M. Walker, 1951a; P. Lepine, 1963; К. Геров, П.Чушков, Р. Георгиева, 1965; В. К. Чернуха, 1968; С.И.Губкин, А. И. Коган, 1971; Н. С. Жосан, 1974; Н. С. Жосан, 1976; Е.В.Гублер, А. А. Генкин, 1978; L. S. Young, 1984; В. Б. Билоштан, 1985; В. П. Павлов, Т. Н. Грязнева, Е. В. Чичикова, 1988; Н. С. Жосан, 1992; В. В. Бурсуков, 1993; W. Zaremba, 1993; С. В. Вальциферова, 1997].
О положительных результатах фаготерапии в ранних случаях заболевания телят колибактериозом сообщали многие исследователи [К. Н. Шерстобаев, 1944; С. Н. Муромцев, 1947; Я. Е. Коляков, С. С. Гительсон, Л. С. Каврук, 1970; H. Fey, 1972; Л. Б. Борисов, 1976; Я. Е. Коляков, 1978; А. С. Овод, А. Г. Морозов, 1986; M. Decun, 1986; А. А. Гутковский, Г. Л. Дворкин, 1989].
Для лечения диареи у новорожденных телят У. Риихикоски (1986) рекомендует следующую программу:
голодная диета;
лечение с применением минеральных солей;
лечение антибиотиками;
лечение путем повышения кислотности внутри кишечника;
профилактика всасывания в кровь бактерий и их токсинов;
создание животным хороших условий содержания и обеспечение их качественными кормами.
В качестве неспецифических профилактических и лечебных средств при колибактериозе рекомендуются и используются различные антибиотики [И. Е. Мозгов,1968; W. J. Sojka, 1971; H. Fey, 1972; B.J. Buntain and I.E. Selman, 1980; E. С. Фортинская, А. М. Наумова, Е. А. Маркова, 1983; H. J. Greene, 1983; М. А. Сидоров, В. Н. Гущин, 1984; H. Balbierz, L. Kuchar, 1984; С. И. Джупина, И. И. Фельдман, В. М. Чекишев, 1986; G. Rademacher, G. Dirksen, 1986; А. А. Гутковский, Г. Л. Дворкин, 1989; Г. П. 3адорожняя, В. Д. Уманец, 1990; В. А. Антипов, 1991; Ю. Юнисова, Т. П. Жарова, 1996; П. А. Паршин, С. В. Шабунин, 1997].
В связи с очень широким применением антибиотиков, приобрела особо важное значение проблема антибиотикоустойчивости микробов. Увеличение антибиотикоустойчивых штаммов микробов может изменить микробиологический, клинический, возможно эпизоотологический фон ряда инфекционных болезней и, в первую очередь, массовых заболеваний молодняка. Антибиотикоустойчивые штаммы кишечной палочки, выделяясь в большом количестве с фекалиями во внешнюю среду инфицируют помещения, предметы ухода за телятами, посуду и др., что приводит к попаданию и заселению кишечника новорожденных клинически здоровых телят. Поэтому в настоящее время антибиотики применяются предварительным определением чувствительности изолированных микробов к антибиотикам [Г. В. Гнатенко, 1968; Я. Е. Коляков, 1978; R. J. Bywater, 1983; Л. А. Таранова, 1984; E. С. Фортинская, А. М. Наумова, Е. А. Маркова, 1983; В. А. Антипов, 1991].
Исследованиями ряда авторов [J. W. Boyd, J.R. Baker, A. Leyland, 1974; К. Эльце, X. Мейер, Г. Штейнбах, 1977; O. M. Radostis, S. D. Acres, 1983; Ю. А. Ширванян, А. А. Акопов, 1985; M. Decun, 1986; А. А. Гутковский, Г. Л. Дворкин, 1989] была установлена высокая чувствительность преобладающего числа штаммов Е. coli к cульфаниламидным препаратам, тогда как, их устойчивость к некоторым антибиотикам значительно повышалась.
Для профилактики колибактериоза новорожденных телят М. А. Сидоров с соавт. (1978, 1980, 1996) рекомендует применять колипротектан сразу после рождения в дозе 10-15 мл per os и затем в течение двух дней в той же дозе (всего 6 раз). Колипротектан способствует резкому снижению заболеваемости (до 10-20%) и повышению сохранности телят (до 95-98 %). Применению материнской и гетерогенной крови при острых желудочно-кишечных заболеваниях новорожденных животных посвящено немало работ отечественных и зарубежных авторов [H. Fey, 1972; Е. В. Андреев, М. Д. Бакуменко, А. К. Панасенко, В. И. Тертышник, Л. Т. Лещенко, Р. И. Бессараб, 1983; Дж. Х. Б. Рой, 1982; У. Риихикоски, 1986; А. А. Гутковский, Г. Л. Дворкин, 1989]. Опыты [В. П. Урбан, 1966; В. К. Чернуха, 1968; В. П. Урбан, И. Л. Найманов, 1985; В. П. Павлов, Т. Н. Грязнева, Е. В. Чичикова, 1988] показали, что использование сывороточных поли- и гаммаглобулинов бывает более результативным, чем применение цельной крови.
Из симптоматических средств E. W. Fisher and de la G. H. Fuente, 1972; H. Rascova, 1974; Е. Рашкова, Т. Сехер, К. Рашка, В. Матейовска, Л. Палак, 1976; J. Blanu, J. H. Parvu, O. Lvanciu, 1983; R. J. Bywater, 1983; В. В. Митюшин, 1984; H. J. Greene, 1984; I. M. Naylor, 1986; Г. Ангелов, Б. Абделмалек, 1987; I. M. Naylor, 1987; А. И. Теш, 1990; Н. Т. Винников, 1993; А. Г. Шитый, Н. С. Дудникова, Л. М. Тихомирова, 1993; М. Х. Шайхаманов, В. П. Грамолин, Б. М. Авакаяц, 1994, рекомендуют при колибактериозе телят, сопровождающимся профузным поносом и сильным обезвоживанием организма, применение водно-электролитных растворов. В связи с тем, что объем плазмы крови уменьшается и она становится гипоосмотической по отношению к кишечной жидкости, так как не только вода, но н ионы Na+, K+, Cl- и HCO>3>- выходят из плазмы в просвет кишечника, а слизистая кишечника превращается в секреторный орган, всасывание воды и электролитов прекращается. Авторы рекомендуют в начале болезни, при легком течении, изотонические растворы а при тяжелом течении — парентерально — гипертонические, с обязательным включением ионов натрия, хлора и бикарбоната.
Некоторые авторы рекомендуют витаминотерапию, например гексавит (А>1>, В>1>, В>2>, В>6>, С, Е, Р). Рутин увеличивает продолжительность циркуляции солевых кристалловидных растворов в кровяном русле и тем самым уменьшает проницаемость капилляров. Витамин С устраняет гипоксию и метаболический ацидоз, содействует нормализации окислительно-восстановительных процессов [F. Blakemore, A. Davies, E. Eylenburg, Т. Moore, K. C. Sellers and A. N. Worden, 1948; R. G. Hansen, P. H. Phillips, G. W. Rupel, 1976; В. А. Аликаев, В. В. Матюшин, 1983; В. И. Федюк, 1983; У. Риихикоски, 1986; А. А. Гутковский, Г. Л. Дворкин, 1989; Я. Я. Самуйленко, 1996].
Антисекреторные факторы (хлорпромазин и другие производные фенотиазина, никотиновая кислота, опий, аспирин, соматостатин, гликокортикоиды, амфотерицин В, производные барбариса и др.) приводят к нормальной абсорбции электролитов и воды в кишечнике, ингибируя чрезмерную секрецию. Хлорпромазин обладает сильным антисекреторным действием, ингибируя диарею, вызванную энтеротоксином TL — Е. coli. Этот препарат ограничивает чрезмерную активность циклического аденозинмонофосфата (сАМР). Он действует анти-секреторно при лечении диареи, вызванной энтеротоксином TS — Е. coli.
Кроме анти-секреторной деятельности фенотиазиновые препараты обладают довольно сильным бактерицидным действием, по отношению к некоторым штаммам Е. coli. Кроме того данные препараты воздействуют на плазмиды, ответственные за синтез адгезивных фимбрий, энтеротоксинов, а также препятствуют комплексной резистентности эшерихий к антибиотикам и тормозят синтез компонентов адгезивных антигенов, ограничивающие колонизирующее действие [A. I. Furowicz, W. Zyska, 1988].
1.2. Специфическая иммунологическая реактивность телят в постнатальный период
Молозиво является для новорожденного исключительно полноценной и легко усвояемой пищей. Обеспечивая потребности новорожденного в необходимых питательных веществах, оно, кроме того, является носителем иммунных тел и антибактериальных веществ, передающихся от матери потомству, служащих для защиты молодого организма от инфекции на первом этапе жизни.
Природа защитных свойств молозива была предметом длительного изучения. Впервые исследования по выяснению роли молозива в создании иммунитета телят начались после опубликования работ P. Ehrlich [1892], который установил, что у детенышей, рожденных от мышей, иммунизированных рицином и абрином невосприимчивость к этим ядам не передается плацентарным путем, а появляется только после сосания молозива иммунизированных мышей.
Оценивая эксперименты Эрлиха И. И., М. Мечников [1951] писал: «В своих этюдах об унаследованном иммунитете Эрлих указал еще на другой важный фактор, именно на прямую передачу материнских антител в молоко при сосании. Он прямо показал своими опытами над мышами, происходившими не от иммунных матерей, а вскормленных иммунными самками, что благодаря этому, молодые мыши были иммунизированы к рицину, абрину и столбнячному токсину. Это интересное поле исследований дало много важных результатов, а в будущем, наверное, будет еще многое достигнуто в этом направлении».
У различных видов животных, у которых эпителиохориальная плацента (лошади, крупный рогатый скот, свиньи, козы) антитела не проникают из материнского организма в плод плацентарным путем. У этих видов животных передача антител от матери новорожденному возможна колостральным путем (через молозиво). С молозивом новорожденные жвачные получает необходимый запас иммунных тел. [П. П. Емельяненко, 1987; Ю. Н. Федоров, 1988; И. М. Карпуть, 1993]
Иммунологическое исследование молозива показало, что концентрация антител в молозиве превосходит таковую в сыворотке крови. Больше того, молозиво содержит антитела, отсутствующие в сыворотке [В. А. Фортушний, 1984].
Результаты исследований Th. Smith and R.B. Little, 1922b; R. S. Comline H. E. Roberts, D. A. Fitchen, 1951; R. S. Comline, H. E. Roberts, D.A. Fitchen. 1951; H. Fey, 1968; A. Kaeckenbeeck, G. Colinet, F. Schoenaers, 1971; P. D Porter, 1972; W. J. Penhale, E. F. Logan, I. E. Selman, E. W Fisher, 1973; У. Дж. Герберт, 1974; G. H. Stott and B. E. Menefel, 1977; J. A. Patt, 1977; И. И. Головистиков, 1979; G. H. Stott, D. B. Marx, B. E. Menefel and G. T. Nightengale, 1979; G. H. Stott, D. B. Marx, B. E. Menefel and G. T. Nightengale, 1979; G. H. Stott, D. B. Marx, B. E. Menefel and G. T. Nightengale, 1979; Т. Brignole, J. Stott, 1980; L. J. Buch, Т. Е. Staley, 1980; И. М. Карпуть, В. М. Холод, 1982; G. H. Stott A. Pellah, 1982; Ю. Н. Федоров, И. Д. Фесенко, М. Ю. Горбунова, С. О. Кодыров, И. Ю. Пичкова, 1983; Э. Купер, 1985; В. М. Чекишев, Т. В. Бондарь, О. А. Колганова, 1985; B. C. Шипилов, В. П. Шишков, В. Г. Зароза, В. П. Карев, Г. Д. Смоленская, 1987; Н. С. Жосан, 1988; В. Г. 3ароза, 1989; С. И. Плященко, В. Т. Сидоров, А. Ф. Трофимов, 1990; Т. Е. Besser, O. Szenci, C. C. Gay, 1990; Ю. Н. Федоров, О. А. Верховский, 1996; F. В. Carry, 1996, также свидетельствуют о том, что абсорбция иммуноглобулинов молозива происходит обычно в течение первых 24—36ч жизни животного, постепенно замедляясь к 10-12 ч возраста вплоть до полного прекращения. Всасывание иммуноглобулинов происходит в кишечнике, и через лимфатические пути они попадают в кровь. Иммуноглобулины появляются в лимфе уже через 1-2 ч после поступления их в двенадцатиперстную кашку животного.
Большое значение молозива в создании иммунитета у новорожденных телят впервые отметили P. H. Romer, H. Much [1906]. В дальнейшем их данные были подтверждены исследованиями многочисленных авторов [R. Aschaffenburg, S. Bartlett, S. K. Kon, P. Terry, S. Y. Thompson and D. M. Walker, 1949a; R. Aschaffenburg, S. Bartlett, S. K. Kon, D. M. Walker, C. Briggs, E. Cotchin and R. Lovell, 1949b; R. Aschaffenburg, S. Bartlett, S. K. Kon, J. M. Roy and D. M. Walker, 1951a; R. Aschaffenburg, S. Bartlett, S. K. Kon, S. Y. Roy, D. M. Walker C. Briggs and R. Lovell, 1951b; C. Briggs, 1951; E. Selman, A. D. McEwan and E. W. 1970; P. Blackmer, 1973; V.C.R. Irvin, 1974; С. В. Вальциферова, 1997; L. Hirszfeld et J Lille-Azyszkowierz, 1979; J. N. Roy, 1980; И. М. Карпуть, В. М. Холод, 1982; С. С. Gay, 1984; J. J. Geene, 1984; R. Morar, 1985; Г. Н. Печникова, Т. В. Окунева, О. Н. Грызлова, 1988; И. М. Карпуть, А. Г. Ульянов, В. И. Бабин, 1990; P.E. Howe, 1991; C. Staak, 1992; W. Zaremba, 1993; Т. Е. Besser, C. C. Gay, 1994; P. P. Игнатьев, Г. Ч. Бондаренко, 1994; F. В. Carry, 1996; Д. Н. Масюк, 1997; L. J. Perino, 1997].
В сыворотке крови новорожденных телят не обнаруживали агглютинины [Th. Smith and R. B. Little, 1922; Th. Smith and R.B. Little, 1922b; S. C. Whipp and S. T. Donta, 1976; A. N. Thepfilopoulos, F.J. Dixon, 1979; J. M. Tyler, J. S. Cullor, M. C. Thurmond, 1989; K. Walser und H. Brumer, 1987] к кишечной палочке, хотя в сыворотке крови матерей и в молозиве они имелись в значительном количестве. После поения теленка молозивом в сыворотке его крови указанные агглютинины появлялись. Более того, молозиво, полученное в первые часы после отела, в дозе 0,2 мл предохраняло морских свинок от заражения 1-1,5 минимальными смертельными дозами культуры кишечной палочки. Авторы пришли к заключению, что кормление телят молозивом имеет огромное значение в создании невосприимчивости к колиинфекции.
В молозиве коров обнаружен иммунный глобулин, имеющий огромное значение, как носитель антител, в создании устойчивости новорожденных телят к инфекционным заболеваниям [H. W. Smith, 1971; H. W. Smith, 1976; H.W. Smith, 1978] .
Исследования механизмов транспорта и катаболизма иммуноглобулинов in vitro показало, что в начале всасывания образуется комплекс между иммуноглобулинами и специфическими рецепторами энтероцитов. Во взаимодействии с рецепторами клеток кишечника участвует Fc-фрагмент иммуноглобулинов. Вероятно, связывание иммуноглобулинов с клетками предохраняет их от катаболизма и способствует транспорту в кровь [D. Eddie, M. Sohulkind, I. Robbins, 1971; И. И. Головистиков, 1979]. Процесс переноса наиболее активно идет при рН 6,0-6,5, что соответствует кислотности жидкости в тонком отделе кишечника новорожденных.
Иммуноглобулины крупного рогатого скота были идентифицированы как аналоги по физико-химической и антигенной характеристике к человеческим IgG, IgA и IgM. Два класса IgG: IgG>1> и IgG>2> иммунологически и биологически различались. Секреторный IgA был идентифицирован в эпителии слизистой оболочки кишечника в секреторных компонентах. IgG>1> является доминирующим в молочной секреции, IgM превышает IgA в секреции пищеварительного тракта преруминантных телят [P. D. Porter, 1973; J. A. Roth, J. W. Sexton, 1975; R. Sakulramrung, G. L. Domingae, 1985; G. Riedel-Caspari, 1993;] .
IgM и IgA играют важную роль в местной (локальной) реакции вымени при заражения бактериальными антигенами, а также, вероятно, эти два иммуноглобулина взаимодействуют в защитном механизме новорожденных телят [R. A. Wilson and I. W. Jutila, 1976; P. Pivont, R. Gregoire and H. Antoine, 1984; L. S. Young, 1985; M. L. Wickstrom, 1987].
У новорожденных интестинальная адсорбция колостральных антител не селективная, таким образом иммуноглобулиновый профиль сыворотки постколостральных телят имеет сходство с молозивом и это обеспечивает необыкновенно высокий уровень секреторного IgA в циркуляции крови. Секреторные IgA и IgM существенно содействуют пассивному иммунитету, хотя их продолжительность в сыворотке крови телят составляет соответственно только 2 и 4 дня [G. G. B. Klaus, A. Bennett and E.W. Jones, 1979; R.K. Braun, B.C. Tennant, 1983; J. Brenner, 1991].
Концентрация иммуноглобулинов в молоке коров снижается быстро в течение первых 3 дней лактации и, маловероятно, что содействует локальной защите алиментарного тракта телят после первой недели. Тем не менее, локальный синтез иммуноглобулинов является доказуемым в иммуноцитах, находящихся на конце эпителиальных клеток пищеварительного тракта и секреция иммуноглобулинов начинается на 2-ой неделе жизни [H. Fey, 1967; E. F. Logan, and W. J. Penhale, 1971; J. E. Butler, C. A. Kiddy, C. Maxwele, M. B. Hylton, A. Asofsky, 1971; J.W. Boyd, 1972; P. D. Porter, D. E. Noakes, and W. D. Allen, 1972; E. F. Logan, 1974; В. М. Чекишев, 1977; Ю. Н. Федоров, И. Д. Фесенко, М. Ю. Горбунова, С. О. Кодыров, И. Ю. Пичкова, 1983; В. М. Чекишев, Т. В. Бондарь, О. А. Колганова, 1985; Т. Е. Besser, 1988].
Эффективность абсорбции лактоглобулинов связана с количеством проглоченного молозива [I. E. Selman, A. D. McEwan, E. W. Fisher, 1971; Т. Brignole, J. Stott, 1980; С. С. Gay, Т. McGuire, S. M. Parish 1983;] и влиянием присутствия коров матерей в течение первых 24 ч жизни [E. Selman, A. D. McEwan and E. W. 1970; D. F. Wolfe, 1985; Ю. Н. Федоров, О. А. Верховский, 1996].
В течение двух часов после приема молозива иммуноглобулины появляются в сыворотке и, через четыре-восемь часов — в синовии. Иммуноглобулиновый профиль синовии имеет сходство с сывороткой крови телят и коров-матерей [H. Fey, 1972]. Это объясняет, почему телята, получившие молозиво более резистентные к септицемии и полиартритам, чем телята, которые недостаточно получают молозиво.
Молозиво превосходный источник энергии, протеина, витаминов А, Д и комплекса витаминов В, а также кальция, фосфора, магния и хлоридов. При дефиците указанных компонентов молозива и, особенно, витамина А резистентность к Е. coli инфекции резко снижается. [Th. Smith and R. B. Little, 1922; H. Fey und S. Lindt, 1982; С. С. Gay, 1984; J. J. Geene, 1984; R. Morar, 1985; P.E. Howe, 1991]
Молозиво коров содержит высокий уровень иммуноглобулинов, среди которых 75 % IgG, IgA и IgM вместе составляют около 20 % от иммуноглобулинов молозива [J. A. Roth, J. W. Sexton, 1975].
По данным исследователей [E. W. Fisher, 1971; J. W. Boyd, 1972; E. W. Fisher, A. A. Martinez, Z. Trainin, 1975], риск заболеваемости и летальности можно предсказать у телят 48 ч возраста при определении уровня сывороточных иммуноглобулинов. Новорожденные телята с уровнем IgG и IgM меньше чем 0,8 и 0,2 мг/мл соответственно, погибали от неонатальной септицемии, тогда как у телят с уровнем IgG и IgM 5,0 и 0,6 мг/мл соответственно, развивалась энтеральная инфекция. Телята с постколостральным сывороточным уровнем 7,5 и 0,8 мг/мл IgG и IgM соответственно, были нормальными (клинически здоровы). Следовательно, телята с умеренно низким уровнем иммуноглобулинов имели достаточный иммунитет, предупреждающий септицемию, но не профилактировали энтерит.
Многочисленные исследования показывают, что имеется корреляция между пассивно приобретенными сывороточными иммуноглобулинами и выживанием неонатальных телят [H. Fey, 1967; H. Fey, 1968; E. F. Logan, A. Stenhouse, D. J. Ormorod and W. J. Penhale, 1974; T. A. Ferris, J. W. Thomas,1975; R. K. Braun, B.C. Tennant, 1983; P. Dobbelaar, I. P. T. M. Noordhuizen and K. Van A. S. Keulen, 1987; A. Lacheretz, J. Chontol, P. Desmettre, 1987].
Определение уровня сывороточного иммуноглобулина у новорожденных является клинически важным, так как их количество оказывает влияние на восприимчивость телят к заболеванию. Иммуноглобулиновые фракции составляют примерно одну третью значения общего сывороточного протеина. Иммуноглобулин дефицитные телята имеют низкое значение общего протеина, на это указывают показания рефрактометра. Рефрактометр не дает прямое измерение сывороточного иммуноглобулина подобно цинк-сульфатному, натрий-сульфитному и глутаральдегидному тестам, но предусматривает быстрый подсчет уровня сывороточного иммуноглобулина и может легко применяться в практической ветеринарии [D. G. McBeath, W. J. Penhale, E. F. Logan,1971; A. D. Me Ewan, E. W. Fisher, I. E. Selman and W. J. Penhale, 1970; N. E. Pfeiffer, T. C. McGuire, 1977; N. E. Pfeiffer, T. C. McGuire, R. E. Bendel and J. M Weikel, 1977; B. Tennant, B.H. Baldwin, 1979; H. Frerking, von E. Henkel and E. Schwartz, 1980; H. Balbierz, M. Nicolaijczuk, J. Zeilinski, 1983]
Объем сыворотки необходимый для рефрактометрического теста очень незначительный (примерно 0,2-0,3 мл) и если доступно центрифугирование, оценку концентрации иммуноглобулина, соответствующую требованиям, можно получить в пределах 30 мин из исследуемых проб. Рефрактометрический тест, как а другие (цинк-сульфатный, натрий-сульфитный, глутаральдегидный) зависят от гемоконцентрации и имеет незначительное значение при определении иммуноглобулинового статуса дегидратированных (обезвоженных) телят [D. G. McBeath, W. J. Penhale, E. F. Logan,1971; Ю. Н. Федоров, 1988].
Рефрактометр позволяет определить иммуноглобулиновый статус неонатальных телят с различной степенью риска. Норма этой оценки для телят голштинской породы крупного рогатого скота (молочного направления) следующая:
Высокий риск заболеваемости и летальности отмечают при уровне общего сывороточного протеина 4,9 г/100 мл;
Средний риск — при уровне общего сывороточного протеина 5,0-5,4 г/100 мл.
Низкий риск — при уровне общего сывороточного протеина 5,5-6,9 г/100 мл. [D. G. McBeath, W. J. Penhale, E. F. Logan,1971; R.K. Braun, B.C. Tennant, 1983]
Для изучения корреляции у крупного рогатого скота между пассивно приобретенными антителами и инфекционными заболеваниями были проведены многочисленные исследования, применяя метод количественного определения иммуноглобулинов [A. D. Me Ewan, E. W. Fisher, I. E. Selman, 1970; J. W. Boyd, 1972; W. J. Penhale, E. F. Logan, I. E. Selman, E. W Fisher, 1973; E. F. Logan, A. Stenhouse, D. J. Ormorod and W. J. Penhale, 1974; G. H. Stott and E. J. Reinhard, 1978; G. G. B. Klaus, A. Bennett and E. W. Jones, 1979; G. H. Stott, D. В. Marx, В. E. Menefel, and G. T. Nightengale, 1979; N. H. Teng, H. S. Kaplan, J. M. Herbert, 1985].
Наиболее широко применяются метод прямой радиальной иммунодиффузии [N. E. Pfeiffer, T. C. McGuire, 1977; В. М. Чекишев, 1977; N. E. Pfeiffer, T. C. McGuire, R. E. Bendel and J. M Weikel, 1977; А. А. Тихомиров, 1997] и цинк-сульфатный тест помутнения [A. D. Mс Ewan, E. W. Fisher, I. E. Selman and W. J. Penhale, 1970; Н. С. Жосан, 1989].
Наиболее точный и простой метод для тестирования уровня иммуноглобулинов, применяемый на ферме или в научной лаборатории является натрий-сульфитный преципитационный тест. Натрий сульфит в соответствующей концентрации (14, 16, 18 %) вызывает преципитацию иммуноглобулинов, подобно тому как и цинк-сульфатный тест. Значение помутнения, вызываемого взаимодействием натрия сульфита и иммуноглобулинов пропорционально степени защиты телят [N. E. Pfeiffer, T. C. McGuire, 1977; N. E. Pfeiffer, T. C. McGuire, R. E. Bendel and J. M Weikel, 1977; T.C. McGuire, D.S. Adams, 1982; Ю. Н. Федоров, 1988].
Для выявления гипогаммаглобулинемии у неонатальных телят применяют также быстрый скрининг тест. Тест основан на коагуляции иммуноглобулинов глутаральдегидом в 10 % концентрации. Иммуноглобулиновая концентрация у исследуемых телят, данным методом, была классифицирована как высокая 6 мг/мл, промежуточная 4,1-6 мг/мл и низкая 4 мг/мл [B. Tennant, B.H. Baldwin, 1979].
1.3. Неспецифическая иммунологическая реактивность новорожденных телят
Результаты многолетних исследований состояния неспецифической иммунологической реактивности организма сельскохозяйственных животных свидетельствует о том, что защитные силы их являются динамичным показателем и определяются как генетическими особенностями организма, так и воздействием различных факторов окружающей среды. Неблагоприятное воздействие окружающей среды приводит к ослаблению устойчивости организма и, как правило, к возникновению и распространению различных заболеваний, в том числе и инфекционных. Организм животных в процессе внутриутробного развития продуцирует клетки, осуществляющие фагоцитоз и молекулы с выраженным противомикробным действием — лизоцим, комплемент, пропердин, иммунные глобулины и другие факторы естественной резистентности. С возрастом животных они динамически изменяются [С. J. Howard, A. A. Glynn, 1971; В. В. Никольский, В. П. Литвин, 1974; П. А. Емельяненко, 1979; В. Т. Сидоров, 1984; B. C. Шипилов, В. К. Копытин, 1984; R.M. Miller, A.J. Guidry, M.J. Poape, 1988; G. Mulcahy, P.J. Gunn, 1988; С. И. Плященко, 1991].
Иммунологическая реактивность у новорожденных телят формируется постепенно и достигает полноценной выраженности только на определенном уровне их индивидуального физиологического развития [B. C. Бузлама, С. М. Сулейманов, В. Н. Долгополов, Н. Н. Коновалов, М. Н. Рецкий, И. С. Толкачев, Т. И. Агеева, 1978].
В качестве критериев степени реактивности животного организма используют [В. В. Никольский, В. П. Литвин, 1974]: динамику накопления антител, колебание бактерицидной активности крови, опсоно-фагоцитарную реакцию, специальную внутрикожную пробу и картину крови. В течение первых 10-15 дней жизни у телят более выражена клеточная защитная функция организма, характеризующаяся активным фагоцитозом микроорганизмов. Так, опсоно-фагоцитарный показатель по отношению к кишечной палочке в первые 10-15 дней жизни равнялся 8, в 30-40 дней — 3,7 а в 60-70 дней — 1,4. Сходные результаты получены при изучении гуморальных защитных факторов организма. Фагоцитарная способность лейкоцитов в крови играет большую роль в противомикробной защите новорожденного теленка. По данным исследователей, фагоцитарная активность гранулоцитов крови новорожденных телят до выпойки молозива летом (июнь-июль) достигает 98-100 % (у матерей 90-95 %). Фагоцитарное число (8-16) несколько ниже, чем в крови коров-матерей (16-19). Активность лизоцима по лизису тест-микроба в агаровом геле определяли в сыворотке крови телят и молозиве коров-матерей. Количество лизоцима в 5 первых порциях молозива равно 13±0,26 мкг/мл, в сыворотке крови новорожденных телят до выпойки молозива — 12±0,69, на 10-й день — 13,4±0,61 и в 3 месяца — 12,5±0,017. Таким образом, авторы не установили различий в содержании лизоцима в сыворотке крови телят в разных возрастных периодах [С. Н. Преображенский, Н. И. Блинов, Н. В. Ершова, Г. В. Вышинский, 1974].
В университете штата Оклахома, национальной лаборатории по изучению болезней животных в США и Гуэлфском университете Канада [O. Barta, V. Barta, 1972] на плодах и новорожденных телятах определяли бактерицидную активность крови. Двукратно разведенную сыворотку крови, в объеме 0,2 мл смешивали с равным объемом взвеси культуры содержащей 104 мл клеток бактерий кишечной палочки. Смесь выдерживали час при температуре 37° С и 0,1 мл ее высевали на плотную среду. Посевы выдерживали 16-18 ч при 37° С, подсчитывали число колоний и определяли дозу сыворотки, задерживающую рост 50 % бактерий. Антитела определяли по реакции агглютинации и пассивной гемагглютинации. Бактерицидная активность сыворотки крови составляла соответственно (в 1 мл сыворотки, задерживающей рост 50 %) бактерий у новорожденных телят 0,0036 и 0,0068.
При изучении иммунологической неспецифической реактивности организма новорожденных телят, [П. А. Емельяненко, 1979] определял фагоцитарную активность нейтрофилов и моноцитов крови, а также содержание в ее сыворотке лизоцима, комплемента и пропердина. Он установил, что у погибших телят от острых желудочно-кишечных болезней достоверно снижался процент фагоцитоза нейтрофилов, незначительно снижался уровень комплемента, пропердина и лизоцима.
При иммунобиологическом исследовании крови и сыворотки крови определяют [Д. Келмен, 1967; В. Я. Мозгис, 1982]:
фагоцитоз лейкоцитов по отношению к золотистому стафилококку 209р;
бактерицидную активность (БАСК);
активность бета-лизинов (БЛСК) сыворотки крови методом фотонефелометрии. Сущность метода заключается в том, что микробы (в данном случае, изолированный от телят штамм кишечной палочки — штамм 83) в течение 5 и 14 ч соответственно подвергаются воздействию исследуемой сыворотки. Чем слабее бактерицидная и бетализиновая активность сыворотки, тем интенсивнее размножаются микробы и тем более мутной становится взвесь;
лизоцимную активность сыворотки крови.
При круглогодовом стойловом содержании коров наступает угнетение иммунологической реактивности, а родившиеся от них телята имеют пониженный уровень гуморальных и клеточных факторов защиты организма (бактерицидная и лизоцимная активность сыворотки крови, фагоцитоз, уровень иммуноглобулинов, по сравнению с телятами, родившихся от коров, пользовавшихся выгулами и пастбищами [С. И. Плященко, В. Т. Сидоров, А. Ф. Трофимов, 1990].
Все новорожденные до приема молозива обладали достаточно выраженной фагоцитарной защитной реакцией. Показатели гуморальных факторов защиты формировались в более позднем возрасте и были выражены значительно слабее. До приема молозива сыворотка крови телят не обладала лизоцимной активностью и не содержала гаммаглобулинов [М. А. Сидоров, В. Н. Гущин, 1984].
На скорость роста телят положительное влияние оказывает 5-дневный период подсоса. Так, у таких телят были выше показатели естественной резистентности (лизоцимная активность, общий белок, глобулины, фагоцитарное число и фагоцитарный индекс) по сравнению с телятами, которых содержали с матерями только 12 ч [В. П. Павличенко, О. В. Милованов, Н. Н. Берникова, 1985].
Реактивность и адаптация сельскохозяйственных животных две взаимосвязанные проблемы; их надо изучать на организменном и иммуноцитологическом уровнях в комплексе со многими причинными факторами воздействующими на организм. Надо учитывать, что в каждом из указанных процессов лежат конкретные механизмы поддержания гомеостаза и адаптации, во многом еще не выясненные и требующие всестороннего изучения [А. Н. Голиков, 1989].
1.4. Получение и применение лактоглобулинов
В 1892 году P. Ehrlih впервые установил, что невосприимчивость к различным токсинам передается через молоко.
Исходя из того, что молозиво и молоко иммунизированных коров обладают выраженными превентивными свойствами, его использовали с профилактической и лечебной целью при соответствующих заболеваниях [C. Briggs, 1951; С. И. Губкин, А. И. Коган, 1971; J.E. Butler, C. Maxwele, 1972; P. Blackmer, 1973; D. M. Barber, 1978; D. M. Barber, 1979; J. J. Geene, 1984 В. Б. Билоштан, 1985; С. В. Вальциферова, 1997].
Из молока были получены иммунные лактоглобулины против сальмонелл, бруцелл, вируса ящура и полиомиелита [P. Lepine, 1963]. Эти препараты оказались пригодными для применения с лечебной и профилактической целью не только для телят, а и для обезьян. Автор приводит данные, подтверждающие преимущества применения лактоглобулинов по сравнению с цельными иммунными сыворотками молозива и крови.
Болгарские исследователи [К. Геров, П. Чушков, Р. Георгиева, 1965] изготовили из молозива препарат, который назвали лактоплазмин. По мнению авторов лактоплазмин содержит лактоглобулины, иммунные тела, витамины, микроэлементы и, являясь биологически ценным продуктом, может быть использован самостоятельно или в комбинации с другими средствами при заразных и незаразных болезнях молодняка, при которых понижается общая резистентность организма. Этот препарат является эффективным средством профилактики и лечения различных видов расстройств пищеварения у телят.
В ветеринарно-бактериологическом институте Бернского университета (директор профессор H. Fey) и научно-исследовательской лаборатории Бернской ветеринарной школы (заведующий Graub) из иммунного молозива изготовлен гамма-глобулиновый препарат (ГГП) или гамалин по Граубу [H. Fey, 1972].
Давая обзор работам, проведенным в этих учреждениях и анализируя литературу по методам получения и результатам изучения свойств ГГП, [K. Walser und H. Brumer, 1987] отмечают высокую терапевтическую и профилактическую эффективность применения этого препарата при инфекционных заболеваниях сельскохозяйственных животных. Они полагают, что наиболее результативным будет применение препарата при колиинфекциях и энтеротоксемиях.
Из молозива коров, иммунизированных внутривыменно против паратифа А. Ф. Барабаш, 1966, выделил путем осаждения сернокислым аммонием специфический противопаратифозный лактоглобулин. Полученный препарат применялся парентерально и показал выраженные профилактические и лечебные свойства на лабораторных животных и телятах.
Из молока коров, вакцинированных внутривыменно ящурным антигеном изготовили иммунолактон против вируса типа А, а затем изготовили бивалентный иммунолактон против вируса типа А и О [В. П. Онуфриев, В. К. Журавлёв, К. В. Чунаев, А. И. Дудников, Ю. Ф. Швецов, Б. П. Мартьянов, 1967]. Введение иммунолактона лабораторным животным, поросятам и телятам в дозе 0,5-1,5 мг на кг массы тела предохраняло животных от заболевания ящуром. Авторы указывают, что от одной коровы со средней продуктивностью 10 л молока в сутки в течение месяца можно изготовить иммунолактон в количестве, которое предохранит от ящура 300 телят или 800 поросят.
Иммунизируя стельных коров вначале подкожно концентрированным адсорбированным столбнячным анатоксином, а затем и внутривыменно нативным столбнячным анатоксином, получили сыворотку молозива с наличием противостолбнячных антитоксинов 850-2000 АЕ/мл, из которой выделили лактоглобулин, обладающий выраженными профилактическими и лечебными свойствами в опытах на белых мышах, крысах, ягнятах, поросятах и жеребятах [B. C. Барабаш, 1968].
Из сыворотки молозива коров, иммунизированных внутривыменно пуллорным антигеном, путем осаждения сернокислым аммонием получен специфический лактоглобулин, обладающий профилактическими свойствами в опытах на белых мышах и на цыплятах 10-30-дневного возраста [М. Г. Кондратюк, 1970].
Из сывороток молозива коров, гипериммунизированных паратифозной вакциной и бруцеллезным антигеном для РА, путем высаливания сернокислым аммонием при 20 % насыщении, выделен специфический паратифозный и бруцеллезный лактоглобулин, из которого изготовили люминисцируюшие лактоглобулины для диагностики паратифа телят и экспресс-индикации бруцелл [В. А. Атамась, 1968; В. А. Атамась, 1969].
Из сыворотки молозива коров, иммунизированных внутривыменно против диплококковой септицемии, получен специфический лактоглобулин и испытан с профилактической и лечебной целью в опытах на телятах. Противодиплококковый лактоглобулин обладал довольно выраженными профилактическими и лечебными свойствами [В. В. Май, 1970].
В ФРГ широко применяют гамма-глобулиновые препараты для профилактики и лечения колибактериоза и налажено промышленное производство гамма-глобулиновых препаратов из сыворотки убойных животных, сыворотки иммунизированных животных и молозива [R. Kruedener, 1970].
Из молока коров, гипериммунизированных внутривыменно модифицированным штаммом БУК вируса болезни Ауески получен иммунолактон против болезни Ауески [Д. Ф. Осидзе, В. К. Муравьев, В. Т. Ночевный, В. П. Онуфриев, В. М. Кравченко, 1972]. Авторы на основании проведенных исследований, установили принципиальную возможность диателической иммунизации коров штаммом БУК вируса болезни Ауески с целью получения специфической лактосыворотки и иммунолактона. Препарат в дозе 0,5-2,0 г/кг массы тела животного обладал выраженным профилактическим действием. Пассивный иммунитет у животных сохранялся 21 день. Лечебный эффект отмечали только после применения иммунолактона в первые двое суток после инфицирования животных.
Путем осаждения сернокислым аммонием молозивной сыворотки, был получен специфический лактоглобулин который обладал профилактическими и лечебными свойствами при экспериментальном и спонтанном заражении телят колибактериозом [Н. С. Жосан, 1974; Н. С. Жосан, 1976].
Для получения лактоглобулина [Е. В. Гублер, А. А. Генкин, 1978] использовали молозиво первого и второго удоев после отела коров. Молозиво подогревали до температуры 38-40° С, затем к нему добавляли свежеприготовленный раствор пепсина из расчета 100 мл на 900 мл молозива. После отделения молозивной сыворотки ее фильтровали через 4-5 слоев марли, затем через бумажный фильтр и консервировали 5 %-ным раствором фенола в 0,5 %-ном отношении к ее объему. Добавляли его при постоянном помешивании из расчета 11,1 мл на 100 мл препарата. Выпаивали лактоглобулин телятам в подогретом виде (38° С) в дозе 100-120 мл, первый раз за 30 мин до кормления молозивом, второй к концу первых суток. Больным животным после голодной диеты (8-9 ч) его давали утром до кормления один раз в сутки 2-3 дня в такой же дозе.
Из сыворотки крови отелившихся коров, находившихся в родильном отделении не менее 10 дней В. П. Павлов, Т. Н. Грязнева, Е. В. Чичикова, 1988, изготовили гаммаглобулин. Кровь брали у клинически здоровых животных (до 2 л от каждого). Из сыворотки крови готовили гаммаглобулин, используя 3 %-ный раствор этакридина лактата. Новорожденным телятам через каждые 3 дня инъецировали специфический гаммаглобулин внутримышечно в дозе 1 мл/кг массы тела, декстрофан и тривит в дозах соответственно 5 мл и 2 мл внутримышечно, а также в течение трех суток 2 раза в день внутримышечно вводили гентамицин в дозе 1,5 мг/кг массы, что позволило профилактировать желудочно-кишечные болезни у 100 % животных.
Для коррекции иммунодефицита неонатальных телят использовали колостральную сыворотку и лактоглобулин. [Н. С. Жосан, 1992]. Препараты вводили внутримышечно или подкожно, в дозе по 0,5 мл на кг живой массы дважды с интервалом 24 ч. Данные препараты повышали естественную резистентность организма, содержали неспецифические антитела против энтеропатогенов циркулирующих, в каждом конкретном хозяйстве при условии изготовления колостральной сыворотки и лактоглобулина, из молозива, полученного от коров данного хозяйства.
Применение молозива и его производных (молозивный иммуногормональный препарат, молозивная сыворотка, гидролизат казеина) снижает заболеваемость телят диареей на 50-60%, при этом болезнь протекает в легкой форме со 100% выздоровлением. Парентеральное введение этих препаратов новорожденным телятам способствует повышению уровня белка в сыворотке крови в 1,5 раза, иммуноглобулинов в 2 раза, уровень лейкоцивот у этих животных не превышал 8000, бактериальная загрязненность фекалий была в 2-3 раза ниже, чем у телят, которым молозиво давали только внутрь. Максимальный лечебно-профилактический эффект молозивная терапия дает в том хозяйстве, где молозиво было получено [В. В. Бурсуков, 1993; С. В. Вальциферова, 1997].
Исходя из литературных данных, следует отметить, что иммунное молозиво, молоко и полученные из них препараты обладают выраженными профилактическими лечебными свойствами.
Сравнительная дешевизна сырья, используемого для получения препаратов из молозива (практически оно не используется в хозяйствах) [М. Х. Шайхаманов, В. П. Грамолин, Б. М. Авакаянц, 1993], простота изготовления, высокая эффективность послужит основанием для получения указанных препаратов в производственных условиях.
заключение
Анализируя данные литературы, касающиеся состояния естественной резистентности и иммунологической реактивности у новорожденных телят при колибактериозе, необходимо выделить следующие основные моменты:
колибактериоз занимает среди заболеваний новорожденных животных одно из ведущих мест и наносит значительный экономический ущерб;
высокий уровень резистентности обеспечивает широкий диапазон ответных реакций организма на влияние различных факторов внешней среды в том числе и микроорганизмов, позволяющих ему адаптироваться на уровне физиологической реактивности;
животные с низким уровнем неспецифической резистентности не способны адаптироваться к изменившимся условиям внешней среды, что часто вызывает истощение резервной защитно-приспособительной реактивности и, как следствие этого, приводит к гибели животных;
сущность защитных факторов против колибактериоза телят ассоциируется с лактоглобулиновой фракцией и уровнем К-антител-агглютининов в сыворотке молозива коров и в сыворотке крови новорожденных телят;
фагоцитирующие лейкоциты, лизоцим, комплемент, пропердин и иммуноглобулины обладают функцией антител, противомикробное действие которых в совокупности выражается бактерицидной активностью сыворотки крови;
уровень иммуноглобулинов является одним из важных показателей резистентности организма новорожденного. При этом следует определять и учитывать иммунный статус до первой выпойки молозива и после его скармливания;
абсорбция колостральных иммуноглобулинов находится в прямой зависимости от ряда факторов важнейшими из которых являются: способность новорожденных телят усваивать их из молозива, условия внешней среды, качество молозива и своевременная выпойка первого молозива в адекватном количестве;
лактоглобулин совместно с ретинолом отличаются высоким иммуно-корректирующим эффектом и широтой профилактического действия;
аминазин совместно с Т-активином обладают иммуно-корректирующим действием и антисекреторной активностью ингибирующей трансформацию аденозинтрифосфата (АТФ) в циклический 3,5-аденозинмонофосфат (АМФ).
РАЗДЕЛ 2 РЕЗУЛЬТАТЫ СОБСТВЕННЫХ ИССЛЕДОВАНИЙ
2.1. Материалы и методы исследований.
2.1.1.Общая методика. Экспериментальная часть работы выполнялась на ферме крупного рогатого скота учебно-опытного хозяйства «Кетросу», района Анений Ной, ОПХ «Колоница» района Криулень и в лаборатории микробиологии кафедры эпизоотологии Государственного aграрного университета Молдовы с 1973 по 1998 год.
Опыты проводились на коровах черно-пестрой породы 5-8 лактации и на родившихся от них телят, с целью изучения состояния неспецифической и специфической иммунологической реактивности новорожденных телят при колибактериозе, а также выявления роли кислотно-основного баланса в течение инфекционного процесса, коррекции иммунодефицитного состояния новорожденных телят. Для лечения применяли антисекреторные препараты, ингибирующие циклический аденозинмонофосфат.
Пробы молозива отбирались в первый день (из первого, второго и третьего доения), на второй, пятый день после отела, а на десятый, двадцатый день и через месяц исследовалось молоко.
В сыворотке крови, молозиве и молоке изучались: динамика накопления агглютининов, иммуноглобулиновый уровень, в т. ч. в динамике.
Сыворотку крови получали путем отстаивания, а из молозива — методом пепсинизации. Из сывороток молозива первого и второго дня после отела выделяли колостральную сыворотку и лактоглобулин. У родившихся телят от опытных и контрольных групп коров в сыворотке крови изучали: динамику накопления О- и К- антител-агглютининов, уровень иммуноглобулинов в зависимости от клинического статуса, показатели фагоцитоза, пропердина, комплиментарной и бактерицидной активности (до поения молозивом, на 2, 5, 10, 20 и 30 день жизни).
Бактерицидные свойства колостральной сыворотки и сыворотки крови новорожденных телят изучали нефелометрическим методом. Профилактические и лечебные свойства молозивных сывороток и лактоглобулина изучались на лабораторных животных и телятах.
Коррекцию иммунодефицита неонатальных телят, проводили с применением лактоглобулина и витамина А.
В качестве анти-секреторных факторов, ингибирующих циклический аденозинмонофосфат (сАМР) применяли хлорпромазин и Т-активин.
Опыты проводились на 562 телятах, 468 коровах, 247 белых мышах и 30 кроликах.
Полученный цифровой материал подвергался биометрической обработке по различным биометрическим методикам: [Дж. У. Снедекор, 1961; И. П. Ашмарин, А. А. Воробьев, 1962; В. Ю. Урбах, 1964; Е. В. Гублер, А. А. Генкин А. А., 1969].
2.1.2. Приготовление колибактериозного антигена. Для приготовления антигена использовали три серотипа колибактерий: О78:К80, О119:К69 и О137:К79.
Использованные серотипы Е. coli по морфологическим и биохимическим свойствам были типичными, отвечающими требуемым иммуногенным, антигенным н вирулентным свойствам.
Е. coli О78:К80, О119:К69 и О137:К79 раздельно высевали в пробирки с мясопептонным бульоном на 12 ч и проверяли на чистоту (микроскопия мазков, окрашенных по Граму). В последующем бульонную культуру высевали на стерильный мясопептонный агар в матрацах. Посевы выращивали в термостате в течение 18 ч при температуре 37° С, проверяли на чистоту и смывали стерильным изотоническим раствором хлорида натрия. Для выделения О-антигена бактериальную суспензию автоклавировали при 120°С в течение 2 ч дня разрушения К-антигена. Густую отмытую суспензию Е. coli содержащую 10 млрд. микробных тел в 1 мл по оптическому стандарту, смешивали с ацетоном в отношении 1:5 до появления коагуляции (свертывания). Плотную часть материала отбирали на воронке Бюхнера, промытой один раз ацетоном и затем высушенной эфиром. Препарат высушивали на воздухе и хранили при 4°С.
Сухой порошок применяли для приготовления суспензии антигена, добавляя 1 мг высушенных ацетоном бактерий на 1 мл барбиталового буфера.
Для выделения К-антигена бактериальную суспензию получали путем смыва из агаровых культур изотоническим раствором хлорида натрия, содержащего 0,5 % формальдегида. Бактерии были экстрагированы при добавлении ацетона непосредственно после их смыва с агаровой культуры и промыты. [E. Neter, E. A. Gorzynski, R. M. Gino, O. Westphal and O. Luderwitz. 1956; E. Neter, 1957].
Полученные О- и К-антигены проверяли на стерильность и на безвредность. Стерильность устанавливали путем высева на МПБ, МППБ и МПА. Посевы выдерживали 10 дней в термостате (+37°С). Безвредность антигенов была проверена на лабораторных животных. Для этого каждый из антигенов вводили 10 белым мышам под кожу в области спины по 0,5 мл и 5 морским свинкам в области внутренней поверхности задней конечности по 1,0 мл. Клинические наблюдения за опытными животными вели в течение 15 дней.
Отсутствие роста в посевах на МПБ, МППБ и МПА и гибели привитых животных позвонило считать антигены стерильными и безвредными. Хранились антигены при температуре +4 +5°С в холодильнике.
2.1.3. Изучение агглютиногенных свойств серотипов О78:К80, О119:К69 и О137:К79 Е. coli. Для определения агглютиногенности изучаемых серотипов использовали 30 кроликов, по 5 на каждый серотип (определение О- и К-антигенов). Антигены вводили кроликам в краевую вену уха в нарастающих дозах (от 0,5 до 2,0 мл) с интервалом в три дня, четырехкратно.
Для определения О-антигена смешивали одну каплю антигена с одной каплей соответствующей О-сыворотки на зеркальном стекле, размещали над водяной баней при 60°С. Результаты учитывали через 10 мин после смешивания.
Позитивную реакцию подтверждали пробирочной агглютинацией, применяя формалинизировавную шестичасовую культуру в МПБ. Основное разведение сыворотки 1:10.
Для определения К-антигена культуру вначале тестировали пластинчатой реакцией агглютинации на предметном стекле. Одну каплю живой суспензии культуры смешивали с одной каплей различных антисывороток. Результаты учитывали в течение 30с после смешивания. Позитивную реакцию подтверждали пробирочной агглютинацией до титра тест сыворотки, применяя как антиген живую пятичасовую культуру в МПБ. Пробирки инкубировали при 37°С два часа, в последующем агглютинационные пробирки центрифугировали при 2000 об/мин в течение трех минут. Одинаковый писк агглютинации учитывали как позитивную реакцию. [E. Neter, E. A. Gorzynski, R. M. Gino, O. Westphal and O. Luderwitz. 1956; E. Neter, 1957].
2.1.4. Получение сыворотки крови, молозива и молока. Кровь от животных (коров, телят) брали из яремной вены в стерильные пробирки и помещали в термостат (+37°С) на 3 ч, после чего выдерживали 12-16 ч при комнатной температуре до полного отделения сыворотки. Затем сыворотку отсасывали в стерильные пробирки и помещали в холодильник (+4 +5°С).
Молозиво от коров брали при первом, втором и третьем доении после отела, а затем на второй, пятый, десятый, двадцатый дни и через месяц.
Сыворотку из молозива и молока получали путем добавления пепсина. На каждые 1000 мл молозива или молока добавляли 100 мл 0,5% раствора пепсина и после тщательного перемешивания смесь помещали в водяную баню при температуре 38° С на четыре часа. Образовавшийся сгусток, отделяли от сыворотки фильтрацией через три слоя марли. Полученную сыворотку пропускам через воронку Бюхнера с двойным слоем бедой фильтровальной бумаги, а затем через бактериальный фильтр Зейтца.
2.1.5. Реакция атглютинации с молозивной и молочной сыворотками. В центрифужные пробирки наливали 5-6 капель 5 % раствора пепсина, приготовленного на изотоническом растворе хлорида натрия и 10 мл исследуемого молозива или молока, тщательно перемешивали и для свертывания ставили в термостат при температуре 38°С на 30 мин. Свернувшееся молозиво отделяли от стенок пробирки стеклянной палочкой и центрифугировали при 3000 об/мин в течение 30 мин.
С полученной сывороткой ставили реакцию агглютинации в объеме 1 мл в разведениях с физиологическим раствором от 1:10 до предельного титра сыворотки. Контроли обычные.
Антиген добавляли по 0,05 мл в каждую пробирку, пробы встряхивали и помешали в термостат при температуре 37°С на 4 ч, после чего производили предварительный учет реакции, а окончательный учет реакции определяли через 24 ч. Молозиво исследовали в день его получения.
2.1.6. Определение титра агглютининов. Для изучения динамики накопления агглютининов в сыворотках крови и молозива использовали реакцию агглютинации, которую ставили в объеме 1 мл (классическим методом) в пробирках с ровным округлым дном с убитыми и живыми О- и К-антигенами (серотипы О78:К80; О119:К69; О137:К79 Е. coli ).
Для определения агглютинационных титров, в наших исследованиях применяли двукратное разведение сывороток. Разведение сывороток крови и молозива производили в агглютинационных пробирках начиная с разведения 1:10.
После добавления антигена (по 0,05 мл) содержимое пробирок встряхивали, затем помещали в термостат на 4 ч при температуре 37°С. После этого производили предварительный учет результатов реакции. В дальнейшем пробирки выдерживали 24 ч при комнатной температуре и определяли окончательный результат.
Для учета результатов реакции агглютинации пользовались четырех крестовой системой обозначения. За агглютинационный титр принимали то наибольшее разведение сывороток, при котором еще наблюдалась видимая агглютинация, оцениваемая в четыре креста.
2.1.7. Определение активности лизоцима в сыворотке крови. Сыворотку крови, отделяли общепринятым методом. Предварительно готовили 1/15М фосфатный буфер (рН 6,2), для чего использовали два исходных раствора: 4,539 г х. ч. КН>2>РО>4> растворенный в 500 мл дистиллированной воды и 2,375 г х. ч. Nа>2>НРО>4>×12Н>2>O, растворенный в 200 мл дистиллированной воды. При смешивании этих растворов в определенном соотношении, получали рН буфера 6,2.
Для приготовления 1 % агара «Дифко» на 1/15М фосфатном буфере брали 1 г сухого агара, помещали в колбу на 200 мл и заливали 99 мл 1/15М фосфатного буфера. Колбу закрывали ватно-марлевой пробкой, помещали в кипящую баню и выдерживали 25-30 мин.
Расплавленный агар охлаждали до 60-70°С и вносили в него ацетоновый порошок Micrococcus lysodeikticus из расчета 10-20 мг на 100 мл объема. Необходимое количество порошка перед внесением в агар суспендировали в 3-5 мл 1/15М фосфатного буфера. Полученную смесь перемешивали до получения однородной взвеси.
Полученную смесь разливали в чашки Петри с таким расчетом, чтобы подучить толщину слоя 4 мм.
После застывания агара в нем при помощи тонкостенной трубочки вырезали лунки диаметром 5 мм на расстоянии 2-3 см от краев чашки и друг от друга. Для подсыхания лунок чашки помещали в термостат приоткрытыми на 60 мин. Исследуемые пробы сыворотки крови разводили соответственно 1/15М фосфатным буфером 1:10, 1:2 и 1:5. Каждую пробу после разведения заливали в отдельную лунку в объеме 0,05 мл.
Параллельно с исследуемым материалом 1/15М фосфатным буфером разводили стандартный кристаллический лизоцим так, чтобы получить его растворы с концентрацией 0,5 мкг/мл, 1; 3; 5; 10; 20; 40; 60; 80 и 100 мкг/мл. По 0,05 каждого разведения стандартного лизоцима разливали по отдельным лункам.
Чашки Петри с исследуемым материалом и стандартным лизоцимом выдерживали 48 ч во влажной камере при комнатной температуре (22-24°С).
После экспозиции при помощи линейки измеряли диаметр зон лизиса Micrococcus lysodeikticus вокруг лунок с исследуемым материалом и стандартным лизоцимом.
Первоначально на полулогарифмической бумаге строили калибровочную кривую, используя результаты, полученные при измерении зон лизиса вокруг лунок с раствором стандартного лизоцима в различных концентрациях. Для этого значения, отражающих диаметры зон лизиса субстрата в миллиметрах, откладывали по оси абсцисс, а показатели концентрации стандартного лизоцима в мкг/мл, вызывающих лизис по оси ординат. Точки пересечения первых и вторых значений соединяли между собой, при этом получалась прямая линия.
Расчет результатов проводили следующим образом. Диаметр зоны лизиса Micrococcus lysodeikticus вокруг лунки с сывороткой крови составил 9,5 мм. На калибровочном графике этому значению соответствовала концентрация 2,5 мкг/мл стандартного лизоцима. Сыворотку крови перед исследованием разводили 1:10 1/15М фосфатным буфером. Следовательно, концентрация лизоцима в исследуемой пробе сыворотки крови равнялась 2,5×10=25 мкг/мл. Так как, литическая активность стандартного лизоцима изменяется в результате изготовления и хранения, полученное абсолютное значение выражали в единицах относительной активности. Активность стандартного лизоцима составляла 20600 ед/мг или 20,6 ед/мк, активность лизоцима исследуемой пробы сыворотки молозива равнялась 20,6×25 =515,0 ед/мл.
2.1.8. Определение содержания комплемента в сыворотке крови.
Исследовали сыворотку крови, полученную из яремной вены животных. Предварительно готовили веронал-мединаловый буфер: 0,575 г веронала растворяли в 50 мл дистиллированной воды, подогретой до 60° С, охлаждали и добавляли 8,5 г хлорида натрия, 0,375 г мединала, 0,5 мл раствора содержащего 1 М MgCl>2> и 0,3 М CaCl>2> (100 мл Н>2>О + 19 г MgCl>2>×6Н>2>О г + 6 г CaCl>2>). Объем доводили до 200 мл дистиллированной водой. Буфер хранили в холодильнике. Перед употреблением буфер разводили дистиллированной водой (1:4). Физиологический раствор: 8,5 г хлорида натрия растворяли в 1 л дистиллированной воды и фильтровали.
Эритроциты барана. Кровь у барана брали из яремной вены в колбу с бусами и встряхивали в течение 10-15 мин для предотвращения свертывания. Дефибринированную кровь фильтровали через двойной слой марли. Эритроциты барана отмывали 3 раза центрифугированием по 10 мин при 3000 об/мин. Из отмытых эритроцитов готовили 5 % взвесь на физиологическом растворе и стандартизовали на ФЭК. С этой целью эритроциты лизировали: к 1 мл отмытых эритроцитов добавляли 9 мл дистиллированной воды. Лизированные эритроциты фотоколориметрировали при зеленом светофильтре против воды. Оптическая плотность лизата — 1,08 при длине 541 мкм в кювете 10 мм.
Для определения титра гемолизина, готовили вначале основное разведение гемолитической сыворотки 1:100 (0,1 мл сыворотки + 9,9 мл буфера), затем из этого разведения поучали разведения от 1:1000 до 1:4000.
|
1:1000 |
1:1200 |
1:1500 |
1:2000 |
1:2500 |
1:3000 |
1:3500 |
1:4000 |
|
|
Буфер, мл |
0,9 |
1,1 |
1. 4 |
1,9 |
2,4 |
2,9 |
3,4 |
3,9 |
|
Гемолизин |
0,1 |
0,1 |
0,1 |
0,1 |
0,1 |
0,1 |
0,1 |
0,1 |
Гемолитическую систему готовили следующим образом. К стандартизованной 5 % взвеси эритроцитов барана медленно, при постоянном помешивании, добавляли равный объем гемолизина в тройном титре. Так, если полный гемолиз наблюдался в пробирке с разведением гемолитической сыворотки 1:1200, то для приготовления гемолитической системы брали разведение 1:400 (0,1 мл цельной гемолитической сыворотки и 39,9 мл буфера). Пробирки с гемолитической системой встряхивали и инкубировали при 37° С в течение 1ч для сенсибилизации эритроцитов барана.
Исследуемую сыворотку, разводили 1:10, разливали в 10 пробирок и доводили буферным раствором (физиологическим) до 1,0 мл. После этого в каждую пробирку добавляли 1 мл гемолитической системы, то есть сенсибилизированных эритроцитов барана, 2-я пробирка — контрольная.
|
Исследуемая сыворотка (1:10), мл |
0,05 |
0,1 |
0,15 |
0,2 |
0,25 |
0,3 |
0,35 |
0,4 |
0,45 |
0,5 |
|
Буферный раствор |
0,95 |
0,9 |
0,85 |
0,8 |
0,75 |
0,7 |
0,65 |
0,6 |
0,55 |
0,5 |
|
Сенсибилизированные эритроциты барана |
1,0 |
1,0 |
1,0 |
1,0 |
1,0 |
1,0 |
1,0 |
1,0 |
1,0 |
1,0 |
Пробирки инкубировали при 37° С в течение 45 мин, охлаждали в холодильнике 10 мин, центрифугировали при 1500 об/мин и колориметрировали на ФЭК в кюветах (10мм) с зеленым светофильтром против воды и определяли процент гемолиза.
Для определения активности комплимента в гемолитических единицах готовили шкалу и кривую гемолиза. Для приготовления шкалы гемолиза к 1 мл стандартной 5 % взвеси эритроцитов барана прибавляли 3 мл дистиллированной воды в результате чего происходил гипотонический лизис эритроцитов (100 % гемолиз). Пробирку центрифугировали (при 1500 об/мин 10 мин) и готовили шкалу гемолиза:
|
100 % гемолиз эритроцитов на дистиллированной воде |
0,1 |
0,2 |
0,3 |
0,4 |
0,5 |
0,6 |
0,7 |
0,8 |
0,9 |
1,0 |
|
Буферный раствор, мл |
0,9 |
0,8 |
0,7 |
0,6 |
0,5 |
0,4 |
0,3 |
0,2 |
0,1 |
0 |
|
Гемолиз, % |
10 |
20 |
30 |
40 |
50 |
60 |
70 |
80 |
90 |
100 |
Полученные разведения и исходный раствор колориметрировали на ФЭК в кюветах (1 мм) и строили график зависимости коэффициентов экстинции от процента гемолиза. На оси абсцисс откладывали процент гемолиза, на оси ординат — показатели экстинции, соответствующие данному проценту гемолиза.
Расчет 50 % гемолитических единиц комплемента в 1 мл проводили следующим образом. После титрования исследуемой сыворотки, инкубации ее при 37° С в течение 45 мин, выдержки в течение 10 мин в холодильнике при 4° С и центрифугирования пробирку из шкалы гемолиза с одной 50 % единицей комплемента (№5) сравнивали с пробирками из ряда титрования сыворотки. Если содержимое пробирки №5 соответствовало по цвету пробирке №6, из ряда титрования комплемента исследуемой сывороткой, то есть дозе комплемента 0,3 мл, то расчет вели следующим образом.
0,3 мл сыворотки (разведение 1:10) соответствует одной 50 % гемолитической единице комплемента, а в 1 мл исследуемой сыворотки содержится:
0,3 мл (1:10) — 1
1,0 мл — Х
Х
= = 3,33 ел/мл.
= 3,33 ел/мл.
2.1.9. Определение содержания пропердина в сыворотке крови. Для определения титра пропердина исследуемую сыворотку разводили мединал-вероналовым буфером (рН 7,2-7,4), начиная с 1:10 до 1:320. В центрифужные пробирки вносили по 0,2 мл каждого разведения сыворотки, до 0,2 мл суспензии инулина (соответствует 3 мг сухого вещества) и по 0,2 мл комплемента в рабочем титре. В контрольный ряд пробирок вместо инулина добавляли по 0,2 мл буфера. Все пробы (опытные и контрольные) помещали на 1 ч в термостат при 37° С, после чего центрифугировали, переносили супернатант в отдельные пробирки по 0,3 мл и добавляли по 0,2 мл стандартной гемолитической системы. Реакцию учитывали после 20-минутной экспозиции при 37° С. За титр пропердина принимали то наибольшее разведение сыворотки, в котором наблюдается полная задержка гемолиза, и в каком разведении в контрольном ряду отличается полный гемолиз. Данные рассчитывали по формуле:
|
Разведение, в котором отмечен полный гемолиз в контроле |
— |
Разведение, в котором отмечена полная задержка гемолиза в опыте |
×10 |
|
Разведение, в котором отмечен полный гемолиз в контроле |
Полученные результаты выражали в ед/мл. При полной задержки гемолиза в разведении 1:35 в опыте, а в контроле — полный гемолиз в разведении 1:50, содержание пропердина в 1 мл испытуемой сыворотки составляет:
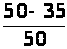 ×10
= 3 ед/мл.
×10
= 3 ед/мл.
Тестированные показатели: комплементарную, лизоцимную, пропердиновую активность, а также содержание иммуноглобулинов в сыворотке крови, молоке и молозиве крупного рогатого скота определяли по методам В. Г. Дорофейчук, 1968; X. Я. Грант, Л. И. Яворский, И. А. Блумберг, 1973; И. М. Архангельский, 1976; В. Н. Андреев, Г. И. Подопригора, 1977; Е. С. Фортинская, А. М. Наумова, Е. А. Маркова, 1977; О. Н. Грызлова, П. А. Емельяненко, В. Н. Денисенко, 1978; Э. Бем, 1979; П. А. Емельяненко, О. И. Грызлова, Г. Н. Печникова и М. Н. Тулупова, 1980; Э. С. Коган, 1981; Г. В. Павлов, Г. Н. Печникова, Смолянская- О. О. Суворова, 1988; В. В. Биктимиров, 1993.
При определении лизоцима, учитывая замедленную активность фермента крупного рогатого скота, приводили тщательную подготовку материалов к исследованию, чтобы избежать ошибок, связанные с действием на тест-микробы других литических факторов исследуемой пробы.
При тестировании комплементарной активности сыворотки крови тщательно осуществляли подбор индикаторной системы чувствительной к комплементу крупного рогатого скота.
Для определения пропердина использовали модифицированный метод предложенный П. А. Емельяненко и др., 1980.
2.1.10. Изучение бактерицидной активности сывороток крови. Для определения бактериостатической и бактерицидной активности сывороток крови О. В. Смирнова, Т. А. Кузьмина (1966) предложили фотонефелометрический метод.
В нашей работе мы учитывали изменения оптической плотности питательной среды с микробами и питательной среды с испытуемой сывороткой крови и микробами сразу после соединения и через 3-, 5-, 7-, 9-, 12- и 24-часовой инкубации.
Для нефелометрического метода чистую культуру Е. coli высевали на МПА и выращивали в термостате при температуре 37° С в течение 24 ч. Затем смывали стерильным изотоническим раствором хлористого натрия и стандартизовали до содержания в 1 мл 2 млрд. микробных тел. Из этой взвеси производили посев на МПБ в пробирки и сутки выращивали в термостате при вышеуказанной температуре.
Для определения бактерицидной активности сывороток крови мы использовали питательную среду (обогащенный пептоном бульон Хоттингера), содержащую 200 мг% амминного азота.
В стерильные кюветы с рабочей длиной 10 мм стерильной пипеткой разливали по 4,5 мл бульона Хоттингера, затем добавляли по 0,5 мл испытуемой сыворотки крови и суточную бульонную культуру Е. coli по одной бактериологической петле (диаметр петли 4 мм).
Контрольные кюветы заполняли теми же компонентами, что и опытные с той лишь разницей, что в один кювет вместо сыворотки добавили 0,5 мл стерильного изотонического раствора хлорида натрия. Содержимое всех кювет тщательно перемешивали стерильной стеклянной палочкой, после чего все кюветы закрывали ватно-марлевыми пробками. Оптическую плотность среды определяли на фотоэлектроколориметре ФЭК-М, используя зеленый светофильтр. После этого кюветы ставили в термостат при температуре 37° С. Повторно определяли оптическую плотность содержимого кювет через 3, 5, 7, 9, 12 и 24 ч. Установку «0» на ФЭК-е производили с помощью кюветы, заполненной дистиллированной водой. Оценку бактерицидной активности сыворотки крови проводили по формуле:
А=100-
 ×100,
×100,
Где: А — бактерицидная активность (в %);
Д — оптическая плотность;
Т — время экспозиции кювет в термостате (в часах).
Затем вычисляли среднюю «напряженность бактерицидной активности» (НБА) по формуле:
Средняя
НБА =
 ,
,
Где: НБА — средняя напряженность бактерицидной активности молозивной сыворотки;
А>1>, А>2>, А>3>.....А>n> — бактерицидная активность сыворотки молозива через 3, 5, 7, 9, 12 и 24ч (в %);
Т>1>, Т>2>, Т>3>....... Т>n> — продолжительность термостатирования (в часах).
2.1.11. Постановка опсонофагоцитарной реакции. При постановке опсонофагоцитарной реакции руководствовались методикой, описанной В. Я. Мозгис (1982). Для приготовления антигена культуру Е. coli выдерживали в термостате при температуре 37° С в течение 18-24 ч в МПБ, а затем на МПА в течение 24 ч. Проверяли на чистоту и смывали стерильным изотоническим раствором хлорида натрия. По оптическому стандарту доводили концентрацию до 2-х млрд. микробных тел в 1 мл. Приготовленную взвесь нагревали в водяной бане при 70° С в течение 30 мин.
Для опсонофагоцитарной реакции применяли только суточную агаровую культуру Е. coli.
Исследования проводили в следующей последовательности. В пронумерованные стерильные пробирки наливали до 0,5 мл 2 % прокипяченного и охлажденного раствора лимоннокислого натрия, 1 мл крови от исследуемых телят и тщательно смешивали, сюда же вносили по 0,5 мл антигена. После осторожного перемешивания пробирки помещали в термостат при 38° С на 3 мин, предварительно погрузив их на 2-3 мин в водяную баню с температурой 38° С. Затем пробирки из термостата извлекали, готовили мазки, сушили на воздухе, нумеровали, фиксировали в метиловом спирте в течение 5 мин и окрашивали по Романовскому-Гимза.
Для приготовления рабочего раствора краски, дистиллированную воду усредняли фосфатными буферами: 1. Двухосновной фосфат натрия (Na>2>HPO>4>×12H>2>O) — 17,814 г на 1 л (рН 8,302). 2. Одноосновной фосфат калия (KH>2>PO>4>) — 13,638 г на 1 л (рН 4,529). Если к литру дистиллированной воды прибааляли по 5 см3 того и другого раствора, то рН такой воды был равен 6,813.
Краску готовили ex tempore из расчета 1,5 капли краски на 1 мл усредненной воды с рН 6,813. Мазки окрашивали в течение 45 мин, промывали водопроводной водой и высушивали на воздухе. Окрашенные мазки исследовали под иммерсионной системой микроскопа и окуляра 7×.
Подсчет фагоцитированных микробов производили в 100 нейтрофилах. На основе подсчета определяли фагоцитарную интенсивность (среднее число поглощенных микробов одним нейтрофилом) и фагоцитарную активность (процент фагоцитирующих нейтрофилов). С кровью подопытных животных поставили 450 реакций.
2.1.12. Определение содержания иммуноглобулинов в сыворотке крови, молозиве и молоке крупного рогатого скота.
Предварительно готовили фосфатный буфер 0,03М, рН 3,0. С этой целью 10,74 г натрия фосфорнокислого двух замещенного (Na>2>HPO>4>×12H>2>O) и 5,84 г хлорида натрия помещали в мерную колбу, растворяли в дистиллированной воде и доводили объем до 1 л. Подобным образом готовили раствор из 4,08 г однозамещенного фосфорнокислого калия (KH>2>PO>4>) и 5,84 г хлорида натрия.
Растворы под контролем потенциометра смешивали в соотношении обеспечивающим рН буфера 8,0.
Пробы сывороток крови получали путем выдержки взятой крови из яремной вены животных 20-30 мин при 37° С в термостате. После отделения образовавшегося сгустка от стенок пробирок стеклянной палочкой, кровь помещали на 16-20 ч в холодильник. На следующий день сыворотку отсасывали в стерильную центрифужную пробирку, центрифугировали 15-20 мин при 3000 об/мин.
Пробы молозива и молока после взятия выдерживали 16-20 ч при 4° С и центрифугировали в течение 1 ч при 6000 об/мин. Затем снимали верхний слой жира, отсасывали надосадочную жидкость, которую и использовали для определения.
Пластинки из 3 %-ного агара «Дифко», смешанного с антисывороткой готовили следующим образом: 360 мг агара помещали в колбу, заливали 12 мл буфера и прибавляли 0,25 мл 1 %-ного раствора мертиолата. Колбу помещали в баню с холодной водой, которую затем нагревали до кипения и выдерживали смесь до полного расплавления агара.
После расплавления гель охлаждали до 56° С и к нему приливали равный объем антисыворотки в рабочем разведении и снова нагревали до температуры 56° С. Смесь тщательно перемешивали и выливали на стекло, помещенное на столике с уровнем в строго горизонтальном положении.
После застывания геля в нем делали лунки диаметром 2 мм на расстоянии 15 мм друг от друга.
При определении содержания иммуноглобулина G пробы сыворотки крови разводили буфером рН 8,0 в 20 и 30 раз. Пробы молозива первого дня лактации в 100, 50 и 20 раз, пробы молозива последующих дней лактации — в 50, 20 и 10 раз, использовали не разведенные и разведенные в 10 раз пробы молока.
При определении содержания иммуноглобулина М пробы сыворотки крови и молозива разводили в 10 и 5 раз, пробы молока использовали не разведенными и разведенными в 5 раз.
При определении содержания иммуноглобулина А пробы сыворотки крови использовали не разведенными, пробы молозива и молока — не разведенными и разведенными в 5 и 2 раза.
Пробы сыворотки крови получали от новорожденных телят до приема молозива во всех случаях использовали не разведенными.
Подготовленные пробы крови, молозива, и молока вносили в лунки геля с антисывороткой с помощью пастеровской пипетки. При этом лунку заполняли полностью, не переливая пробы за ее пределы.
Параллельно с опытными пробами на каждом стекле ставили контроль — разливали в лунки стандартный препарат определенного иммуноглобулина. При этом его разводили буферным раствором с таким расчетом, чтобы в 1 мл раствора препарата содержалось 5,0; 2,0; 1,0; 0,5; 0,2 и 0,1 мл белка. Таким образом, на каждом стекле получалось 6 лунок с контрольной пробой препарата.
После заполнения лунок, стекла помешали в эксикатор с водой и выдерживали при комнатной температуре, при определении содержания иммуноглобулинов G и А в течение 24 ч, а при определении иммуноглобулина М — 48 ч.
По окончании сроков инкубации стекла извлекали из эксикатора и измеряли диаметр кольца преципитата вокруг лунок с помощью линейки Беринг-Верке, после окрашивания агаровых пластин. С этой целью стекла с агаром погружали на двое суток в буфер. В течение этого срока буфер трижды заменяли новой порцией. После отмывания от не участвующих в реакции веществ, пластинки агара высушивали при комнатной температуре под фильтровальной бумагой. Затем пластинки (без фильтровальной бумаги) помещали в 0,1 %-ный раствор амидо-черного 10В на 3 ч, промывали трехкратно раствором уксусной кислоты и высушивали.
Для приготовления 0,1 %-ного раствора амидо-черного 10В, брали 1 г краски, растворяли в 100 мл ледяной уксусной кислоты (СН>3>СООН), после чего объем раствора доводили дистиллированной водой до 1 л. Раствор хранили в темной посуде при комнатной температуре. Раствор уксусной кислоты для промывания: брали 70 мл ледяной уксусной кислоты и доводили объем дистиллированной водой до 1 л.
Для расчета содержания иммуноглобулинов в испытываемых пробах строили калибровочную кривую на полулогарифмической бумаге: на оси ординат откладывали концентрацию белка в пробах стандарта, по оси абсцисс — диаметр кольца преципитации. Калибровочную кривую строили отдельно по каждому иммуноглобулину определенного класса.
Количество иммуноглобулина в испытуемой пробе определяли путем сравнения диаметра кольца преципитации вокруг ее лунки с калибровочной кривой [G. Mancini, A. O. Carbonara and J.F. Heremans, 1965; У. Дж. Герберт, 1974; В. М. Чекишев, 1977; П. А. Емельяненко, О. И. Грызлова, Г. Н. Печникова и М. Н. Тулупова, 1980; P. В. Петров, 1987; Н. С. Жосан, 1989; Н. В. Матузенко, Е. В. Андреев, А. И. Собко, 1990].
2.1.13. Экспресс-метод для определения содержания иммунных глобулинов. Кровь получали от телят из яремной вены до приема молозива и через 24 ч после рождения. Сначала ее выдерживали около часа в (30-35° С), а затем в холодильнике 10-12 ч. После чего отделяли сыворотку в отдельные пробирки.
Раствор цинк сульфата (208 мг/л ZnSO>4>×7H>2>O) готовили в мерной колбе на дистиллированной воде, которую выдерживали 4-5 дней и затем кипятили в течение 10-15 мин с целью растворения диоксида углерода.
Для приготовления стандарта мутности брали 3 мл раствора хлорида бария (BaCl>2>×2H>2>O) в 100 мл дистиллированной воды, вносили в мерную колбу на 100 мл и доводили до метки 0,2N раствором серной кислоты. Полученный раствор соответствует 20 ед. цинк сульфатному тесту [A. D. MсEwan, E. W. Fisher, I. E. Selman and W. J. Penhale, 1970].
Калибровочную таблицу для калориметрического определения содержания иммуноглобулинов на ФЭК-56М получали путем разведения стандарта мутности.
Для определения иммунных глобулинов подбирали пробирки одинакового диаметра и наливали в каждую по 6 мл цинк сульфатного раствора, а в контрольною пробирку — 6 мл дистиллированной воды. Во все опытные пробирки (по количеству образцов) и в контрольную добавляли по 0,1 мл исследуемой сыворотки и выдерживали при комнатной температуре (20° С) 60 мин. После экспозиции пробирки взбалтывали, чтобы обеспечить одинаковое перераспределение осадка и помещали в фотоэлектроколориметр, который предварительно устанавливали на «0» по дистиллированной воде. Измерение проводили на ФЭК-56М, ври длине волны 400±5 нм (синий светофильтр) в кюветах толщиной слоя 10 мм. Показание прибора ФЭК-56М определяли образцах сыворотки через контрольную пробирку умножением различия на фактор 10. 20 единиц цинк сульфатного теста помутнения были эквивалентны соответственно 20-30 мг/мл содержания колостральных иммунных глобулинов в сыворотке крови теленка [A. D. MсEwan, E. W. Fisher, I. E. Selman and W. J. Penhale, 1970].
2.1.14. Определение уровня пассивной защиты у новорожденных телят. Метод основан на том, что белки сыворотки крови могут преципитироваться различными солями, включая сульфит натрия (Na>2>SO>3>). Этот феномен зависит от концентрации соли и молекулярной характеристики белков. Готовили 14-, 16-, и 18 %-ный растворы Na>2>SO>3>, используя для этих целей безводный реактив. Соответственно 14, 16 и 18 г его растворяли в 100 мл дистиллированной воды. Кроме безводного сульфита натрия использовали Na>2>SO>3>×7Н>2>О, при этом количество его для приготовления соответствующих концентраций увеличивали вдвое.
В пробирки вносили по 1,9 мл раствора сульфита натрия каждой концентрации и добавляли по 0,1 мл испытуемой сыворотки. Пробирки тщательно встряхивали и выдерживали 1 ч при комнатной температуре. Реакцию считали отрицательной, если преципитат не образуется и положительной, если видны хлопья преципитированного белка или заметно даже небольшое его присутствие, в данном случае уровень иммуноглобулинов в испытуемой сыворотке соответствует промежуточному значению. Концентрация иммуноглобулина меньше 5 мг/мл соответствовала помутнению при концентрации 18 %; от 5 до 15 мг/мл — 16 и 18 %, и больше 15 мг/мл — 14, 16 и 18 % сульфита натрия. [N. E. Pfeiffer, T. C. McGuire, 1977; N. E. Pfeiffer, T. C. McGuire, R. E. Bendel and J. M Weikel, 1977; Ю. Н. Федоров, 1988].
Сывороточный гемолиз не влиял на результаты преципитации иммуноглобулинов.
2.1.15. Выделение лактоглобулина из колостральной сыворотки. Лактоглобулины из сыворотки молозива получали солевым методом. За основу взяли методику фракционного высаливания противогриппозных сывороток.
К сыворотке добавляли два или три объема дистиллированной воды в зависимости от содержания белка. В смесь, при перемешивании мешалкой, добавляли тонкой струей насыщенный раствор сернокислого аммония. После тщательного перемешивания смесь оставляли в покое на 10-15 ч в холодильнике до полного формирования осадка.
Лактоглобулин сыворотки молозива подвергался высаливанию 38 объемными процентами насыщенного раствора сернокислого аммония по формуле:
Х=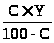 ,
,
где: Х — объем насыщенного раствора сернокислого аммония;
С — требуемая концентрация сернокислого аммония;
Y — объем разбавленной молозивной сыворотки.
рН насыщенного раствора сернокислого аммония — 7,3-7,5.
Выпавший осадок лактоглобулина отделяли от раствора путем фильтрации через воронку Бюхнера с двойным слоем хроматографической бумаги.
Плотный осадок переносили в целлофановые мешочки и подвергали диализу в проточной водопроводной воде, а затем в дистиллированной воде до тех пор, пока в растворе не обнаруживалось даже следов сернокислого аммония, что проверяли реактивом Несслера.
По окончании диализа определяли концентрацию белка (рефрактометрически) в растворе лактоглобулина и дистиллированной воды разбавляли его до содержания 10 % белка. К раствору добавляли хлорид натрия до изотонического насыщения.
Подученный препарат подвергали стерилизующей фильтрации через фильтр Зейтца с пластинкой СФ и расфасовывали по ампулам и флаконам.
Стерильность полученного препарата подтверждали высевом на МПБ, МПА и среду Китт-Тарроци, а безвредность на белых мышах и морских свинках. Посевы выдерживали в термостате при 37° С восемь суток. Лактоглобулин вводили подкожно двум морским свинкам массой 230-240 г в дозе по 3 мл и четырем белым мышам массой 16-18 г по 0,5 мл с последующим наблюдением за животными в течение 5 дней.
Исключение примесей балластных белков в препарате производили электрофоретически. Препарат хранили в холодильнике при температуре +3 +5° С.
2.2. Изучение специфической иммунологической реактивности телят в постнатальный период.
Уровень иммуноглобулинов в сыворотке крови телят до приема молозива и через 24 ч после рождения определяли цинк-сульфатным методом [A. D. MсEwan, E. W. Fisher, I. E. Selman and W. J. Penhale, 1970].
Концентрацию иммуноглобулинов молозива коров с учетом их возраста вычисляли по уровням линейной регрессии [H. Balbierz, M. Nicolaijczuk, J. Zeilinski, 1983].
Эффективность колостральных иммуноглобулинов изучали по отношению содержания иммуноглобулинов в сыворотке крови неонатальных телят к количеству поступивших с молозивом.
Количественную характеристику абсорбции колостральных иммуноглобулинов классов G, М и А устанавливали методом радиальной иммунодиффузии [G. Mancini, A. O. Carbonara and J.F. Heremans, 1965; В. М. Чекишев, 1977]. Выделенные иммуноглобулины классов G, М и А служили антигенами для иммунизации кроликов [R. A. Wilson and I. W. Jutila, 1976] с целью получения соответствующих моноспецифических антисывороток.
Степень катаболизма иммуноглобулинов изучали определением их классов в сыворотке крови методом радиальной диффузии. Иммуноглобулины в фекалиях определяли по E. F. Logan, W. J. Penhale, 1972; Р. П. Маслянко, 1982.
После измерения диаметров зон преципитации концентрацию иммуноглобулинов рассчитывали математическим способом, исходя из прямо пропорциональной зависимости между ней и квадратом диаметра кольца [Э. Бем, 1979].
Колостральную сыворотку отделяли после добавления сычужного фермента. Лактоглобулин получали добавлением к сыворотке молозива насыщенного раствора сульфата аммония с последующим диализом против 0,01М рН 7,6 Na>2>HPO>4>; KН>2>PO>4>.
Статистическую обработку результатов исследования проводили с использованием критерия достоверности Стьюдента.
2.2.1. Уровень иммуноглобулинов в сыворотке крови телят до приема молозива и через 24ч после рождения и взаимосвязь между концентрацией иммуноглобулинов и проявлением синдрома нарушения пищеварения. Проведенные исследования показали, что концентрация колостральных иммунных глобулинов в крови телят до приема молозива составила 0,18±0,08 мг/мл, этот уровень главным образом связан с IgМ. Через 24 часа после приема молозива содержание иммуноглобулинов достигло 23,94±1,71 мг/мл или увеличилось в 133 раза (табл. 2.1).
Таблица 2.1
Содержание иммунных глобулинов в сыворотке крови новорожденных телят (n=20).
|
Группы телят |
Средняя арифметическая, мг/мл |
Дисперсия |
Среднее квадратическое отклонение |
Средняя арифметическая ошибка |
Точность прямых измерений |
Доверительный интервал, мг/мл |
Относительная ошибка (погрешность), % |
|
До приема молозива |
0,18 |
0,0035 |
0,019 |
0,008 |
0,02 |
0,16-0,20 |
11,1 |
|
Через 24 ч после рождения: I группа — контроль |
23,94 |
14,69 |
3,83 |
1,71 |
4,75 |
18,19-28,69 |
19,8 |
|
II группа — энтеритная форма колибактериоза |
18,26 |
0,16 |
0,4 |
0,18 |
0,49 |
17,77-18,75 |
2,68 |
|
III группа — септическая форма колибактериоза |
15,37 |
0,66 |
0,43 |
0,19 |
0,53 |
14,84-15,90 |
1,24 |
Новорожденные телята с содержанием иммуноглобулинов 23,94± 1,71 мг/мл (доверительный интервал 19,19-28,69 мг/мл) переболели легкой формой колибактериоза на третьи сутки. Даже такая высокая концентрация иммуноглобулинов не обеспечивала иммунологический статус новорожденных телят, что связано со значительной инфицированностью внешней среды и снижением вследствие этого защитного уровня молозива.
Содержание иммуноглобулинов через 24 часа в сыворотке крови телят сильно варьирует и составляет от 19,85 мг/мл до 29,11 мг/мл. Такая разница объясняется количеством проглоченного теленком молозива.
Способность новорожденных животных адаптироваться к изменениям внешней среды лимитировано и изменение условий, не влияющих на взрослых животных, может плохо отразиться на состояние здоровья телят. Известно, что стресс является способствующим фактором возникновения колибактериоза у телят. Причиной стресса могут быть различные нарушения условий содержания, порой самые безобидные, поэтому предотвратить стресс весьма трудно. Даже перемещение является причиной стресса, который приводит к увеличению уровня заражения инфекционными агентами. Низкая температура, сквозняки, влажная подстилка способствуют возникновению и распространению инфекции. Следовательно, иммунологический статус, необходимый для защиты организма, зависит от количества молозива, проглоченного теленком в первые минуты жизни и условий внешней среды.
Исходя из этого, невозможно точно указать количество колостральных иммуноглобулинов, обеспечивающих иммунологическую защиту животных.
При концентрации иммуноглобулинов в сыворотке крови телят 18,26±0,18 мг/мл (доверительный интервал 17,77-18,75 мг/мл) заболевание протекает с расстройством пищеварения,и сопровождается средней тяжестью дегидратации а также диареей. Такие телята отказываются от молозива, с трудом поднимаются, задняя часть туловища испачкана фекалиями, влажная.
Длительность течения болезни при такой концентрации иммуноглобулинов обычно составляет 4-6 дней. Уровень иммуноглобулинов в сыворотке крови телят с энтеритной формой колибактериоза ниже, чем в контрольной группе (Р<0,05).
В сыворотке крови телят III группы концентрация иммуноглобулинов была самой низкой и составляла 15,37±0,19 мг/мл при доверительном интервале 14,82-15,90 мг/мл. По сравнению с контрольной группой изменения в содержании иммуноглобулинов статистически достоверны (Р<0,01). У животных отмечалась септическая форма колибактериоза, депрессия, дегидратация. Течение болезни проявляется профузным поносом, усиленной перистальтикой кишечника, глаза запавшие, телята не способны были стоять на ногах и погибали в 7-10 дневном возрасте. Существенное уменьшение концентрации иммунных глобулинов в данной группе животных ассоциировалось с тяжелой степенью дегидратации и выраженной диареей.
Коэффициент вариации в исследуемых сыворотках колебался от 2,29 % во 2-й опытной группе до 15,9 % в контрольной.
При бактериологическом исследовании ректальных тампонов и патологического материала выделили вирулентные штаммы Е. coli. Пробы брали до лечения в течение 12 часов с начала диареи.
Исходя из результатов исследований, рекомендуем при содержании иммуноглобулинов в сыворотке крови телят суточного возраста 18,26±0,18 мг/мл задавать взамен молока дважды в день 2 л изотонического раствора глюкозы, хлорида натрия и антибактериальные препараты. При концентрации иммуноглобулинов 15,37±0,19 мг/мл, парентерально вводить кровь, сыворотку крови, сыворотку молозива (внутривенно) и антибактериальные препараты. Применение антибиотиков и изотонических растворов глюкозы и хлорида натрия при септической форме колибактериоза не является эффективным, так как выживание телят в таких случаях зависит от высокой концентрации иммуноглобулинов в сыворотке крови.
2.2.2. Количественная характеристика абсорбции колостральных иммуноглобулинов новорожденными телятами. Известно, что здоровье и выживаемость новорожденных, связаны с поступлением в их организм молозива. Природа защитных свойств молозива была предметом длительного изучения, вследствие чего установлено, что одним из определяющих этих свойств факторов являются иммуноглобулины. Иммуноглобулины молозива обеспечивают защиту от так называемой «местной» микрофлоры, так как в результате самопроизвольной антигенной стимуляции этой микрофлорой в организме матерей всегда имеются соответствующие антитела.
Всасывание иммуноглобулинов происходит в кишечнике, и через лимфатические пути они попадают в кровь. Иммуноглобулины появляются в лимфе уже через 1-2 ч после поступления их в двенадцатиперстную кишку животных. Факторы, обеспечивающие всасывание нерасщепленных молекул белка, как и причины, по которым впоследствии оно становится невозможным, до настоящего времени изучены недостаточно. J.W.Boyd (1972) указывает на существование специальных внутриклеточных рецепторов узнавания, обеспечивающих селективную передачу белков в кровь. Исследование механизмов транспорта и катаболизма иммуноглобулинов in vitro показало, что на первых этапах всасывания формируется комплекс между иммуноглобулинами и специфическими рецепторами энтероцитов. Во взаимодействии с рецепторами клеток кишечника участвует Fc — фрагмент иммуноглобулинов. По-видимому, связывание иммуноглобулинов с клетками предохраняет их от катаболизма и способствует транспорту в кровь [И. И. Головистиков, 1979].
Высокий уровень иммуноглобулинов в организме новорожденных обеспечивается поступлением необходимого количества молозива в оптимальные сроки. Низкое содержание иммуноглобулинов в крови новорожденных животных связано с недостаточным или слишком поздним скармливанием молозива, низким содержанием иммуноглобулинов в нем, а также нарушением абсорбции.
Проведенные нами исследования показали, что концентрация иммунных глобулинов в молозиве подопытных коров изменялась по дням лактации. Так, уровень иммунных глобулинов в молозиве коров первого дня лактации составил 57,02±0,86 мг/мл при доверительном интервале 55,09-58,95 мг/мл. На второй и третий день концентрация иммуноглобулинов составила соответственно 26,42±0,06 и 11,56±0,42 мг/мл при доверительном интервале 26,29-26,55 и 10,40-12,72 мг/мл (табл. 2.2).
Таблица 2.2
Содержание иммунных глобулинов в молозиве подопытных коров (n=19)
|
Дни после отела |
Средняя арифметическая, мг/мл |
Дисперсия |
Среднее квадратическое отклонение |
Средняя арифметическая ошибка |
Точность прямых измерений |
Доверительный интервал, мг/мл |
Относительная ошибка (погрешность), % |
|
Первый |
57,02 |
5,28 |
2,29 |
0,86 |
1,93 |
55,09-58,95 |
3,38 |
|
Второй |
26,42 |
0,96 |
0,16 |
0,06 |
0,13 |
26,29-26,55 |
0,49 |
|
Третий |
11,56 |
0,86 |
0,93 |
0,42 |
1,16 |
10,40-12,72 |
10,03 |
При изучении концентрации иммунных глобулинов в сыворотке крови телят и в молозиве их коров-матерей установлено, что абсорбция зависит от уровня иммунных глобулинов в молозиве коров, количества принятого молозива, задержки первого кормления, а также от рН молозива.
Так, при содержании иммунных глобулинов в молозиве первого дня 59,00±0,59 мг/мл при доверительном интервале 57,36-60,64 мг/мл, уровень колостральных иммуноглобулинов в сыворотке крови телят равнялся 23,096±1,00 (рис. 2.1) при доверительном интервале 20,32-25,87 мг/мл.
При концентрации иммуноглобулинов в молозиве 57,02±0,86 и 54,53±0,57 мг/мл (доверительный интервал соответственно равен 55,09-58,95 и 53,25-55,81 мг/мл; содержание колостральных иммунных глобулинов в сыворотке крови телят также изменяется и составляло 18,1±0,11 и 15,06±0,12 мг/мл при доверительном интервале соответственно 17,85-18,35 и 14,79-15,33 мг/мл (табл. 2.3).
Таблица 2.3
Содержание иммунных глобулинов в молозиве коров первого дня лактации (n=19).
|
Группы |
Средняя арифметическая, мг/мл |
Дисперсия |
Среднее квадратическое отклонение |
Средняя арифметическая ошибка |
Точность прямых измерений |
Доверительный интервал, мг/мл |
Относительная ошибка (погрешность), % |
|
Первая (контрольная) |
59,00 |
1,66 |
1,29 |
0,59 |
1,64 |
57,36-60,64 |
2,78 |
|
Вторая (опытная) |
57,02 |
5,28 |
2,29 |
0,86 |
1,93 |
55,09-58,95 |
3,38 |
|
Третья (опытная) |
54,53 |
2,28 |
1,51 |
0,57 |
1,28 |
53,25-55,81 |
2,35 |
При изучении эффективности абсорбции колостральных иммуноглобулинов новорожденными телятами нами установлено, что количественная оценка всасывания иммуноглобулинов в контрольной группе составляет 39,21±2,04 % при доверительном интервале 33,55-44,87 процентов.
В первой опытной группе (энтеритная форма колибактериоза) и во второй (септическая форма колибактериоза) этот показатель соответственно равнялся 31,78±0,11 % доверительный интервал 31,53-32,03 % и 27,63±0,23 % при доверительном интервале 27,11-28,15 % (табл. 2.4).
При бактериологическом исследовании ректальных тампонов и посмертного материала выделили вирулентные штаммы Е. coli . Пробы брали до лечения в пределах 12 часов с начала диареи.
Таблица 2.4
Содержание иммунных глобулинов в сыворотке крови новорожденных телят (n=19).
|
Группы |
Средняя арифметическая, мг/мл |
Дисперсия |
Среднее квадратическое отклонение |
Средняя арифметическая ошибка |
Точность прямых измерений |
Доверительный интервал, мг/мл |
Относительная ошибка (погрешность), % |
|
Контрольная |
23,096 |
4,89 |
2,21 |
1,00 |
2,77 |
20,32-25,87 |
12,02 |
|
Первая опытная (энтеритная форма колибактериоза) |
18,1 |
0,078 |
0,28 |
0,11 |
0,25 |
17,85-18,35 |
1,38 |
|
Вторая опытная (септическая форма колибактериоза) |
15,06 |
0,10 |
0,31 |
0,12 |
0,27 |
14,79-15,33 |
1,79 |
Следовательно, количественная оценка абсорбции колостральных антител характеризует иммунологический статус новорожденных телят в нашем случае она составляет соответственно 39,21±2,04 % (контрольная группа), 31,78±0,11 (энтеритная форма колибактериоза) и 27,63±0,23 % (септическая форма колибактериоза) (табл. 2.5; рис. 2.2). Таким образом, установлена прямая зависимость между концентрацией иммуноглобулинов в сыворотке крови новорожденных животных и их резистентностью.
Таблица 2.5
Показатели абсорбции колостральных иммуноглобулинов новорожденными телятами через 24 ч после рождения (n=19)
|
Группы |
Средняя арифметическая, мг/мл |
Дисперсия |
Среднее квадратическое отклонение |
Средняя арифметическая ошибка |
Точность прямых измерений |
Доверительный интервал, мг/мл |
Относительная ошибка (погрешность), % |
|
Первая (контрольная) |
39,21 |
20,18 |
4,49 |
2,04 |
5,66 |
33,55-44,87 |
14,43 |
|
Вторая (опытная) |
31,78 |
1,69 |
0,28 |
0,11 |
0,25 |
31,53-32,03 |
0,78 |
|
Третья (опытная) |
27,63 |
0,37 |
0,61 |
0,23 |
0,52 |
27,11-28,15 |
1,88 |

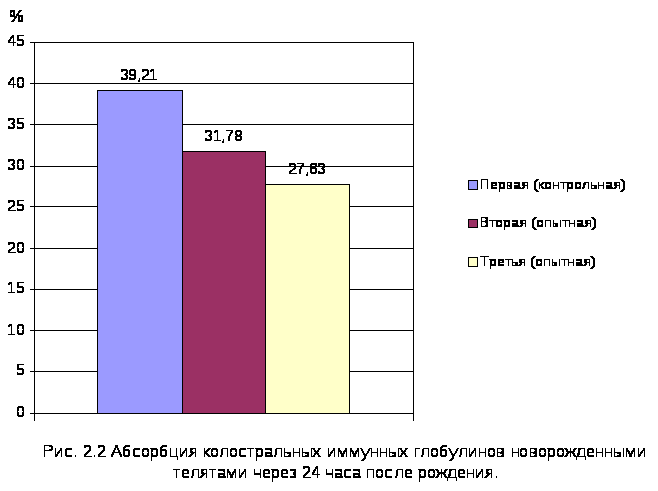
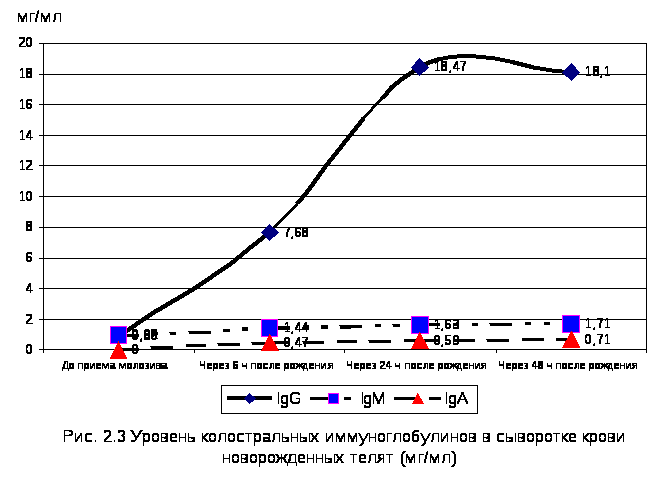
2.2.3. Уровень колостральных иммуноглобулинов в сыворотке крови новорожденных телят через 6, 24 и 48 ч после рождения. Иммуноглобулины, обнаруженные в сыворотке крови телят до приема молозива, считаются важнейшими защитными факторами для этих животных. Как правило, это было незначительное количество IgG и IgM (та6л. 2.6). Тем не менее, в отдельных случаях перед приемом молозива наблюдали очень высокий уровень IgM (5,6 мг/мл) и IgG (4,2 мг/мл). IgА не обнаруживался в пробах до приема молозива. В сыворотке крови всех телят устанавливали уровень трех классов иммуноглобулинов в течение 6 ч от рождения, и их максимум достигал к 48 часам. Большое отклонение в иммуноглобулиновом уровне было заметно у, отдельных животных. Двое телят имели уровень IgG 9,4 и 14,42 мг/мл в течение 24 ч от рождения. В противоположность у одного теленка этот показатель равнялся 34,2 мг/мл, что, по-видимому, связано с внутриматочной инфекцией. IgМ более эффективно абсорбируется (63,8 %), чем IgА (47 %) или IgG (11,19 %) в течение 6 ч после рождения. Механизм торможения абсорбции макромолекул последовательный и прогрессирующий, который воздействует на все три класса иммуноглобулинов вскоре после рождения. Установленное снижение в абсорбции различимо между классами и указывает, что механизм торможения независимо действует для каждого класса. Прекращение абсорбции было более быстрым для IgG и медленным для IgА с промежуточной позицией для IgМ. Следовательно, каждый класс иммуноглобулинов абсорбируется независимо, а сам абсорбционный процесс следует рассматривать как неспецифический.
Таблица 2.6
Концентрация колостральных иммуноглобулинов в сыворотке крови новорожденных телят (n=7), мг/мл
|
Иммуноглобулины |
Уровень иммуноглобулинов |
Доверительный интервал |
Р< |
|
До приема молозива |
|||
|
G |
0,86±0,32 |
0,14-1,58 |
|
|
M |
0,92±0,34 |
0,16-1,68 |
|
|
A |
0 |
- |
|
|
Через 6 ч после рождения |
|||
|
G |
7,68±0,90 |
5,66-9,70 |
0,01 |
|
M |
1,44±0,32 |
0,72-2,16 |
0,05 |
|
A |
0,47±0,05 |
0,36-0,58 |
|
|
Через 24 ч после рождения |
|||
|
G |
18,47±0,53 |
17,28-19,66 |
0,01 |
|
M |
1,63±0,29 |
0,98-2,28 |
0,05 |
|
A |
0,59±0,16 |
0,23-0,95 |
0,05 |
|
Через 48 ч после рождения |
|||
|
G |
18,10±0,54 |
16,89-19,31 |
0,05 |
|
M |
1,71±0,25 |
1,16-2,27 |
0,05 |
|
A |
0,71±0,06 |
0,58-0,84 |
0,05 |
2.2.4. Количественное определение колостральных иммуноглобулинов в сыворотке крови и в фецес неонатальных телят. Данные табл. 2.7 характеризуют уровень иммуноглобулинов в сыворотке крови и в копрофильтратах у телят, больных колибактериозом.
Таблица 2.7
Количественные показатели иммунных глобулинов в сыворотке крови и копрофильтратах телят при колибактериозе (Х±S>x>), мг/мл
|
Группы телят |
Содержание иммуноглобулина |
Р — значение |
||
|
Сыворотка крови (1) |
Копрофильтраты (2) |
1 (Р<) |
2 (Р<) |
|
|
Павшие (n=4) |
Значение павших к выжившим |
|||
|
IgG |
6,88±0,19 |
5,17±0,03 |
0,01 |
0,01 |
|
IgМ |
0,31±0,02 |
0,27±0,01 |
0,01 |
0,05 |
|
IgА |
0,43±0,02 |
0,13±0,01 |
0,01 |
0,01 |
|
Общий Ig |
7,62±0,023 |
5,57±0,05 |
0,01 |
0,01 |
|
Выжившие (n=4) |
Значение выживших к здоровым |
|||
|
IgG |
9,73±0,02 |
7,20±0,02 |
0,01 |
0,01 |
|
IgМ |
0,75±0,02 |
0,30±0,01 |
0,01 |
0,01 |
|
IgА |
0,39±0,01 |
0,43±0,01 |
0,01 |
0,01 |
|
Общий Ig |
11,37±0,05 |
7,93±0,04 |
0,01 |
0,01 |
|
Клинически здоровые (n=5) |
Значение клинически здоровых к павшим |
|||
|
IgG |
15,56±0,05 |
11,42±0,02 |
0,001 |
0,001 |
|
IgМ |
1,78±0,01 |
0,76±0,01 |
0,001 |
0,001 |
|
IgА |
1,48±0,01 |
0,58±0,02 |
0,001 |
0,001 |
|
Общий Ig |
18,82±0,07 |
12,76±0,05 |
0,001 |
0,001 |
Существенное различие была установлено как в общих иммунных глобулинах, так и в отдельных классах иммуноглобулинов. Активность катаболизма имеет существенные различия. Так, сывороточная концентрация IgG у павших телят была в 2,26 раза меньшей, чем у клинически здоровых, концентрация IgМ и IgА также у павших была соответственно в 5,74 и 3,44 раза меньшей.
Общий иммуноглобулиновый уровень был выше в контрольной группе в 2,47 раза по сравнению с группой павших телят. Уровень катаболизма IgG, IgМ и IgА в группе павших телят составил соответственно 24,86, 12,91 и 69,77 %. В группе выживших телят этот показатель равнялся соответственно 26,01, 60,00 и 51,69 %. Уровень катаболизма общих иммуноглобулинов в группе выживших телят был в 1,12 раза выше, чем в группе павших телят, а IgМ — в 4,64 раза.
2.2.5. Концентрация иммуноглобулинов в сыворотке крови и секрете молочной железы коров. В результате проведенных исследований установлено, что концентрация иммунных глобулинов в сыворотке крови и в секрете молочной железы имеет существенное различие (табл.2. 8; рис. 2.4). Так, содержание иммуноглобулинов различных классов значительно варьирует, особенно заметно превалирует концентрация IgG в сыворотке крови и в секрете молочной железы по сравнению с IgМ и IgА. Количественное содержание IgG в молозиве составляет 49,5±0,21 мг/мл при доверительном интервале 49,27-49,73 мг/мл, тогда как концентрация IgМ и IgА равняется соответственно 4,16±0,16 мг/мл (доверительный интервал 3,98-4,34 мг/мл) 3,93±0,28 мг/мл (доверительный интервал 3,65-4,21 мг/мл).
Содержание IgG в сыворотке крови равняется 18,28±0,27 мг/мл при доверительном интервале 18,0-18,56 мг/мл, в то время как концентрация IgМ и IgА составляет 2,54±0,07 мг/мл (доверительный интервал 2,46-2,62 мг/мл) и 0,49±0,02 мг/мл (доверительный интервал 0,46-0,52).
Таблица 2.8
Концентрация иммуноглобулинов в сыворотке крови и секрете молочной железы коров (n=18)
|
Иммуноглобулины |
Концентрация (мг/мл) |
Процент к общему уровню иммуноглобулинов |
||||
|
Сыворотка |
Сыворотка |
|||||
|
крови |
молозива |
молока |
крови |
молозива |
молока |
|
|
IgG |
18,28±0,27 |
49,5±0,21 |
0,59±0,03 |
85,78 |
85,96 |
76,62 |
|
IgМ |
2,54±0,07 |
4,16±0,17 |
0,05±0,01 |
11,92 |
7,22 |
6,50 |
|
IgА |
0,49±0,02 |
3,93±0,28 |
0,13±0,01 |
2,30 |
6,82 |
16,88 |
|
Общий Ig |
21,31±0,36 |
57,59±0,66 |
0,77±0,05 |
100 |
100 |
100 |
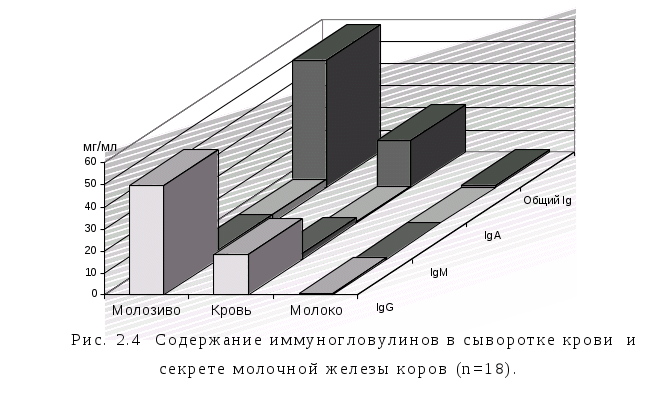
При изучении количественного содержания иммунных глобулинов в сыворотке молока установлено значительное превышение IgG (0,59±0,03 мг/мл, доверительный интервал 0,56-0,62 мг/мл) в сопоставлении с уровнем IgМ (0,05±0,01 мг/мл, доверительный интервал 0,04-0,06 мг/мл) и IgА (0,13±0,01 мг/мл, доверительный интервал 0,12-0,14 мг/мл).
Процентное отношение содержания иммунных глобулинов в сыворотке крови и секрета молочной железы и общему уровню иммуноглобулинов составляет соответственно в сыворотке крови IgG — 85,78; IgМ — 11,92 и IgG — 2,3 %. В сыворотке молозива эти показатели равняются: IgG — 85,96; IgМ — 7,25; IgА — 6,82 %.
Процентное отношение в сыворотке молока к общему уровню иммуноглобулинов составляли: IgG — 76,62; IgМ — 6,50; IgА — 16,88 %.
2
.2.6.
Содержание иммуноглобулинов в сыворотке
крови коров и новорожденных телят.
Гуморальный
иммунитет обуславливается специфическими
антителами, принадлежащими к пяти
классам иммуноглобулинов, основными
из которых являются три: IgG, IgМ, IgА.
Известно также, что в неблагополучном
по инфекционным заболеваниям хозяйстве
даже при наличии комплекса стрессовых
факторов (нарушение условий содержания,
режима кормления) часть животных не
заболевают или переболевают бессимптомно,
что объясняется иммунологической
резистентностью животных, определяемой
уровнем иммуноглобулинов.
В связи с этим количественное определение отдельных классов иммуноглобулинов, как метод исследования состояния гуморального иммунитета приобретает в настоящее время особую значимость в определении иммунологической резистентности новорожденных животных.
Большинство исследователей септическую форму колибактериоза телят связывают с недостаточностью иммуноглобулина М, а энтеритную — иммуноглобулинов G и А. Такую роль различных классов иммуноглобулинов следует учитывать при разработке профилактических мероприятий, диагностике и лечении.
В практике большое значение имеет своевременная оценка иммунологического статуса новорожденных животных, основанная на поступлении иммуноглобулинов молозива. Для этих целей используют такие методы, как радиальная иммунодиффузия в агаре, рефрактометрия, спектрофотометрия, осаждение иммуноглобулинов сульфатом цинка или аммония и сульфитом натрия, с последующей нефелометрией или колориметрированием [R. Aschaffenburg, S. Bartlett, S. K. Kon, P. Terry, S. Y. Thompson and D. M. Walker, 1949a; R. Aschaffenburg, S. Bartlett, S. K. Kon, D. M. Walker, C. Briggs, E. Cotchin and R. Lovell, 1949b; G. Mancini, A. O. Carbonara and J.F. Heremans, 1965; В. М. Чекишев, 1977; P. Blackmer, 1973].
В результате проведенных исследований установлено, что концентрация иммуноглобулинов в сыворотке крови телят до приема молозива является незначительной и общее содержание их составляет 1,03 мг/мл. В суточном возрасте общее содержание иммуноглобулинов в сыворотке крови телят в 1,38 раза меньше, чем у коров сразу после отела и составляет соответственно 16,9 и 22,3 мг/мл.
По мнению многих ученых в условиях крупных ферм новорожденным телятам в суточном возрасте необходимо иметь в сыворотке крови столько иммуноглобулинов, сколько их имеется в сыворотке взрослых и если этого не обеспечить, то желудочно-кишечная патология неизбежна [B. C. Шипилов, В. П. Шишков, В. Г. Зароза, В. П. Карев, Г. Д. Смоленская, 1987].
У исследованных нами телят суточного возраста концентрация IgМ в сыворотке крови была меньшей соответственно в 1,8 и 1,7 раза, чем у коров (табл. 2.9). Незначительное увеличение количества иммуноглобулинов отдельных классов в сыворотке крови телят 2-3-дневного возраста (IgG — 15,35 мг/мл, IgМ — 1,11 мг/мл, IgА — 0,50 мг/мл и соответственно 15,84 мг/мл, 41,82 мг/мл, 0,70 мг/мл) объясняется тем, что в первые один-два дня жизни через кишечник могут проходить иммуноглобулины молозива в неизмененном виде.
Увеличение концентрации иммуноглобулинов в сыворотке крови телят 9-10-дневного возраста происходит в связи с тем, что спустя неделю после рождения слизистая оболочка тонкого кишечника приобретает способность синтезировать собственные защитные иммуноглобулины, так называемые секреторные [D. Eddie, M. Sohulkind, I. Robbins, 1971; P. D. Porter, D. E. Noakes, and W. D. Allen, 1972].
Таблица 2.9
Содержание иммунных глобулинов в сыворотке крови телят
|
Определяемая величина |
Число исследований |
Содержание иммуноглобулинов, мг/мл |
Общее содержание, мг/мл |
||
|
G |
М |
А |
|||
|
Сыворотка крови коров сразу после отела |
4 |
20,55 |
1,965 |
0,4 |
22,915 |
|
Сыворотка крови телят до приема молозива |
4 |
0,5 |
0,495 |
0,0375 |
1,0325 |
|
Сыворотка крови телят суточного возраста |
4 |
15,35 |
1,115 |
0,5275 |
16,9925 |
|
Сыворотка крови телят 2 3 дневного возраста |
4 |
15,84 |
1,8225 |
0,705 |
18,3675 |
|
Сыворотка крови телят 9 10 дневного возраста |
4 |
16,81 |
2,0275 |
0,77 |
19,6075 |
Эти данные составляют соответственно по сравнению с 2-З-дневным, возрастом IgG — 15,84 мг/мл, IgМ — 1,82 мг/мл, IgА — 0,70 мг/мл против IgG — 16,81 мг/мл, IgМ — 2,03 мг/мл, IgА — 0,77 мг/мл. Статистическая обработка результатов определения количества иммуноглобулинов отдельных классов в сыворотке крови коров и телят представлена в таблицах 2.10; 2.11; 2.12; 2.13. Данные таблиц показывают, что относительная ошибка опытов составляет от 2,96 до 16,3 %, точность прямых измерений от 0,006 мг/мл IgА до 1,9 мг/мл IgG; средняя арифметическая ошибка определения иммуноглобулинов в сыворотке крови телят составляет: IgG от 0,0015 до 0,27; IgМ — 0,024-0,19; IgА — 0,006-0,0048; дисперсия соответственно IgG от 0,0007 до 0,0036; IgG — 0,01-0,017; IgА — 0,01-0,06.
Таблица 2.10.
Содержание иммунных глобулинов в молозиве коров первого дня лактации (n=4)
|
Определяемая величина |
Средняя арифметическая, мг/мл |
Дисперсия |
Среднее квадратическое отклонение |
Средняя арифметическая ошибка |
Р |
tp |
Точность прямых измерений |
Доверительный интервал, мг/мл |
Относительная ошибка (погрешность), % |
|
IgG |
20,55 |
1,09 |
1,04 |
0,6 |
0,05 |
3,182 |
1,91 |
18,64-22,46 |
9,3 |
|
IgМ |
1,965 |
0,03 |
0,18 |
0,1 |
0,05 |
3,182 |
0,32 |
1,64-2,28 |
16,3 |
|
IgА |
0,4 |
0,0003 |
0,017 |
0,01 |
0,05 |
3,182 |
0,03 |
0,37-0,43 |
7,5 |
Таблица 2.11
Динамика накопления иммуноглобулина G в сыворотке крови телят (n=16).
|
Определяемая величина |
Средняя арифметическая, мг/мл |
Дисперсия |
Среднее квадратическое отклонение |
Средняя арифметическая ошибка |
Р |
tp |
Точность прямых измерений |
Доверительный интервал, мг/мл |
Относительная ошибка (погрешность), % |
|
Телята до приема молозива |
0,5 |
0,0007 |
0,026 |
0,015 |
0,05 |
3,182 |
0,047 |
0,45-0,54 |
9,4 |
|
Телята суточного возраста |
15,35 |
0,0036 |
0,06 |
0,35 |
0,05 |
3,182 |
1,11 |
14,24-16,46 |
7,23 |
|
Телята 2-3-дневного возраста |
15,84 |
0,0017 |
0,04 |
0,024 |
0,05 |
3,182 |
0,76 |
15,08-16,6 |
4,79 |
|
Телята 9-10-дневного возраста |
16,81 |
0,0022 |
0,047 |
0,27 |
0,05 |
3,182 |
0,86 |
15,95-17,67 |
5,1 |
Данные результатов определения иммуноглобулинов отдельных классов характеризуют становление гуморального иммунитета у здоровых телят.
Сравнение полученных данных о содержании иммуноглобулинов и их классов в сыворотке крови новорожденных телят с имеющимся в литературе [М. И. Немченко, Т. Д. Гришина, 1983; В. М. Чекишев, В. М. Васильев, А. И. Кабанцев, 1983] свидетельствует о том, что оно минимально, то есть имеет место гипогаммаглобулинемия.
2.2.7. Уровень антител против О-антигенов Е. coli в нормальном коровьем молозиве, молоке и сыворотке крови новорожденных телят. Телята не получают материнские антитела внутриматочно, а приобретают их из молозива, абсорбируя глобулины, через верхнюю часть тонкого кишечника [R. Aschaffenburg, S. Bartlett, S. K. Kon, D. M. Walker, C. Briggs, E. Cotchin and R. Lovell, 1949b; R. Aschaffenburg, S. Bartlett, S. K. Kon, J. M. Roy and D. M. Walker, 1951a; G. H. Stott A. Pellah, 1982; С. С. Gay, Т. McGuire, S. M. Parish, 1983].
Таблица 2.12
Динамика накопления иммуноглобулина М в сыворотке крови телят (n=16)
|
Определяемая величина |
Средняя арифметическая, мг/мл |
Дисперсия |
Среднее квадратическое отклонение |
Средняя арифметическая ошибка |
Р |
tp |
Точность прямых измерений |
Доверительный интервал, мг/мл |
Относительная ошибка (погрешность), % |
|
Телята до приема молозива |
0,495 |
0,0017 |
0,04 |
0,024 |
0,05 |
3,182 |
0,076 |
0,42-0,57 |
15,3 |
|
Телята суточного возраста |
1,115 |
0,007 |
0,027 |
0,015 |
0,05 |
3,182 |
0,048 |
1,07-1,16 |
4,3 |
|
Телята 2-3-дневного возраста |
1,8225 |
0,01 |
0,1 |
0,017 |
0,05 |
3,182 |
0,54 |
1,28-2,36 |
2,96 |
|
Телята 9-10-дневного возраста |
2,0275 |
0,017 |
0,13 |
0,19 |
0,05 |
3,182 |
0,60 |
1,43-2,63 |
2,96 |
Значение Е. coli в патогенезе диареи новорожденных телят и исключительная важность кормления их молозивом, увеличивающем резистентность к колиинфекции указывали Th. Smith and R. B. Little, 1922; Th. Smith and R.B. Little, 1922b еще в 20-х годах нашего столетия. Первоначальным фактором, связанным с восприимчивостью телят к Е. coli инфекции, является отсутствие в их сыворотке иммуноглобулинов [С. С. Gay, S. M. Parish, T. C. McGuire 1982].
Таблица 2.13
Динамика накопления иммуноглобулина А в сыворотке крови телят (n=16)
|
Определяемая величина |
Средняя арифметическая, мг/мл |
Дисперсия |
Среднее квадратическое отклонение |
Средняя арифметическая ошибка |
Р |
tp |
Точность прямых измерений |
Доверительный интервал, мг/мл |
Относительная ошибка (погрешность), % |
|
Телята до приема молозива |
0,0375 |
0,01 |
0,1 |
0,002 |
0,05 |
3,182 |
0,006 |
0,032-0,044 |
16,0 |
|
Телята суточного возраста |
0,5275 |
0,06 |
0,24 |
0,015 |
0,05 |
3,182 |
0,048 |
0,48-0,57 |
9,09 |
|
Телята 2-3-дневного возраста |
0,705 |
0,03 |
0,19 |
0,011 |
0,05 |
3,182 |
0,035 |
0,67-0,74 |
4,96 |
|
Телята 9-10-дневного возраста |
0,77 |
0,06 |
0,24 |
0,014 |
0,05 |
3,182 |
0,045 |
0,72-0,81 |
5,84 |
У некоторых телят, которые получали молозиво, этот дефицит имеет место подчеркивал C. C. Gay, 1984. По-видимому, некоторые из этих телят утрачивали способность к абсорбции иммуноглобулинов из молозива в течение 4-6 ч после рождения. Факторы, контролирующие абсорбцию гамма глобулинов у новорожденных телят и потеря гаммаглобулинов из их сыворотки, не известны. Потеря гаммаглобулинов из сыворотки телят случается при экскреции в мочу и фецес при нормальном катаболизме. Количество гаммаглобулинов, экскретирующихся в фецес, заметно увеличивается у телят при диареи.
Телята меньше 2-недельного возраста не продуцируют антител после энтеробактериальной О-антигенной (Е. coli) инокуляции. Способность реагировать на антигенную инокуляцию зависит от возраста телят и естественно от иммуногена [С. С. Gay, 1971].
Глобулиновая фракция молозива, ответственная за защиту телят не была идентифицирована, несмотря на то, что различные антитела, имеющиеся в молозиве, трансформируются в сыворотку неонатальных телят [R. Aschaffenburg, S. Bartlett, S. K. Kon, P. Terry, S. Y. Thompson and D. M. Walker, 1949a; R. Aschaffenburg, S. Bartlett, S. K. Kon, D. M. Walker, C. Briggs, E. Cotchin and R. Lovell, 1949b].
Антигены приготовили из трех серотипов Е. coli. Серотипы О78:К80; О119:К69; О137:К79 были изолированы от телят с инфекционной диареей в учхозе «Кетросу» района Анений Ной. Бактерии выращивали на МПА 8-10 ч при 37° С в бактериальных матрацах. Клетки смывали из агара стерильным физиологическим раствором хлорида натрия и бактериальную суспензию автоклавировали при 120° С в течение 2 ч для разрушения К-антигена. Густую отмытую суспензию смешивали с ацетоном в отношении 1:5 до появления коагуляции (свертывания). Плотную часть материала отбирали на воронке Бюхнера промытой один раз ацетоном и затем высушенной эфиром. Препарат был высушен на воздухе и сохраняли при 4° С.
Сухой порошок применяли для приготовления суспензии антигена, добавляя 1 мг высушенных ацетоном бактерий к 1 мл барбиталового буферам [E. Neter, 1957].
Молозиво получали от 23 коров при первом доении после отела.
Двадцать три пробы цельного молока получали от коров средней лактации, сравнивая уровень антител с уровнем в молозиве из того же стада (коровника). Молочную сыворотку приготовляли из молозива и молока и тестировали на антитела, тем же методом что и сыворотку крови. Пробы крови отбирали у телят вскоре после рождения с интервалом 12-24 ч. Кровь выдерживали при комнатной температуре до появления сгустка и сыворотку отделяли в течение 6 ч. Пробы сыворотки крови, молозива и молока хранили при –20° С до тестирования.
2.2.7.1. Серологический тест. Сыворотку инактивировали при 56° С в течение 30 мин для разрушения комплементной активности. Все сыворотки, в том числе молозивные и молочные, абсорбировали с эритроцитами барана, исключая какие либо антитела к эритроцитам. Агглютинирующие антитела определяли по методу E. Neter, E. A. Gorzynski, R. M. Gino, O. Westphal and O. Luderwitz, 1956, за исключением того, что эритроциты барана были утилизированы и был приготовлен антиген, высушенный ацетоном.
Антиглобулиновый тест применяли для определения титра не агглютинирующих антител [E. Neter, 1957] используя антиглобулиновую (бычью) сыворотку. Все титрации проводили в двукратном разведении, ранг серии от 2-2 до 2-11, применяя 0,1 мл разведенной сыворотки и 0,1 мл 2 % суспензии антигена с эритроцитами барана, которые были промыты 3 раза после антигенной абсорбции.
Агглютинационный тест учитывали после 10-120 минутной выдержки при комнатной температуре.
Антиглобулиновый тест определяли после инкубации в термостате при 37° С в течение 1 ч и выдержки в холодильнике при 4° С в течение 12 ч.
Высшее сывороточное разведение, которое имело хорошо выраженную агглютинацию, учитывали как титр этой сыворотки. Чувствительность антител к 2-меркаптоэтанолу определяли инкубированием сыворотки на 0,1М растворе CH>2>SH CH>2>OH, при 37° С в течение 30 мин. Резистентность к этой обработке была взята как показание, что антитела не являются макроглобулинами.
2.2.7.2 Абсорбционный тест. Определение специфичности антител абсорбционным методом было проведено с сывороткой молозива и крови телят. Сывороточные образцы получали от телят через 12-24 ч после кормления молозивом, и были разделены по уровню антител против трех антигенов. Сыворотку молозива и сыворотку крови телят абсорбировали с каждым из трех серотипов Е. coli , которые инкубировали в колбе-матраце на МПА.
Таблица 2.14
Уровень колостральных антител против О-антигенов Е. coli в сыворотке молозива коров
|
Антиген |
Тест |
Количество проб |
Образцы с положительным результатом, % |
Средний титр |
|
О78:К80 |
Агглютинационный |
15 |
93 |
7,86±0,93 |
|
Антиглобулиновый |
15 |
100 |
9,07±0,98 |
|
|
О119:К69 |
Агглютинационный |
14 |
100 |
8,43±0,99 |
|
Антиглобулиновый |
13 |
100 |
9,46±1,21 |
|
|
О137:К79 |
Агглютинационный |
16 |
100 |
9,62±0,71 |
|
Антиглобулиновый |
17 |
100 |
10,23±0,99 |
Микроорганизмы смывали из агара барбиталовым (вероналовым) буфером и центрифугировали при 12000 об/мин в течение 30 мин. Затем бактерии прибавляли к сыворотке крови телят и молозивной сыворотке — 2×1011 бактерий на 1 мл пробы. Абсорбцию проводили при 37° С в течение 60 мин и суспензию центрифугировали при 12000 об/мин в течение 30 мин. Супернатант сыворотки крови телят и молозивной сыворотки удаляли из массы клеток и выдерживали при 20° С до тестирования.
Пробы имели достоверный титр антител против всех трех антигенов за исключением 2-х проб, которые не имели агглютинирующих антител против О78:К80, но имели антитела, обнаруживаемые антиглобулиновым тестом в титрах 7-8. В двух других образцах сыворотки молозива отсутствовали антитела, выявляемые антиглобулиновым тестом против О-антигена О137:К79, но имели агглютинирующие антитела в титрах от 2 до 5. Высшую среднюю активность агглютинирующих и антиглобулиновых антител отмечали против О137:К79.
Затем в порядке уменьшения активности антител к О-антигену были О119:К69 и О78:К80.
Титры агглютинирующих и антиглобулиновых антител в сыворотке молока были значительно меньшими, чем в сыворотке молозива и, в дополнение, сфера действия антител была более ограниченной. Учитывали только средний титр антител реагирующих против специфических О-антигенов Е. coli: О78:К80, О119:К69, О137:К79 (табл. 2.14; 2.15).
Таблица 2.15
Уровень антител против О-антигенов Е. coli в сыворотке молока коров
|
Антиген |
Тест |
Количество проб |
Образцы с положительным результатом, % |
Средний титр |
|
О78:К80 |
Агглютинационный |
15 |
(3) 20 |
3,0±0 |
|
Антиглобулиновый |
15 |
(12) 80 |
3,75±0,94 |
|
|
О119:К69 |
Агглютинационный |
14 |
(3) 21,4 |
2,67±0,57 |
|
Антиглобулиновый |
13 |
(7) 50 |
3,86±0,90 |
|
|
О137:К79 |
Агглютинационный |
16 |
(4) 25 |
3,25±0,50 |
|
Антиглобулиновый |
17 |
(13) 76,47 |
4,31±1,08 |
2.2.7.3. Уровень антител против О-антигенов Е. coli в сыворотке крови новорожденных телят. Активность антител не была обнаружена в сыворотке неонатальных телят перед кормлением молозивом. После выпаивания молозива сывороточные образцы от 23 телят взаимодействовали с одним или тремя тестированными О-антигенами. Количество сывороточных проб и процент положительных тест реакций представлен в табл. 2.16, из которой следует, что некоторые сыворотки реагировали только в антиглобулиновом тесте, в агглютинационном тесте реакция была отрицательной. Все сыворотки тест позитивные в агглютинационном опыте, тестировались и в антиглобулиновой реакции. Обработка исследованных сывороток этантиолом показала примерно те титры антител, которые были определены агглютинационным и антиглоблииновым методами.
Таблица 2.16
Количество образцов с положительным тест результатом при исследовании уровня антител против О-антигенов Е. coli в сыворотке крови неонатальных телят.
|
Антиген |
Количество образцов |
Положительный тест результат, % |
|
|
Агглютинационный тест |
Антиглобулиновый тест |
||
|
О78:К80 |
15 |
(8) 53,3 |
(10) 66,7 |
|
О119:К69 |
15 |
(8) 53,3 |
(11) 73,3 |
|
О137:К79 |
15 |
(10) 66,7 |
(12) 80,0 |
Результаты титрации парных образцов сыворотки молозива, полученного при первом доении коров и сыворотки, приготовленной из крови телят через 12-24 ч после кормления молозивом представлены в табл. 2.17.
В таблице учтены только тест позитивные реакции против специфических О-антигенов Е. coli : О78:К80, О119:К69, О137:К79. В некоторых пробах титр антител в сыворотке крови телят был незначительно выше или равнялся уровню антител в сыворотке молозива. Тем не менее, средний титр в сыворотке крови телят был почти на два разведения меньше, чем в сыворотке молозива.
Таблица 2.17
Уровень антител против О-антигенов Е. coli в спаренных сыворотках молозива и крови неонатальных телят.
|
Антиген |
Тест |
Количество проб |
Средний титр в сыворотках |
|
|
молозива коров |
крови телят |
|||
|
О78:К80 |
Агглютинационный |
8 |
7,87±0,84 |
5,37±0,87 |
|
Антиглобулиновый |
10 |
9,1±0,99 |
5,9±0,73 |
|
|
О119:К69 |
Агглютинационный |
8 |
8,37±0,92 |
6,12±0,83 |
|
Антиглобулиновый |
11 |
9,45±0,82 |
7,27±0,84 |
|
|
О137:К79 |
Агглютинационный |
10 |
9,60±0,84 |
7,3±0,77 |
|
Антиглобулиновый |
12 |
10,25±0,96 |
9,42±0,82 |
2.2.7.4 Абсорбция антител против О-антигенов Е. coli из сыворотки молозива и крови телят. Две пробы сыворотки молозива и 3 сыворотки крови однодневных телят были абсорбированы Е. coli антигеном для определения специфичности антител. Данные, полученные абсорбционным тестом, представлены в табл. 2.18.
Все исследуемые пробы, абсорбированные антигеном, подавляли активность антител к антигену, в то же время, как правило, превосходили антитела против других антигенов. Антитела против антигена О137:К79 имели влияние на абсорбцию по сравнению с другими антигенами.
Почти все пробы сывороток имели антитела в высоких титрах против О-антигенов к трем серотипам Е. coli . Сходные антитела были выявлены в нормальном молоке в меньшей степени и в значительно низких титрах. Колостральные антитела вероятно, отражали контакт (инфицирование) коров с различными серотипами Е. coli . Титр антител против О78:К80 был наименьшим из трех тестированных антигенов. Результаты абсорбционного теста указывают, что антитела против О78:К80 наиболее легко абсорбируются по сравнению с другими серотипами Е. coli . Эти данные указывают, что антитела реагирующие с О78:К80 не обладают высокой специфичностью и могут вступать в перекрестную реакцию с другими микроорганизмами, инфицирующими коров. Высокий уровень антител, выявляемый в сыворотке молозива, соответствовал и высокому уровню антител в сыворотке крови телят через несколько часов после кормления молозивом.
Таблица 2.18
Показатели абсорбции антител против О-антигенов Е. coli молозивом и сывороткой неонатальных телят.
|
Источник антител |
Абсорбент |
Титр антител |
|||||
|
О78:К80 |
О119:К69 |
О137:К79 |
|||||
|
АГТ* |
АНТ |
АГТ |
АНТ |
АГТ |
АНТ |
||
|
Молозиво №4456 |
нет |
(1:32) 5 |
(1:128) 7 |
(1:128) 7 |
(1:256) 8 |
(1:64) 6 |
(1:512) 9 |
|
О78:К80 |
0 |
0 |
(1:32) 5 |
(1:128) 7 |
(1:64) 6 |
(1:512) 9 |
|
|
О119:К69 |
0 |
0 |
0 |
0 |
(1:16) 4 |
(1:128) 7 |
|
|
О137:К79 |
0 |
0 |
(1:32) 5 |
(1:128) 7 |
0 |
0 |
|
|
Молозиво №3457 |
нет |
(1:32) 5 |
(1:64) 6 |
(1:32) 15 |
(1:256) 18 |
(1:64) 6 |
(1:1024) 10 |
|
О78:К80 |
0 |
0 |
(1:16) 4 |
(1:64) 6 |
(1:64) 6 |
(1:1024) 10 |
|
|
О119:К69 |
0 |
(1:32) 5 |
0 |
0 |
(1:64) 6 |
(1:1024) 10 |
|
|
О137:К79 |
0 |
(1:8) 3 |
(1:32) 5 |
(1:128) 7 |
0 |
0 |
|
|
Сыворотка телят №4757 (однодневный возраст) |
нет |
(1:64) 6 |
(1:128) 7 |
(1:128) 7 |
(1:256) 8 |
(1:32) 5 |
(1:512) 9 |
|
О78:К80 |
0 |
0 |
(1:16) 4 |
(1:128) 7 |
(1:32) 5 |
(1:512) 9 |
|
|
О119:К69 |
(1:32) 5 |
(1:128) 7 |
0 |
0 |
(1:32) 5 |
(1:128) 7 |
|
|
О137:К79 |
(1:16) 4 |
(1:64) 6 |
(1:128) 6 |
(1:128) 7 |
0 |
0 |
|
|
Сыворотка телят №4759 (однодневный возраст) |
нет |
(1:128) 7 |
(1:256) 8 |
(1:256) 8 |
(1:512) 9 |
(1:256) 8 |
(1:512) 9 |
|
О78:К80 |
0 |
0 |
(1:32) 5 |
(1:64) 6 |
(1:128) 7 |
(1:256) 8 |
|
|
О119:К69 |
(1:4) 2 |
(1:8) 3 |
0 |
0 |
(1:64) 6 |
(1:256) 8 |
|
|
О137:К79 |
(1:32) 5 |
(1:64) 6 |
(1:64) 6 |
(1:128) 7 |
0 |
0 |
|
|
Сыворотка телят №4767 (однодневный возраст) |
нет |
(1:64) 6 |
(1:64) 6 |
(1:128) 7 |
(1:512) 9 |
(1:64) 6 |
(1:1024) 10 |
|
О78:К80 |
0 |
0 |
(1:16) 4 |
(1:256) 8 |
(1:16) 4 |
(1:32) 5 |
|
|
О119:К69 |
(1:8) 3 |
(1:16) 4 |
0 |
0 |
(1:16) 4 |
(1:128) 7 |
|
|
О137:К79 |
(1:16) 4 |
(1:32) 5 |
(1:32) 5 |
(1:512) 9 |
0 |
0 |
* Титр выражен в log>2>> >АГТ — агглютинационный тест АНТ — антиглобулиновый тест
Антитела в сыворотках молозива и крови однодневных телят были резистентными к инактивации CH>2>SH-CH>2>OH-меркаптоэтанолом, что указывает на то, что они не являются макроглобулинами. Этот результат подтверждает данные, что антитела не продуцируются новорожденными телятами, а приобретаются после рождения из молозива. У семи из 23 телят, кормившихся молозивом, активность агглютинирующих антител против трех О-антигенов Е. coli не была установлена. Эти же телята имели и низкий уровень иммуноглобулинов после кормления молозивом, что указывает на анормальную абсорбцию иммуноглобулинов из желудочно-кишечного тракта.
2.2.8. Уровень антител против К-антигенов Е. coli в нормальном коровьем молозиве, молоке и в сыворотке крови новорожденных телят. Большое значение молозива в защите неонатальных телят от колибактериоза было установлено давно [Th. Smith and R. B. Little, 1922]. Полагают, что молозиво обеспечивает защиту в ассоциации с принадлежащей ему активностью антител против Е. coli и, особенно, с содержащими агглютининами к К-антигену [B. Kaijser, S. Ahlstedt, 1977].
Значение К агглютининов, как фактора в молозиве, который защищает телят против колибактериоза установили [С. J. Howard, A. A. Glynn, 1971], после того как некоторые телята приобретали после рождения агглютинины из молозива, которые реагировали с К-антигеном из штаммов Е. coli ассоциирующими с заболеванием.
Агглютинины к К-антигену из Е. coli ассоциировались с увеличением вирулентности микроорганизмов [С. С. Gay, S. M. Parish, T. C. McGuire 1982]. Следовательно, антитела против К-антигена считают доказанным фактором в резистентности неонатальных телят против Е. coli инфекции.
Вначале исследования по определению антител в К-антигену, проводили прямой агглютинацией к идентифицированным микроорганизмами [E. Neter, E. A. Gorzynski, R. M. Gino, O. Westphal and O. Luderwitz. 1956]. Более чувствительные методы выявления антител в человеческой крови из пуповины к энтеробактериальным антигенам были установлены при применении пассивной гемагглютинации [E. Neter, 1957]. Антитела, выявляемые этими чувствительными гемагглютинационными тестами, могут быть существенными в увеличении резистентности неонатальных телят против колибактериоза. Эти антитела, пассивно приобретенные после рождения телятами из молозива коров, были тестированы в динамике.
В опыте находились 23 телят черно-пестрой породы в первые 5 дней. Телят кормили молозивом, а затем свежим молоком.
Антигены приготовляли из трех серотипов Е. coli . Серотипы О78:К80, О119:К69 и О137:К79 были изолированы от телят с инфекционной диареей в учхозе «Кетросу» района Анений Ной. Бактерии выращивали на кровяном агаре, при этом гладкие и слизеподобные колонии пересевали на агар Мак-Конки.
Часть колоний из агара Мак Конки были тестированы пластинчатой реакцией агглютинации (на предметном стекле со специфическими К-антисыворотками). Колонии-тест-положительные были отобраны для приготовления К-антигена Е. coli.
Культивирование бактерий и приготовление антигенов было сходным с получением О-антигенов Е. coli за исключением того, что Е. coli применявшаяся для приготовления К-антигена не нагревалась, и для смыва бактерий использовали солевой раствор (0,85 %), содержащий 0,5 % формальдегида. Бактерии были экстрагированы при добавлении ацетона непосредственно после их смыва с агаровой культуры и промыты.
Пробы молозива отбирали от 23 коров в молочном стаде учебного хозяйства при первом доении после отела. Двадцать три пробы цельного молока получали от коров средней лактации, сравнивая уровень антител с уровнем в молозиве. Молочную сыворотку приготовляли из молозива и молока и тестировали на антитела, подобным методом, как и сыворотку крови. Пробы крови отбирали у телят вскоре после рождения с интервалом 12-24 ч. Кровь выдерживали при комнатной температуре до появления сгустка и сыворотку отделяли в течение 6 ч. Пробы сыворотки крови, молозива и молока хранили при -20° С.
2.2.8.1. Серологический тест. Для разрушения комплементной активности сыворотку инактивировали при 56° С в течение 30 мин. Все сыворотки в том числе молозивные и молочные, абсорбировали с эритроцитами барана, исключая какие-либо антитела к эритроцитам. Агглютинирующие антитела определяли по методу Е. Neter (1957), за исключением того, что эритроциты барана были утилизированы и был приготовлен антиген высушенный ацетоном.
Антиглобулиновый тест применяли для определения неагглютинирующих антител [E. Neter, 1957], применяя антиглобулиновую (бычью) сыворотку. Все титрации проводили в двукратном разведении ранг серии от 2-2 до 2-11, применяя 0,1 мл разведенной сыворотки и 0,1 мл 2 % суспензии антигена с эритроцитами барана, которые были промыты 3 раза после антигенной абсорбции.
Агглютинационный тест учитывали после 90-120 минутной инкубации при комнатной температуре.
Антиглобулиновый тест определяли после 1 ч инкубации в термостате при 37° С и 12 ч экспозиции в холодильнике при 4° С.
Предельное сывороточное разведение, которое имело хорошо выраженную агглютинацию, учитывали как титр этой сыворотки. Чувствительность антител к 2-меркаптоэтанолу определяли инкубацией сыворотки в 0,1М растворе CH>2>SH-CH>2>OH при 37° С в течение 30 мин. Резистентность к этой обработке была взята как показание, что антитела не являются макроглобулинами.
2.2.8.2. Абсорбционный тест. Определение специфичности антител абсорбционным методом было проведено с сывороткой молозива и крови телят. Сывороточные образцы получали от телят через 12-24 ч после кормления молозивом и были разделены по уровню антител против трех антигенов. Сыворотку молозива и сыворотку крови телят абсорбировали с каждым из трех серотипов у Е. coli . Три серотипа Е. coli инкубировали в колбе-матраце на МПА.
Микроорганизмы смывали из агара барбиталовым (вероналовым) буфером и центрифугировали при 12000 об/мин в течение 30 мин.
Затем бактерии прибавляли к сыворотке крови телят и молозивной сыворотке из расчета 2×1011 бактерий на 1 мл. Абсорбцию проводили при 37° С в течение 60 мин и суспензию центрифугировали при 12000 об/мин 30 мин. Супернатант сыворотки телят и молозивной сыворотки удаляли из массы клеток, а осадок хранили при -20° С до тестирования.
2.2.8.3. Уровень антител против К-антигенов в сыворотках молозива и молока. Средний титр антител в молозивной сыворотке против К антигенов из 3 штаммов Е. coli представлен в табл. 2.19. Агглютинирующиеся антитела выявляются менее часто, чем антиглобулиновые антитела в тестированных образцах молозива. Положительный тест — результат установлен соответственно в 53,83 % (антиген О78:К80) и 88,24 % (антиген О137:К79) случаев. Средний титр антител в аналогичных условиях равнялся 4,63±0,73 и 6,87±0,56.
Таблица 2.19.
Уровень колостральных антител против К-антигенов Е. coli в сыворотке молозива коров
|
Антиген |
Тест |
Количество проб |
Образцы с положительным результатом, % |
Средний титр |
|
О78:К80 |
Агглютинационный |
15 |
53,33 |
4,63±0,73 |
|
Антиглобулиновый |
15 |
80,00 |
5,92±0,67 |
|
|
О119:К69 |
Агглютинационный |
14 |
71,43 |
5,6±0,70 |
|
Антиглобулиновый |
13 |
92,31 |
6,42±0,67 |
|
|
О137:К79 |
Агглютинационный |
16 |
75,00 |
6,25±0,69 |
|
Антиглобулиновый |
17 |
88,24 |
6,87±0,56 |
Из 69 исследованных проб сывороток молозива агглютининовым и антиглобулиновым тестами 9 проб имели антитела, обнаруживаемые только антиглобулиновым методом. Средний титр антител был существенно выше при тестировании антиглобулиновым методом по сравнению с агглютинационным, тем не менее, в 4 образцах получены совпадающие результаты, а в одном случае титр агглютининов был выше на одно разведение. Средний титр был вычислен только из тест положительных образцов. Высокий титр антител был установлен агглютинационным и антиглобулиновым методами против К антигена серотипа О137:К79 (6,25±0,69 и 6,87±0,56).
Результаты исследования образцов молока, полученного от коров на различных стадиях лактации, показаны в табл. 2.20. Антитела против Е. coli К-антигена были обнаружены в молоке агглютинационным и антиглобулиновым методами, но частота и уровень активности антител были намного меньшими, чем в молозиве.
Таблица 2.20
Уровень антител против К-антигенов Е. coli в сыворотке молока коров
|
Антиген |
Тест |
Количество проб |
Образцы с положительным результатом, % |
Средний титр |
|
О78:К80 |
Агглютинационный |
15 |
21,43 |
3,33±0,58 |
|
Антиглобулиновый |
15 |
42,87 |
3,17±0,41 |
|
|
О119:К69 |
Агглютинационный |
14 |
(3) 23,08 |
2,67±0,58 |
|
Антиглобулиновый |
13 |
(8) 66,67 |
3,63±0,75 |
|
|
О137:К79 |
Агглютинационный |
16 |
(6) 46,15 |
3,17±0,41 |
|
Антиглобулиновый |
17 |
(11) 84,62 |
4,36±0,92 |
Двенадцать из 37 проб (тест положительных) сывороток молока имели антитела против 3 тестированных антигенов. Причем 12 проб были тест положительными при исследовании как агглютинационным, так и антиглобулиновым тестами, 27 тест положительных проб выявляли только антиглобулиновым методом. Из 14 проб сывороток молока выявляли антитела против К антигена Е. coli штамм О78:К80 в трех совпадающих пробах агглютинационным и антиглобулиновым методом. Антитела к К антигену Е. coli штамм О119:К69 выявлены тестированными методами из 13 проб в 3 случаях и дополнительно в 5 случаях только антиглобулиновым методом. Антитела к К антигену Е. coli штамм О137:К79 выявляли в 6 совпадающих случаях обеими методами и дополнительно в 5 пробах только антиглобулиновым методом.
2.2.8.4. Уровень антител против К антигена в сыворотке новорожденных телят. Результаты исследования сывороточных проб телят в возрасте от 1 до 5 дней указывает, что большинство телят приобретают после рождения антитела, выявляемые антиглобулиновым, а не агглютинационным тестом. Сопоставление уровня антител в сыворотке молозива и в сыворотке крови телят через 12-24 ч после кормления молозивом, представлены в табл. 2.21.
Таблица 2.21
Уровень антител против К-антигенов Е. coli в спаренных сыворотках молозива и крови неонатальных телят.
|
Антиген |
Тест |
Количество проб |
Средний титр в сыворотках |
|
|
молозива коров |
крови телят |
|||
|
О78:К80 |
Агглютинационный |
5 |
4,40±0,54 |
3,20±0,45 |
|
Антиглобулиновый |
5 |
6,80±0,87 |
5,80±0,45 |
|
|
О119:К69 |
Агглютинационный |
7 |
5,43±0,53 |
4,14±0,37 |
|
Антиглобулиновый |
6 |
7,17±0,41 |
6,17±0,41 |
|
|
О137:К79 |
Агглютинационный |
6 |
5,83±0,41 |
4,83±0,41 |
|
Антиглобулиновый |
7 |
7,43±0,53 |
6,71±0,48 |
В нескольких случаях титр антител в сыворотке крови телят был эквивалентным уровню колостральных антител, но, как правило, титр антител в сыворотке телят был меньшим на 1-2 разведения, чем титр в сыворотке молозива. Телята, лишенные молозива, не содержали антител против К антигена Е. coli тестированных серотипов. Антитела в молозиве и антитела в сыворотке неонатальных телят не инактивировались при обработке 2-меркаптоэтанолом CH>2>SH CH>2>OH. Средний титр антител в сыворотке крови телят, кормившихся молозивом варьировал в зависимости от тестированных К антигенов Е. coli О78:К80, О119:К69, О137:К79. Существенное различие в уровне антител установлено при их выявлении агглютинационным и антиглобулиновым методами(табл. 2.22).
Уровень антител при исследовании антиглобулиновым методом был на 2-3 разведения большим по сравнению с агглютинационным тестом.
Таблица 2.22
Количество образцов с положительным тест результатом при исследовании уровня антител против К-антигенов Е. coli в сыворотке крови неонатальных телят.
|
Антиген |
Количество образцов |
Положительный тест результат, % |
|
|
Агглютинационный тест |
Антиглобулиновый тест |
||
|
О78:К80 |
16 |
31,25 |
33,33 |
|
О119:К69 |
16 |
33,33 |
56,26 |
|
О137:К79 |
16 |
31,25 |
62,5 |
2.2.8.5. Абсорбция антител против К-антигенов Е. coli из сыворотки молозива и крови телят. Результаты абсорбционного теста колостральных образцов, полученных от коров при первом доении после отела и сывороточных образцов, взятых у телят в течение 1 дня после кормления молозивом представлены в табл. 2.23.
Все исследуемые пробы, абсорбированные антигенами, подавляли активность антител к антигенам Е. coli О78:К80, О115:К69. Антитела против антигена О137:К79 имели наименьшее влияние на абсорбцию по сравнению с другими антигенами. Два серотипа Е. coli О78:К80, О119:К69 имели К антиген В типа, тогда как серотип О137:К79 имел К антиген А типа. Это различие в типах К антигена и имеет значение в перекрестной реакция с серотипом О137:К79.
Таблица 2.23
Показатели абсорбции антител против К-антигенов Е. coli молозивом и сывороткой неонатальных телят.
|
Источник антител |
Абсорбент |
Титр антител |
|||||
|
О78:К80 |
О119:К69 |
О137:К79 |
|||||
|
АГТ* |
АНТ |
АГТ |
АНТ |
АГТ |
АНТ |
||
|
Молозиво №4456 |
нет |
0 |
(1:32) 5 |
0 |
(1:64) 6 |
(1:32) 5 |
(1:64) 6 |
|
О78:К80 |
0 |
0 |
0 |
0 |
0 |
0 |
|
|
О119:К69 |
0 |
0 |
0 |
0 |
0 |
0 |
|
|
О137:К79 |
0 |
(1:16) 4 |
0 |
0 |
0 |
0 |
|
|
Молозиво №3457 |
нет |
0 |
(1:16) 4 |
0 |
(1:32) 5 |
(1:64) 6 |
(1:16) 4 |
|
О78:К80 |
0 |
0 |
0 |
0 |
0 |
0 |
|
|
О119:К69 |
0 |
0 |
0 |
0 |
0 |
0 |
|
|
О137:К79 |
0 |
0 |
0 |
0 |
0 |
0 |
|
|
Сыворотка телят №4757 (однодневный возраст) |
нет |
(1:8) 3 |
(1:128) 7 |
(1:64) 6 |
(1:128) 7 |
(1:128) 7 |
(1:1024) 10 |
|
О78:К80 |
0 |
0 |
0 |
0 |
(1:64) 6 |
(1:128) 7 |
|
|
О119:К69 |
0 |
0 |
0 |
0 |
(1:64) 6 |
(1:512) 9 |
|
|
О137:К79 |
0 |
0 |
0 |
0 |
0 |
0 |
|
|
Сыворотка телят №4759 (однодневный возраст) |
нет |
(1:32) 5 |
(1:128) 7 |
(1:32) 5 |
(1:256) 8 |
(1:64) 6 |
(1:512) 9 |
|
О78:К80 |
0 |
0 |
0 |
0 |
0 |
0 |
|
|
О119:К69 |
0 |
0 |
0 |
0 |
0(1:64) 6 |
0 |
|
|
О137:К79 |
0 |
0 |
0 |
0 |
0 |
0 |
|
|
Сыворотка телят №4767 (однодневный возраст) |
нет |
0 |
(1:64) 6 |
(1:16) 4 |
(1:64) 6 |
(1:32) 5 |
(1:128) 7 |
|
О78:К80 |
0 |
0 |
0 |
0 |
0 |
(1:16) 4 |
|
|
О119:К69 |
0 |
0 |
0 |
0 |
(1:16) 4 |
(1:64) 6 |
|
|
О137:К79 |
0 |
0 |
0 |
0 |
0 |
0 |
* Титр, выраженный в log>2>
АГТ — агглютинационный тест
АНТ — антиглобулиновый тест
Наличие и титр антител против К антигенов Е. coli О78:К80, О119:К69 и О137:К79 в молозиве был ниже, чем антитела против О антигенов в тех же самых пробах. Процент тест положительных молозивных образцов с К-антигенами составлял при исследовании агглютинационным методом от 53,33 да 75,00, а при исследовании антиглобулиновым методом от 53,33 до 88,24, тогда как асе образцы были тестированы положительно с Е. coli О антигенами. Анти-К активность антител в молоке имела частоту и титр ниже, чем активность в молозиве. Пробы молока не имели антител против всех трех К антигенов.
Иммуноглобулины в молозиво извлекаются селективной секрецией из сыворотки крови коров. Следовательно, специфичность антител в молозиве определяется специфичностью антител в сыворотке крови коров. Кроме того, уровень антител в сыворотке крови коров зависит от воздействия на иммунокомпетентные клетки специфических иммуногенов, то есть активность антител молозива коров является рефлекторным отражением специфичности антигена. Вероятно, если определенные серотипы Е. coli имеются во внешней среде (коровник, родильное отделение), коровы впоследствии продуцируют специфические антитела и передают их через молозиво телятам. Это имеет место в том случае, когда инвазивные вирулентные серотипы Е. coli вызывают антигенную стимуляцию иммунокомпетентных клеток большую, чем не вирулентные штаммы.
Высший средний титр колостральных антител анти-К антигена был установлен против штамма О137:К79. Идентификация антител в сыворотке молозива и в сыворотке крови телят против серотипов О78:К80 и О119:К69 в более низких титрах и низшей частоте может постулироваться их незначительной вирулентностью и инвазивностью или тем, что антитела обладают перекрестной реакцией и не специфичны для этих серотипов.
Результаты абсорбционного теста сыворотки молозива и сыворотки крови телят указывают на перекрестную абсорбцию антител против Е. coli К антигенов исследуемых серотипов. Перекрестная абсорбция была, особенно, очевидной с 2-мя типами В антигенов, так как абсорбция с 1 антигеном идентифицирует антитела против 2 антигенов (О78:К80 и О119:К69). Эти данные подтверждают значительную перекрестную реактивность антител к К-антигенам. Такая кросс-реактивность антител обеспечивает защиту неонатальных телят против инфекции с более чем 1 штаммом Е. coli . Следовательно, антитела с перекрестной активностью против К антигенов Е. coli в естественных условиях способствуют повышению резистентности неонатальных телят против различных штаммов Е. coli .
Активность антител против К антигенов в молозиве и сыворотке телят 1-2 дневного возраста была более низкой чем активность против О антигенов. Низкий титр К-антител в сыворотке неонатальных телят можно было предположить, так как незначительная частота и низкий титр К-антител был в молозиве коров. В кишечнике новорожденных телят иммуноглобулины абсорбируются не селективно. Антитела против Е. coli К антигенов в молозиве были резистентные против обработки 2-меркаптоэтанолом, как и антитела в сыворотке новорожденных телят. Эти результаты указывают, что антитела не являются макроглобулинами.
Различная концентрация антител (титр) в сыворотке новорожденных телят против К и О антигенов указывает, что кормление молозивом может стимулировать или подавлять синтез антител к определенным антигенам в одинаковой степени независимо от места их локализации в бактериальных клетках.
2.2.9. Протективный механизм молозива при колибактериозе новорожденных телят. Выявление механизма протективного действия молозива при колибактериозе новорожденных телят является определяющим фактором в прогнозировании данного заболевания. В молозиве присутствуют протективные антитела против Е. coli и низкое их содержание в отдельных образцах связано с возрастом коров [Th. Smith and R. B. Little, 1922b]. В последующем [C. Briggs, 1951], изучая антигенную структуру Е. coli , доказал, что реакция между К антигеном и антителами связана с защитой и основана на иммунологическом протективном значении молозива. Сущность протективного фактора против колибактериоза ассоциируется с лактоглобулиновой фракцией [R. Aschaffenburg, S. Bartlett, S. K. Kon, P. Terry, S. Y. Thompson and D. M. Walker, 1949a; R. Aschaffenburg, S. Bartlett, S. K. Kon, J. M. Roy and D. M. Walker, 1951a].
Молозиво от коров получали из хозяйств: «Кетросу» района Анений Ной и ОПХ «Колоница» района Криулень. Молозиво (100 мл) нагревали до 37° С и добавляли 4 мл разведенного (1:10) сычужного фермента. После отделения сыворотки (через 30-60 мин) проводили фильтрацию для освобождения от казеина. Бульонную культуру Е. coli тестировали для определения DL>50> путем внутрибрюшинного заражения белых мышей. Эта доза составила 0,25 мл. Мышей заражали интраперитонально в дозе DL>50> тест- микроорганизмами и через 2 ч после инъекции вводили 0,5 молозивной сыворотки в разведении 1:5-1:20 соответственно титру. Мыши были под наблюдением в течение 72 ч. в порядке дифференциации между «К» и «О» агглютинацией, суспензию готовили для тест агглютинации следующим образом: 1- живая; 2- гретая при 100°С; 3- гретая при 120°С 2 ч. В опытах использовали три серотипа Е. coli: О78:К80, 0119:К69, 0137:К79. Для протективного теста использовали 18-24 ч бульонную культуру Е. соli каждого из вышеназванных серотипов.
Агглютинация («О», «К») и протективный тест на белых мышах представлены в табл. 2.24. «К» антитела, содержащиеся в молозиве и обладающие протективной активностью для белых мышей, отличались, как по титру, так и по эффекту для разных серотипов и проб сывороток молозива. Тест агглютинация против микробной суспензии, приготовленной тремя различными методами, является убедительным доказательством характеризующим высокую специфичность реакции. Данные табл. 2.24 подтверждают, что содержащиеся в молозиве антитела-агглютинины против «К» антигена Е. соli обладают высокой протективной активностью. Молозиво, содержащее антитела-агглютинины против «О» антигена обеспечивает незначительную защиту. Пробы молозивной сыворотки, не содержащие агглютинины против «К» и «О» антигенов, не обладали протективной активностью. Следовательно, протективная эффективность проб сывороток молозива против серотипов Е. соli может быть установлена определением титра «К» антител.
Культура Е. соli была изолирована из крови яремной вены двух больных телят, один из которых погиб через 24 ч, а другой через 36 ч. Телятам скармливали молозиво и определяли массу тела, как у живых, так и у погибших. Из одиннадцати телят, девять получавших 400 мл молозива в первое кормление, погибали в течение двух недель. Исследованием штаммов Е. соli, изолированных от этих телят определили содержание «К» антител-агглютининов в пробах, используемого молозива. Изолированные штаммы Е. соli, выделённые из крови сердца и костного мозга при аутопсии принадлежали к серотипам О78:К80, О119:К69, О137:К79.
Таблица 2.24
Титр колостральных антител и протективные свойства молозивной сыворотки к серотипам Е. coli, изолированных от павших телят.
|
Серотипы |
Тест |
Пробы молозивной сыворотки |
||
|
I |
II |
III |
||
|
О78:К80 |
«К» агглютинация |
- |
1:320 |
- |
|
«О» агглютинация |
1:80 |
- |
- |
|
|
Протективный |
7/10* |
5/10 |
8/10 |
|
|
О119:К69 |
«К» агглютинация |
1:640 |
- |
1:160 |
|
«О» агглютинация |
- |
1:80 |
- |
|
|
Протективный |
5/10 |
8/10 |
6/10 |
|
|
О137:К79 |
«К» агглютинация |
1:1280 |
1:160 |
- |
|
«О» агглютинация |
- |
- |
- |
|
|
Протективный |
1/10 |
7/10 |
9/10 |
* летальность мышей отмечали в числителе, а количество зараженных в знаменателе.
Исследованные пробы молозива различались как по титру, так и по содержанию «К» антител по отношению к исследуемым серотипам Е. соli. Так, например, проба молозива 1 не содержала «К» антител против серотипа О78:К80, проба II – против серотипа О119:К69, проба III – против серотипов О78:280; О137:К79.
Доказательством взаимосвязи между защитой телят молозивом и присутствием в том же самом молозиве антител- агглютининов против «К» антигенов, в частности, к исследуемым серотипам Е. соli подтверждается протективным действием молозива – как результат специфической активности.
Так как, «К» антитела-агглютинины в молозиве являются фактором, защищающим белых мышей против экспериментального колибактериоза, эффективность проб молозива против определенных серотипов Е. coli можно тестировать агглютинационным методом. Следовательно, энзоотия колибактериоза новорожденных телят объясняется дефицитом агглютининов против «К» антигена Е. coli. Молозиво, скармливаемое телятам, погибшим от энзоотического колибактериоза, не содержало антител агглютининов против «К» антигенов Е. coli , изолированных от павших телят.
Таким образом, протективный механизм молозива является в значительной степени иммунологическим и ассоциируется с антителами против «К» антигенов Е. coli , вызывающих колибактериоз неонатальных телят.
2.2.10. Факторы, влияющие на выживание новорожденных телят. Успешное выращивание новорожденных телят зависит от целого ряда факторов, в том числе от своевременного скармливания молозива в достаточном количестве, а также от состояния молочной железы коровы, умения и мастерства обслуживающего персонала. Ущерб на отдельных фермах республики был в диапазоне от 10 до 80 % и часто отражал организацию содержания и кормления [А. В. Драгомир, Е. А. Драгомир, А. Ф. Карышева, Ф. В. Спатарь, 1985].
Однако наиболее важным фактором, влияющим на выращивание новорожденных телят, является абсорбция колостральных антител в соответствующем физиологическом количестве. Молозиво здоровых коров — наиболее сильнодействующее лекарственное средство для телят. Оно обеспечивает большую защиту новорожденных, чем фармацевтика [V.C.R. Irvin, 1974]. Это было доказано в большинстве случаев, когда выживание телят в значительной степени зависело от адекватного поглощения молозива и абсорбции.
Летальность среди телят, лишенных молозива, достигала 70-90 % с наибольшим падежом от эшерихиоза или пастереллезной пневмонии. Несмотря на то, что новорожденные телята иммунокомпетентные при рождении, они не способны быстро анамнестически реагировать, то есть подвергать действию лимфоцитов определенным антигенным повторным воздействиям. Поэтому несколько дней необходимо телятам для установления иммунологической реактивности и продуцирования иммуноглобулинов. В этот лаг-период (промежуток времени между раздражением и ответной реакцией) телята остаются иммунологически незащищенными и восприимчивыми к патогенным микроорганизмам. Следовательно, здоровье телят зависит от поглощения и абсорбции колостральных иммуноглобулинов, обеспечивающих максимальную резистентность от инфекционных заболеваний в течение первой недели жизни [T.C. McGuire, D.S. Adams, 1982].
Известно, что телята способные абсорбировать иммуноглобулины наиболее эффективно в течение первых 8 ч после рождения с незначительным снижением уровня в течение 24 ч. Это происходит в результате «закрытия» интестинального эпителия и прекращения активного транспортного механизма [G. H. Stott, D. B. Marx, B. E. Menefel and G. T. Nightengale, 1979]. Кроме того, иммуноглобулиновый уровень у коров снижается быстро после родов, в течение первых трех дней. Поэтому необходимость раннего поглощения молозива является неотъемлемым элементом системы выращивания телят.
Новорожденные телята очень лимитированы гликогеном (животный крахмал), который накапливается в печени. Он должен использоваться как адекватный источник энергии после рождения для поддержания позитивного энергетического уровня. Высокий уровень циркуляции в крови телят молозива перед рождением до некоторой степени обеспечивает глюконеогенез (образование глюкозы из протеина и жира) в первые 12 ч.
Молозиво превосходный источник энергии, протеина, витаминов А и D, комплекса витаминов В, а также кальция, фосфора, магния и хлоридов.
В наших исследованиях (учхоз «Кетросу» района Анений Ной, ОПХ «Колоница») 26,6 % (4/15) телят, лишенных молозива, выживали, в то время как ни один теленок, кормившийся молозивом не погиб. На фермах применялось стандартное лечение, включая заменитель молока с электролитным раствором 3 дня и более и последующие 2 дня реадаптации — заменитель молока, когда диарея была корректирована (нейтрализована). Антибиотики орально не применяли для того, чтобы не уничтожить нормальную бактериальную популяцию и не вызвать фунгальный (грибной) гастроэнтерит. Значительная перегрузка организма телят антибиотиками может содействовать повышению восприимчивости к фунгальной инфекции, не поддающейся лечению[М. А. Сидоров, 1981; У. Риихикоски, 1986].
2.2.11. Клиническая оценка эффективности вакцинации коров для профилактики колибактериоза телят. Колибактериоз телят относится к числу широко распространенных инфекционных болезней молодняка и регистрируется во всех развитых странах. Наиболее ощутимый ущерб он наносит в животноводческих комплексах за счет высокой смертности, отставания в росте и снижения продуктивности переболевших животных. Одним из основных предрасполагающих факторов колибактериоза является агамма- или гипогаммаглобулинемия, обусловленные неполноценностью молозива полученного и скармливаемого несвоевременно новорожденным в условиях снижающих резистентность макроорганизма [Ю. А. Малахов, О. А. Тугаринов, М. К. Пирожков, Т. И. Исхакова, 1993].
Заболевание телят колибактериозом на 2-5 дни жизни вынуждает вакцинировать против эшерихиоза стельных коров для создания невосприимчивости у нарождающегося молодняка. Против колибактериоза вакцинируют животных во всех развитых странах. Первые вакцины колибактериоза телят начали применять в 40-х годах. Они представляли инактивированную микробную суспензию штаммов кишечной палочки, выделенных от больных или павших животных и использовались в хозяйствах различных регионов с незначительной эффективностью. Для профилактики колибактериоза в настоящее время применяют поливалентную гидрооксиалюминевую формол-тиомерсаловую вакцину против колибактериоза (эшерихиоза) поросят, телят и ягнят. Изготовляют два варианта:
Вакцина против колибактериоза поросят: О8:К43, О9:К30, О78:К80, О138:К81, О139:К82, О141:К85:К88, О147:К87:К88, О149:К91:К88.
Вакцина против колибактериоза телят и ягнят: О8:К43, О9:К30, О15:К14, :К30, О20, О26:К60, О41, О55, О78:К80, О86:К61, О115, О117:К62, О119:К69, О101:К99.
На фоне применения данной вакцины мы предусмотрели:
Установить серотиповую принадлежность эшерихий, выделенных от больных и павших телят.
Определить уровень антител, приобретенный телятами в результате вакцинации стельных коров в сыворотке молозива и сыворотке крови телят против выделенных серотипов кишечной палочки.
Разработать экспресс-метод для индикации «К» антител в колостральной сыворотке коров и сыворотке крови новорожденных телят.
Для выделения культур Е. coli отбирали патологический материал от павших телят, а от больных фецес, из хозяйств неблагополучных по колибактериозу, Культуры выделяли и идентифицировали в соответствии с «Методическими рекомендациями по бактериологической диагностике колибактериоза» (1981; 1991).
Принадлежность исследуемых культур Е. coli к О серогруппам устанавливали в РА с типоспецифическими О колисыворотками согласно наставлению по их применению.
Способность Е. coli образовывать фимбриальные адгезины определяли в капельной РА с моноспецифическими антиадгезивными сыворотками К99, F41, Аtt25, 987Р, К88 ав, К88 ас, К88 аd.[О. А. Полякова, Н. И. Евглевская, Н. А. Соколова, 1986].
Вакцинацию стельных коров проводили «Поливалентной гидроокисиалюминиевой формол-тиомерсаловой вакциной против колибактериоза (эшерихиоза) телят и ягнят согласно наставлению по ее применению.
Пробы фецес разводили стерильным физиологическим раствором 1:100, затем пастеровской пипеткой засевали две капли взвеси кала, культивировали на 5 %-ном кровяном агаре, Мак Конки агаре, средах Эндо и Левина при 37°С 18 ч. Е. coli пересевали со сред Эндо и Левина на МПА и МПБ. Полученные агаровые и бульонные культуры выдерживали в темном месте при комнатной температуре для дальнейшего исследования и серотипизации.
Для бактериологического исследования отбирали следующий материал от падших телят: сычуг, верхний отрезок тощей кишки, нижний отрезок тощей кишки, части подвздошной, слепой кишки, мезентеральные лимфатические узлы, печень, селезенку и почки. Пробы отбирали также из других мест, таких как суставы, головной мозг, пуповина и легкие, если клинически и патологоанатомически выявляли аномалии в них. Пробы отбирали, избегая контаминации и культивировали на 5 %-ном кровяном агаре и Мак Конки агаре. Пробы подвздошной кишки и селезенки культивировали на среде Левина. Засеянные пробирки и чашки Петри инкубировали при обычной атмосфере (аэробные условия) при 37°С 18 ч. Три колонии из преобладающих типичных колоний Е. coli были пересеяны дополнительно на среды с углеводами (глюкоза, лактоза, сахароза, манит и др.). Пробирки инкубировали при 37° С 18 ч.
Критериями для установления диагноза на колибактериоз были: клиническое и патологоанатомические исследования, отличающиеся от других случаев диареи и гибели телят и которые ассоциировались со следующими бактериологическими данными:
большое количество Е. coli в верхнем отрезке кишечника и региональных лимфатических узлах, а также в других интестинальных органах;
отсутствие других патогенных бактерий, поражающих телят.
2.2.11.1. Определение «О» антигена. Выделенные культуры классифицировали серологически в порядке сравнения с серотипами, применяемыми при вакцинации. Культуру в начале тестировали пластинчатой реакцией агглютинации на предметном стекле, 18-часовую культуру из плотной питательной среды (МПА) смывали физиологическим раствором и полученную суспензию использовали как антиген. Эту суспензию нагревали при 100° С один час для инактивирования В и L типов «К» антигена и тестировали, смешивая одну каплю антигена с одной каплей соответствующей «О» сыворотки на зеркальном стекле, размещая его над водяной баней при 60° С. Результаты учитывали через 10 мин после смешивания. Позитивную реакцию подтверждали пробирочной агглютинацией, применяя формалинизированную шестичасовую культуру в МПБ.
2.2.11.2. Определение «К» антигена. Культуру вначале тестировали пластинчатой реакцией агглютинации на предметном стекле. Одну каплю живой суспензии культуры смешивали с одной каплей различных антисывороток. Результаты учитывали в течение 30 секунд после смешивания. Позитивную агглютинацию подтверждали пробирочной агглютинацией до титра тест сыворотки, применяя как антиген живую пятичасовую культуру в МПБ. Пробирки инкубировали при 37° С два часа, в последующем агглютинационные пробирки центрифугировали при 2000 об/мин в течение трех минут.
Одинаковый диск агглютинации учитывали как позитивную реакцию.
Эффективность вакцинации коров, предназначенную для профилактики колибактериоза телят определяли в двух хозяйствах.
Проведенная серологическая типизация выделенных культур Е. coli от павших телят, родившихся от вакцинированных коров, свидетельствует, что, как правило, серологическая принадлежность в сравнении с вакцинными серотипами имеет существенное различие (табл. 2.25).
Таблица 2.25
Данные серологической типизации по «О» антигену Е. coli
|
Вакцинный серотип |
Период исследования |
Количество павших телят от коров |
Изолированные серотипы в группе |
||
|
вакцинированных |
невакцинированных |
вакцинированной |
контрольной |
||
|
О8:К43 О9:К30 О15:К14:К30 О20 |
Ноябрь-декабрь 1991 г. |
3 |
2 |
2/3* |
3 |
|
О26:К60 О41 О55 О78:К80 О86:К61 О115 |
Ноябрь-декабрь 1992г. |
2 |
3 |
1/3 |
2/2 |
|
О117:К62 О119:К69 О101:К99 |
Октябрь-декабрь 1993 г. |
3 |
2 |
1/2 |
1/2 |
* числитель — типируемые; знаменатель — нетипируемые.
Из 12 родившихся телят в октябре-декабре 1993 года семь были от вакцинированных коров, и пять от невакцинированных. У четырех телят из пяти от невакцинированных коров наблюдали диарею и из них два погибли. Из семи телят от вакцинированных коров пять заболели на 2-3-й день жизни, три из них погибли.
Нетипируемые штаммы Е. coli . были обнаружены во всех уровнях кишечника за исключением внутренних органов. Кроме того, нетипируемые штаммы Е. coli выделяли из фекальных проб телят вначале диареи.
Из других органов (печень, селезенка) выделяли штаммы Е. coli обладающие «О» групповым антигеном, принадлежащими к серотипам О101; О8.
Таблица 2.26
Результаты серологической типизации по «К» антигену Е. coli
|
Возраст телят (дней) |
Органы, из которых выделены Е. coli (всего / адгезивнопозитивных) |
||||||||
|
печень |
селезенка |
почки |
сычуг |
верхний отрезок тощей кишки |
нижний отрезок тощей кишки |
подвздошная кишка |
слепая кишка |
лимфоузлы |
|
|
2 |
1/1 |
1/0 |
- |
- |
1/1 |
- |
- |
- |
1/1 |
|
3 |
1/0 |
1/0 |
1/1 |
- |
1/1 |
- |
- |
- |
- |
|
4 |
1/0 |
1/0 |
1/1 |
- |
1/1 |
- |
- |
- |
- |
|
5 |
1/1 |
1/0 |
- |
- |
- |
- |
1/1 |
- |
1/0 |
У выделенных от телят Е. coli обнаруживали фимбрии К99, К88ас. (табл. 2.26). При анализе О серогрупповой принадлежности исследуемых культур и способности экспрессировать фимбрии установили взаимозависимость. Так, фимбрии обнаруживали у выделенных от телят штаммов Е. coli относящихся к серотипам О8 и О101.
Уровень антител молозива, полученного от вакцинированных коров и уровень антител сыворотки крови телят от этих коров представлены в табл. 2.27; 2.28.
Таблица 2.27
Титр антител в молозиве коров и сыворотке крови телят (серотип О8) Е. coli
|
Возраст телят (дней) |
Сыворотка молозива* |
Сыворотка крови телят |
Исход** |
||
|
«О» антиген |
«К» антиген |
«О» антиген |
«К» антиген |
||
|
2 |
20*** |
0 |
10 |
0 |
пал |
|
3 |
80 |
10 |
20 |
0 |
выздоровел |
|
4 |
40 |
0 |
0 |
0 |
пал |
|
5 |
20 |
0 |
0 |
0 |
пал |
|
5 |
160 |
10 |
40 |
10 |
выздоровел |
* сыворотка молозива первого дня после отела.
** теленок пал или выздоровел.
*** обратная величина дроби.
Таблица 2.28
Титр антител в молозиве коров и сыворотке крови телят (серотип О101) Е. coli
|
Возраст телят (дней) |
Сыворотка молозива |
Сыворотка крови телят |
Исход |
||
|
«О» антиген |
«К» антиген |
«О» антиген |
«К» антиген |
||
|
3 |
20 |
0 |
0 |
0 |
пал |
|
2 |
10 |
0 |
0 |
0 |
пал |
|
3 |
40 |
0 |
10 |
0 |
выздоровел |
|
4 |
20 |
0 |
0 |
0 |
пал |
|
4 |
40 |
0 |
20 |
10 |
выздоровел |
Данные таблицы подтверждают, что вакцинация стельных коров не ингибирует серотипы Е. coli входящие в состав вакцины. Прекращение вспышек колибактериоза связано с другими факторам.
В наших исследованиях мы установили, что колибактериоз телят вызывался двумя серотипами Е. coli О8 и О101. Более ранние наблюдения [A. Dam, 1971; H. Fey, 1971a] в других странах также указывают, что более чем один серотип Е. coli может быть изолирован при заболевании телят колибактериозом.
Известно, что «К» антитела играют важную роль в защите лабораторных животных против экспериментальной Е. coli инфекции [R. K. Braun, B. C. Tennant, 1983]. Большинство тестированных телят в наших исследованиях не обладали «К» антителами к Е. coli штаммов изолированных при заболевании телят колибактериозом.
ВЫВОДЫ
Содержание колостральных иммуноглобулинов в сыворотке крови телят отражает уровень резистентности животных.
Концентрация иммунных глобулинов в молозиве в течение первых трех дней лактации снижается последовательно в 2,16 раза на второй день и в 4,93 раза на третий – по сравнению с первым днем.
Уменьшение абсорбции колостральных иммуноглобулинов в 1,23 и в 1,45 раза по сравнению с контрольной группой (здоровые) животных способствует возникновению соответственно энтеритной и септической формы колибактериоза.
Угасание абсорбции колостральный антител происходит в следующей последовательности IgG, IgM и IgA.
Количественное содержание иммуноглобулинов в сыворотке крови и в сыворотке коров имеет существенное различие.
Активность антител против К-антигенов в молозиве коров и в сыворотке крови телят 1-2-дневного возраста была меньшей, чем активность против О-антигенов Е. coli.
Различная концентрация антител (титр) в сыворотке новорожденных телят против О и К-антигенов Е. coli указывает, что кормление молозивом может стимулировать или подавлять синтез антител к определенным антигенам в одинаковой степени независимо от места их локализации в бактериальных клетках.
Протективный механизм является иммунологическим фактором и коррелирует с уровнем иммуноглобулинов против К-антигенов Е. coli и возникновением колибактериоза телят.
Иммунизация стельных коров поливалентной гидроокисьалюминиеной формол-тиомерсаловой вакциной против колибактериоза телят не ингибирует серотипы Е. coli 08 и 0101, входящие в состав вакцины и выделенные от павших телят.
РАЗДЕЛ 3 ИЗУЧЕНИЕ НЕСПЕЦИФИЧЕСКОЙ ИММУНОЛОГИЧЕСКОЙ РЕАКТИВНОСТИ НОВОРОЖДЕННЫХ ТЕЛяТ
3.1. Биохимический анализ крови новорожденных телят
В опытах использовали 30 телят. Их возраст в период исследования был от 2 до 10 дней. Кровь брали из яремной вены для определения следующих показателей:
рН;
СО>2> (Hg mm);
О>2> (Нg mm);
НСО>3> (m Eg/I);
N+ (m Еg/I);
K+ (m Eg/I-);
Cl (m Eg/I);
Гематокрит (%);
Мочевина (mg/dl);
Неорганический фосфор (mg/dl);
Общий белой (g/dl).
Анализ этих показателей проводили дважды в день. В порядке изучения отклонений учитывали два условия: выживание и смерть телят. Это состояние определяли функцией учтенных параметров как максимум различий, выступающий между двумя группами.
Характеристика 2-х показателей приведена в табл. 3.1; 3.2; 3.3. Данные таблиц показывают, что некомпенсированный метаболический ацидоз всегда присутствует. Дегидратация отмечается при высоком гематокрите.
Корреляционные табл. 3.1 и 3.2 между основными показателями указывают, что определенные параметры имеют очень сходное поведение (например, 1 и 4 бикарбонатная концентрация изменяется непосредственно в зависимости от рН крови, феномен — хорошо известный). На другие параметры передается их обратная связь, как например, частичное соприкосновение с противоположной связью СО>2> и О>2>.
Корреляционная матрица (табл. 3.3-3.5) показывает на сопротивление между парциальным давлением О>2> в венозной крови и показателями кислотно-щелочного равновесия, в частности парциального давления СО>2> в порядке нейтрализации метаболического ацидоза в нормальном противодействии уменьшающим парциальное давление СО>2>. Следовательно, снижение парциального давления СО>2> указывает на уменьшение количества О>2> в тканях.
Группа результатов подтверждает, что некомпенсированный метаболический ацидоз вызывает диарею у телят. Существует связь между кислотно-щелочным равновесием и катаболизмом. Вероятно, что катаболизм, который имеет место при диарее, практически выражает плохо функционирующие физиологические механизмы. Кислотность убывает в основном, с фецес и это состояние прямое следствие диареи.
Моделью определения основных показателей является математическая обработка сравнимых данных, полученных в этом исследовании.
При статистической обработки результатов исследовании с применением ЭВМ «НАИРИ» крови новорожденных телят, установлено, что использование трех показателей легко измеряемых величин (мочевина, гематокрит, хлориды) позволяет прогнозировать с вероятной точностью до 78% возникновения заболевания и, следовательно, лечить с применением регидратационной терапии, возмещением кислотности и потери ионов натрия.
Некомпенсированный метаболический ацидоз – основной признак при диареи телят. Установлена связь между кислотно-щелочным равновесием и катаболизмом. Возможно, что катаболизм отражает плохо функционирующие физиологические механизмы, воздействующие на причинный агент (бактерии, вирусы).
Данные исследования показывают, что выжившие телята имеют преимущество в количественном отношении изучаемых величин перед павшими животными. Концентрация калия (мера катаболизма) имеет тенденцию к увеличению, в то время как концентрация натрия в крови уменьшается (табл. 3.6-3.7).
Таблица 3.3
Корреляционная матрица биохимических показателей нормальное значение
|
pН X>1> |
pCO>2> X>2> |
pO>2> X>3> |
CO>3>H X>4> |
Na+ X>5> |
K+ X>6> |
Cl X>7> |
PCV X>8> |
Urea X>9> |
P X>10> |
|
|
X>2> pCO>2> |
0,081 |
|||||||||
|
X>3> pO>2> |
0,202 |
0,808 |
||||||||
|
X>4> CO>3>H |
0,251 |
0,739 |
0,858 |
|||||||
|
X>5> Na+ |
0,408 |
0,749 |
0,823 |
0,895 |
||||||
|
X>6> K+ |
0,244 |
0,515 |
0,542 |
0,457 |
0,534 |
|||||
|
X>7> Cl |
0,050 |
0,010 |
0,226 |
0,0007 |
0,109 |
0,000 |
||||
|
X>8> PCV |
0,247 |
0,192 |
0,395 |
0,169 |
0,133 |
0,148 |
0,642 |
|||
|
X>9> Urea |
0,168 |
0,055 |
0,112 |
0,230 |
0,121 |
0,282 |
0,633 |
0,844 |
||
|
X>10> P |
0,191 |
0,532 |
0,705 |
0,581 |
0,684 |
0,142 |
0,364 |
0,737 |
0,697 |
|
|
X>11> Proteins |
0,124 |
0,181 |
0,228 |
0,047 |
0,047 |
0,411 |
0,463 |
0,652 |
0,597 |
0,579 |
Таблица 3.4
Корреляционная матрица между изучаемыми отклонениями (выжившие телята)
|
pН Y>1> |
pCO>2> Y>2> |
pO>2> Y>3> |
CO>3>H Y>4> |
Na+ Y>5> |
K+ Y>6> |
Cl Y>7> |
PCV Y>8> |
Urea Y>9> |
P Y>10> |
|
|
Y>2> pCO>2> |
-0,115 |
|||||||||
|
Y>3> pO>2> |
-0,224 |
0,811 |
||||||||
|
Y>4> CO>3>H |
-0,267 |
0,726 |
0,888 |
|||||||
|
Y>5> Na+ |
-0,113 |
0,453 |
0,511 |
0,660 |
||||||
|
Y>6> K+ |
-0,266 |
0,343 |
0,486 |
0,372 |
0,141 |
|||||
|
Y>7> Cl |
-0,086 |
0,123 |
0,165 |
0,0927 |
-0,090 |
0,830 |
||||
|
Y>8> PCV |
-0,000 |
0,253 |
0,364 |
0,192 |
0,084 |
0,571 |
0,652 |
|||
|
Y>9> Urea |
-0,174 |
0,334 |
0,503 |
0,458 |
0,016 |
0,619 |
0,713 |
0,773 |
||
|
Y>10> P |
-0,012 |
0,457 |
0,448 |
0,486 |
0,350 |
0,358 |
0,493 |
0,701 |
0,703 |
|
|
Y>11> Proteins |
-0,488 |
0,249 |
0,194 |
0,141 |
0,232 |
0,353 |
0,453 |
0,580 |
0,408 |
0,684 |
Таблица 3.5
Корреляционная матрица между изучаемыми отклонениями (павшие телята)
|
pН Y>1> |
pCO>2> Y>2> |
pO>2> Y>3> |
CO>3>H Y>4> |
Na+ Y>5> |
K+ Y>6> |
Cl Y>7> |
PCV Y>8> |
Urea Y>9> |
P Y>10> |
|
|
Y>2> pCO>2> |
-0,341 |
|||||||||
|
Y>3> pO>2> |
-0,231 |
0,514 |
||||||||
|
Y>4> CO>3>H |
-0,035 |
0,678 |
0,532 |
|||||||
|
Y>5> Na+ |
-0,160 |
0,720 |
0,613 |
0,753 |
||||||
|
Y>6> K+ |
-0,170 |
0,485 |
0,435 |
0,093 |
0,352 |
|||||
|
Y>7> Cl |
-0,231 |
0,073 |
0,460 |
-0,265 |
-0,035 |
0,455 |
||||
|
Y>8> PCV |
-0,131 |
0,195 |
0,224 |
0,209 |
0,283 |
0,149 |
0,415 |
|||
|
Y>9> Urea |
-0,550 |
0,048 |
0,064 |
-0,130 |
0,147 |
0,120 |
0,294 |
0,231 |
||
|
Y>10> P |
-0,345 |
0,209 |
0,094 |
0,239 |
0,474 |
0,043 |
0,115 |
0,416 |
0,473 |
|
|
Y>11> Proteins |
-0,182 |
0,108 |
0,353 |
0,307 |
0,241 |
-0,198 |
0,219 |
0,764 |
0,159 |
0,103 |
Таблица 3.6
Дисперсионный анализ изучаемых отклонений (выжившие телята)
|
рН Y1 |
PCO2 Y2 |
pO2 Y3 |
CO3H Y4 |
Na+ Y5 |
K+ Y6 |
Cl Y7 |
PCV Y8 |
Ureea Y9 |
P Y10 |
Proteins Y11 |
|
|
X>1>> >pН |
170,186 |
||||||||||
|
X>2> pCO>2> |
448,045 |
||||||||||
|
X>3> pO>2> |
336,005 |
||||||||||
|
X>4> CO>3>H |
8767,769 |
||||||||||
|
X>5> Na+ |
526,250 |
||||||||||
|
X>6> K+ |
98,543 |
||||||||||
|
X>7> Cl |
3,647 |
||||||||||
|
X>8> PCV |
31,369 |
||||||||||
|
X>9> Urea |
51,209 |
||||||||||
|
X>10> P |
51,599 |
||||||||||
|
X>11> Proteins |
55,281 |
Таблица 3.7
Дисперсионный анализ изучаемых отклонений (павшие телята)
|
рН Y1 |
PCO>2> Y2 |
pO>2> Y3 |
CO>3>H Y4 |
Na+ Y5 |
K+ Y6 |
Cl Y7 |
PCV Y8 |
Ureea Y9 |
P Y10 |
Proteins Y11 |
|
|
X1 pH |
32,49 |
||||||||||
|
X2 pCO>2> |
37,85 |
||||||||||
|
X3 pO>2> |
10,019 |
||||||||||
|
X4 CO3H |
42,259 |
||||||||||
|
X5 Na+ |
3,408 |
||||||||||
|
X6 K+ |
56,822 |
||||||||||
|
X7 Cl |
19,709 |
||||||||||
|
X8 PCV |
27,002 |
||||||||||
|
X9 Ureea |
36,472 |
||||||||||
|
X10 P |
35,284 |
||||||||||
|
X11 Proteins |
27,359 |
3.2. Коррекция кислотно-основного баланса при диареи неонатальных телят
Как правило, лечение диареи неонатальных телят направлено на, непосредственно, идентифицированные микроорганизмы с последующей элиминацией их при применении антибиотиков и химиотерапевтических препаратов. Тем не менее, многие животные не поддаются лечению, а в некоторых случаях увеличивается патогенность микроорганизмов.
При диареи телят увеличивается масса фецесс до 40 %, главным образом, за счет увеличения экскреции воды, что сопровождается значительной потерей кальция, магния, натрия и калия. Экстенсивная потеря внутриклеточного калия сопровождается выходом натрия, в течение дегидратации при диарее или в течение лечения, если калий не включается в замещающий раствор. Особенно, подвергаются потери хлорида натрия в диарейных испражнениях, так как их компоненты секретируются в гастроинтестинальном тракте[В. В. Митюшин, 1984].
Система бикарбонатно-угольная кислота является одной из наиболее важных буферных систем в организме и изменение в этой системе рефлекторно изменяет кислотно-основной баланс в целом организме. Сохранение кислотности или щелочности (основности) вызывает рефлекторное изменение бикарбонатов в сыворотке в смеси угольная кислота (Н>2>СО>3>) и бикарбонаты (ВНСО>3>) , основу которых составляют, главным образом, натрий или калий.
Согласно формуле Гендерсона и Гассельбаха, рассчитанной на основании константы диссоциации Н>2>СО>3> (К= 6, I) и соответствия растворимых в плазме бикарбонатов (27 ммоль/л) и угольной кислоты (1,35 ммоль/л), рН крови равен:
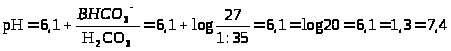
Нормальное соотношение угольной кислоты и бикарбонатам равняется 1:20. Если рН сыворотки известно, кислото (угольная) бикарбонатное отношение определяется также по Гендерсона-Гассельбаха уравнению. Таким образом, при рН 7,4 отношение составляет 1:20, при рН 7,2 1:12, при рН 7,1 падает до 1:10. Следовательно, если рН ниже чем 7,3 то будет не компенсированный ацидоз.
При нормальном состоянии, натрий в клетках находится в незначительной концентрации, при аномальном состоянии может поступать в клетки в увеличенном количестве, замещая калий. Калий выделяется из клетки и поступает в кровь и если почечная функция нормальная избыток выделяется в мочу. Высокий уровень калия в сыворотке приводит к очень значительному дефициту калия в организме [E. W. Fisher and de la G. H. Fuente, 1972; А. Г. Шитый, Н. С. Дудникова, Л. М. Тихомирова, 1993].
Дегидратацию при диареи учитывают как неотложную для замещения потерянной жидкости парентеральным введением растворов. Применяют исключительно 0,9 % раствор хлорида натрия для этой цели не без опасности содержания избытка ионов хлорида и недостатка других ионов, удаленных при таком состоянии.
В настоящем исследовании преследуются две цели:
первая изучить и зарегистрировать изменения в кислотно-основном уровне электролитов в сыворотке телят при диареи;
вторая изучить влияние при парентеральном введении электролитных растворов при этих изменениях.
Значение гематокрита получали, применяя гематокритные пробирки. Центрифугировали при 2500 об/мин в течение 30 мин. Сахар крови определяли в 0,05 мл цельной крови. Уровень изучаемых показателей определяли у 10 телят 4-6 дневного возраста. Балансирующий электролитный раствор применяли для парентеральной терапии в течение этого исследования [Е. Рашкова, Т. Сехер, К. Рашка, В. Матейовска, Л. Палак, 1976; J. Blanu, J. H. Parvu, O. Ivanciu, 1983].
Случай 1. Телята (4) 4 дневного возраста. Санитарное состояние на ферме неудовлетворительное. В первый день животные были вялые и сонные, не способные ходить и питаться, вставали с трудом. Обрабатывали химиотерапевтическими препаратами и вводили один литр раствора электролита интраперитонеально. На второй день животные выздоравливали.
Случай 2. Телята (4) 5 дневного возраста. В первый день животные были вялыми и летаргическими с профузной диареей, на второй день телята были неспособные подниматься. Антибиотикотерапию проводили на первый, второй и третий день. Один литр электролитного раствора вводили в первый день и два литра на второй день. На третий день диарея была менее явной, на 4 день фецес были почти нормальными.
Случай 3. Телята (2) 6 дневного возраста болели диареей в неопределенное предшествующее время. Антибиотикотерапия не улучшала общее состояние. Телята были в коматозном состоянии с холодным липким ртом, поверхностным дыханием и стекловидными запавшими глазами. Два литра электролитного раствора вводили в первый день. Утром на второй день животные были бодрыми, способными передвигаться и пить энергично молоко. Температура была 39,2° С, фецес были жидкими, но не диарейными.
Анализ образцов крови, сыворотки и уровень гематокрита указывает, что в случае 1 увеличивается калий и натрий в сыворотке крови, несмотря на краткое течение заболевания. В случае втором, электролитное изменение находилось в соответствии с клиническим состоянием. Сывороточные бикарбонаты, хлориды и кальций были ниже нормы, а сывороточный калий выше нормы, когда симптомы были тяжелыми и приходили к нормальному значению с улучшением клинического состояния. В случае 3 сывороточный бикарбонат, кальций и натрий были ниже нормы, а сывороточный калий и неорганический фосфор (12,5мг/100 мл) были увеличены , когда клиническое состояние было тяжелым. В течение 48 ч лечения с восстанавливающим раствором электролитные образцы и клиническое состояние были нормальными.
Наиболее постоянным показателем в трех случаях было уменьшение в сывороточном бикарбонатном уровне, сопровождающееся низким рН сыворотки.
В двух случаях (2, 3) рН была ниже 7,3 в первый день заболевания и превалировало состояние не компенсаторного ацидоза. Компенсация в большинстве случаев поддерживалась до бикарбонатного уменьшения в пределах 20. Вероятно, ацидоза будет избыточным при сохранении кислотности и потери щелочности. Характерной особенностью в изучаемых случаях было отклонение от нормы уровня сывороточного калия, который был увеличен во всех случаях в начальной стадии заболевания. Дефицит калия в клетках вызывает состояние нечувствительности (оцепенение, коматозное состояние). Является общепризнанным фактом, что с уменьшением бикарбонатов, происходит повышение хлоридов, которые компенсируют бикарбонаты. Уровень глюкозы во втором и третьем случаях был ниже, чем 10мг/100мл.
Уровень гематокрита указывал на степень дегидратации. Следовательно, раннее применением антибиотиков и химиотерапевтических препаратов, контролирующих диарею, предотвращает электролитное изменение. При экстенсивных отклонениях сывороточных рН, бикарбонатов, хлоридов, натрия, калия и кальция лечение антибиотиками и химиотерапевтическими препаратами является бесперспективным.
3.3. Изменение фагоцитарныхных показателей у новорожденных телят в зависимости от клинического статуса
Общая невосприимчивость животных к неблагоприятным факторам внешней и внутренней среды обусловлена иммунобиологической реактивностью организма. Известно, что неспецифические защитные факторы организма детерминируются генетическим характером и могут быть выражены с различной силой в зависимости от физиологического состояния животных, сезона года, условий кормления и содержания [С. J. Howard, A. A. Glynn, 1971; В. В. Никольский, В. П. Литвин, 1974; В. Я. Мозгис, 1982; В. Т. Сидоров, 1984; В. П. Павличенко, О. В. Милованов, Н. Н. Берникова, 1985].
Новорожденные животные обладают незначительной устойчивостью к возбудителям инфекционных заболеваний. До выпойки молозива основную защитную функцию выполняют клеточные защитные факторы. Важнейшим фактором к клеточной защитной системе организма является опсонофагоцитарная реакция лейкоцитов.
Целью исследования было изучение фагоцитарных показателей у новорожденных телят в онтогенезе, и их взаимосвязь с сохранностью животных в условиях хозяйств республики.
Изучали показатели клеточной защиты организма новорожденных телят: фагоцитарную активность и интенсивность. При постановке опсонофагоцитарной реакции а качестве тест культуры использовали серотип О78 Е. coli . Бактериальную взвесь готовили из 24 ч агаровой тест культуры путем смыва физиологическим раствором и по оптическому стандарту мутности устанавливали концентрацию 2 млрд. микробных тел в 1 мл.
В мазках подсчитывали 50 нейтрофилов и количество захваченных ими микробов, после чего вычисляли фагоцитарную активность и фагоцитарную интенсивность [В. Я. Мозгис, 1982].
В результате проведенных опытов на телятах от рождения до 30-днеаного возраста было установлено, что новорожденный организм обладает достаточно выраженной клеточной защитной реакцией (табл. 3.8-3.15) Фагоцитарная активность лейкоцитов у 2-дневных телят составляла (в %) 44,78±0,58, фагоцитарная интенсивность 1,94±0,02. Показатели активности и интенсивности фагоцитоза повышались до 5-дневного возраста, затем в возрасте 10 дней наблюдалось их снижение. Количество лейкоцитов, участвующих в фагоцитозе, в этот период снизилось до 41,67 ± 0,65, что на 10,66 % (Р<0,05) ниже, чем у 5-дневных телят.


В первой опытной группе (больные телята) фагоцитарная активность на 2-й,. 5-й, 10-й день имеет существенные различия, т. е. наблюдается достоверное снижение, как фагоцитарной активности, так и интенсивности соответственно на 6,67; 11,33 и на 6,34 % фагоцитарной активности и в этот же наблюдаемый период отмечали снижение фагоцитарной интенсивности в 1,02, 1,09 и в 1,01 раза по сравнению с контрольной группой.
Во второй подопытной группе (больные-павшие) фагоцитарные показатели: активность н интенсивность имеет существенное различие на 2-й и 5-й день возрастного периода новорожденных телят. Так, фагоцитарная активность на 2-й день в контрольной группе была 44,78±0,58, во второй опытной группе 24,57±0,43; на 5-й день соответственно 52,33±0,60 и 23,50±0,15. Следовательно, количество лейкоцитов, участвующих в фагоцитозе уменьшилось на 20,2 % на 2-й день и на 28,83 % на 5-й день. Фагоцитарная интенсивность в контрольной и во 2-й подопытной группе также имеет существенные различия. Так, на 2-й день в контрольной группе этот показатель равнялся 1,94±0,02, а во второй опытной группе 1,79±0,004, т. е. в 1,08 раза меньше, на 5-й день это значение, также достоверно было различимо 2,32±0,02 и 1,77±0,016, т. е. фагоцитарная интенсивность в этот период снизилась в 1,З1 раза (Р<0,05), по сравнению с контрольной группой. Уровень фагоцитарной активности и фагоцитарной интенсивности в крови телят в послемолозивный период статистически достоверно повышались до 5-дневного возраста в контрольной группе (здоровые) соответственно +7,55 и +0,38( Р<0,001) и первой подопытной +2,89 и + 0,23 (Р<0,001). Во второй подопытной группе достоверное повышение фагоцитарных показателей отмечали при сравнении с их уровнем в период до поения молозива. Следовательно, лейкоцитарный фагоцитоз является важнейшим показателем иммунобиологического состояния. Так, снижение фагоцитарных показателей фагоцитарной активности на 6,67 % (2-й день) и 11,33 % 5-й день и соответственно фагоцитарной интенсивности в 1,02 и 1,09 раза на 2-й и 5-й дни указывает на высокий риск заболеваемости. При снижении фагоцитарной активности на 20,21 (2-й день) и 28,83 % (5-й день) по сравнению с контрольной группой (здоровые) неизбежно приводит к гибели новорожденных животных.
3.4. Бактерицидная активность сыворотки крови новорожденных телят
Состояние естественной резистентности организма наиболее полно характеризует бактерицидная активность сыворотки крови, которая заключается в способности подавлять рост микроорганизмов и зависит от активности всех гуморальных факторов неспецифической устойчивости. Бактерицидную активность сыворотки крови используют как один из критериев оценки тяжести состояния организма при инфекционных болезнях и эффективности проводимой терапии в ветеринарной практике[П. А. Емельяненко, 1979; В. В. Биктимиров, 1993].
Цель наших исследований определить уровень бактерицидной активности сыворотки крови у новорожденных телят в динамике с момента рождения (до выпойки молозива) до 30-дневного возраста и предложить иммуностимуляторы, обеспечивающие высокую эффективность проводимой терапии.Бактерицидную активность сыворотки крови определяли по О. В. Смирновой и А. Т. Кузьминой (1966). В качестве тест микробов использовали S форму Е. coli.
Для измерения ингибирования роста Е. coli в бульоне Хоттингера при добавлении сыворотки крови, культуру высевали в МПБ. В пробирку стерильно разливали по 4,5 мл бульона Хоттингера, добавляли по 1 мл испытуемой сыворотки и по 0,1 мл взвеси суточной культуры. Две пробирки со средой оставляли в качестве контроля. Из каждой опытной и контрольной пробы стерильно отбирали по 2 мл и определяли их исходную оптическую плотность (Ао/исх и Ак/исх) на ФЭК-56 М при длине волны 364 нм (красный светофильтр), кювета 3 мм против бульона Хоттингера (оптический контроль)[Н. Н. Максимюк, Л. Я. Телешевская, 1995].
По классическому методу оптическую плотность измеряют при зеленом светофильтре. При использовании зеленого светофильтра, регистрируется окраска среды. Для исключения влияния этого фактора необходим дополнительный оптический контроль для каждой пробы в начале опыта. Чем больше гемолиз сыворотки, тем сильнее окраска среды и выше неспецифическое поглощение. Применение для этой цели красного светофильтра исключает влияние окраски среды. Пробы культивировали 6 ч при 37°С и рассчитывали бактерицидную активность в %.
Бактерицидную активность определяли как отношение роста микроорганизмов в опытной среде по сравнению с контрольной (без сыворотки) в процентах.
В результате проведенных исследований было установлено, что сыворотка крови новорожденных телят обладает выраженной бактерицидной активностью (табл. 3.16-3.19).
Бактерицидная активность сыворотки крови телят зависит от возраста, причем наименьший ее уровень отмечали у новорожденных телят до приема молозива.
На 2-й день жизни телят имело место достоверное повышение бактерицидной активности сыворотки у здоровых телят на 12,09 %, в 1-ой подопытной группе (больные) на 6,09 % и снижение во 2-й подопытной группе на 1,58 % в сравнении с периодом по выпойки молозива. На 5-й и 10-й дни отмечали повышение бактерицидной активности у здоровых телят в сравнении с периодом до поения молозива, соответственно на 14,09 % и 14,18 %, в 1-ой подопытной группе соответственно на 19,18 % и 9,91 %. Во 2-й подопытной группе отмечали снижение бактерицидной активности на 5-й день в сравнении с периодом до выпойки молозива на 1,86 %.
Данные исследования указывают, что наиболее критический возрастной период отмечен у телят 5- дневного возраста, так в 1-й опытной группе после 5- дневного возраста, установили достоверное снижение бактерицидной активности. Это различие составляет в сравнении с 10- дневным возрастом 9,27 %. Во второй подопытной группе гибель телят отмечали в 5-6 дневном возрасте.
При изучении величины связи между бактерицидной и фагоцитарной активностью установили высокую степень корреляции (r=0,88; Р=0,02) между фагоцитарной и бактерицидной активностью у здоровых телят до 20-и дневного возраста, в 30-и дневном возрасте коэффициент корреляции был равен 0,66.
В 1-й подопытной группе (больные телята) величина связи между фагоцитарной и бактерицидной активностью у телят до 5-дневного возраста равнялась 0,81, а в 30-дневном возрасте коэффициент корреляции несколько снижался до 0,74, тем не менее, связь считается высокой. Во второй подопытной группе (больные-павшие телята) коэффициент корреляции равен r= 0,96, что указывает на наличие высокой отрицательной связи, с увеличением фагоцитарной активности (2-й день) уменьшается бактерицидная активность.
Таблица 3.16
Показатели гуморальной защитной реакции организма новорожденных телят (Р<0,05)
|
Возраст телят |
Бактерицидная активность (%) сыворотки крови телят(x±m) |
||
|
Здоровые, n=11 |
Больные, n=11 |
Павшие, n=7 |
|
|
До поения молозива |
21,82 ± 0,55 |
21,91 ± 0,58 |
21,86 ± 0,55 |
|
2-й день |
33,91 ± 0,44 |
28,00 ± 0,30 |
20,28 ± 0,52 |
|
5-й день |
35,91 ± 0,44 |
41,09 ± 0,23 |
20,00 ± 0,82 |
|
10-й день |
36,00 ± 0,25 |
31,82 ± 0,42 |
|
|
20-й день |
37,27 ± 0,27 |
34,18 ± 0,42 |
|
|
30-й день |
42,18 ± 0,42 |
36,73 ± 0,27 |
|

3.5. Комплементарная активность сыворотки крови новорожденных телят
Механизм действия комплемента заключается в повреждении структуры клеточной стенки микробной клетки, которая не распадается целиком, в ней образуются полости, отверстия через которые вытекает цитоплазма. Повреждение оболочки нарушает осмотическое давление, что также вызывает гибель клетки. Феномен лизирующего действия комплемента хорошо выражен в отношении грамотрицательных бактерий, но грамположительные бактерии, как правило, не разрушаются комплементом.
В микробиологической и клинической иммунологии широко используется гемолитический метод титрования комплемента и сравнительно реже определение содержания комплемента до его бактерицидной или цитотоксической активности и количественное определение в весовых единицах.
В основе принципа гемолитического метода определения комплемента лежит гемолиз сенсибилизированных гемолизином эритроцитов в присутствии комплемента. Количество комплемента в испытуемой сыворотке выражают в условных единицах гемолиза. Титр комплемента определяют по 50 % гемолиза [С. J. Howard, A. A. Glynn, 1971; И. М. Архангельский, 1976; О. Н. Грызлова, П. А. Емельяненко, В. Н. Денисенко, 1978].
В результате проведенных исследований установлено, что (табл. 3.20-3.23) комплементарная активность у новорожденных телят (контрольная группа здоровые телята) возрастает достоверно (Р<0,05) до 30-дневного возраста по сравнению с периодом до поеная молозивом. Так, активность комплемента на 2-й день, после выпойки молозива увеличилась в 1,35 раза, на 5-й день в 1,38 раза, на 10-й день в 1,46 раза и на 30-й день в 1,67 раза, что соответствует до выпойки молозива 153,75±0,55, на 2-й день 208,08±0,42, на 5-й день 21З,00±0,68, на 10-й день 224,67±0,47 и на 30-й день 256,83±0,60 ед/мл.
В первой подопытной группе (больные телята) также имеет место тенденция увеличения активности комплемента в связи с возрастным периодом. Так, на 2-й, 5-й, 10-й день это увеличение составило 1,19, 1,13 и 1,14 раза, что соответствует до выпойки молозива 153,50±66, на 2-й день 183,50±0,50, на 5-й день 173,50±0,61 и на 10-й день 774,50 ± 0,51. Достоверное увеличение комплементарной активности сыворотки крови отмечали на 20-й и 30-й дни по сравнению с 5-м и 10-м дневным периодом 184,67±0,38 и 194,50±0,44 по сравнению с 173,50±0,81 и 174,50±0,51 (Р<0,05).
Во второй подопытной группе (павшие телята) отмечали не достоверное повышение комплементарной активности. Так, активность комплемента до выпойки молозива, была равна 153,71±1,03 на 2-й и 5-й день, этот показатель соответствовал 169,43±0,57 и 170,00±0,82 (увеличение равнялось в 1,10 и 1,11 раза).
Следовательно, увеличение комплементарной активности в 1,0 и 1,11 раза не было адекватным значению комплемента, обеспечивающему сохранность новорожденных телят.

Таблица 3.20
Комплементарная активность сывороток крови телят в зависимости от клинического статуса (x±m)
|
Возраст телят |
Здоровые, n=12 |
Больные, n=12 |
Павшие, n=7 |
|
До поения молозивом |
153,75 ± 0,55 |
153,50 ± 0,66 |
153,71 ± 1,03 |
|
2-й день |
208,08 ± 0,42 |
183,50 ± 0,50 |
169,43 ± 0,57 |
|
5-й день |
213,00 ± 0,68 |
173,50 ± 0,61 |
170,00 ± 0,82 |
|
10-й день |
224,87 ± 0,47 |
174,50 ± 0,51 |
|
|
20-й день |
242,50 ± 0,47 |
184,64 ± 0,38 |
|
|
30-й день |
256.83 ± 0,60 |
194,50 ± 0,44 |
|
3.6. Лизоцимная активность сыворотки крови
Лизоцим по своей природе является ферментом (ацетилмурамидаза) и содержится почти во всех органах и тканях животных. Содержание его в сыворотке крови новорожденных телят коррелирует с бактерицидной активностью. Лизоцим стимулирует фагоцитоз нейтрофилов и макрофагов, синтез антител, а также способен разрушать липополизахаридные поверхностные слои клеточных стенок большинства бактерий. Снижение титра лизоцима. или исчезновение его в крови приводит к возникновению инфекционной болезни [X. Я. Грант, Л. И. Яворский, И. А. Блумберг, 1973; И. М. Архангельский, 1976; Ю. А. Ширванян, А. А. Акопов, 1985; С. И. Плященко, 1991].
На основании проведенных опытов на телятах (табл. 3.24-3.27) от рождения до 30-дневного возраста установлено, что новорожденный организм обладает достаточно выраженной лизоцимной активностью. Так, после выпойки молозива лизоцимная активность у здоровых телят (контрольной группы) увеличилась на 2-й день в 2,77 раза, на 5-й день в 2,82 раза и на 10-й день в 2,9 раза. На 30-й день лизоцимная активность в процентном отношении равнялась уровню лизоцимной активности сыворотки крови новорожденных телят 2-дневного возраста.
В первой подопытной группе (больные телята) лизоцимная активность увеличивалась после выпойки молозива, на 2-й день в 1,97 раза, на 5-й день в 1,41 раза и на 10-й день в 1,89 раза. На 30-й день лизоцимная активность существенно не отмечалась от уровня ее у телят 2-дневного возраста.
Во второй подопытной группе (павшие телята) лизоцимная активность сывороток крови после выпойки молозива, на второй день увеличилась в 1,3 раза и на 5-й день в 1,25 раза. Увеличение было существенно недостоверным.
Т
аблица
3.24
Лизоцимная активность крови телят в зависимости от клинического статуса (X±m)
|
Возраст телят |
Здоровые, n=10 |
Больные, n=10 |
Павшие, n=7 |
|
До поения молозива |
1,49±0,007 |
1,49±0,005 |
1,49±0,006 |
|
2-й день |
4,13±0,03 |
2,93±0,013 |
1,95±0,02 |
|
5-й день |
4,21±0,01 |
2,10±0,03 |
1,87±0,015 |
|
10-й день |
4,32±0,009 |
2,82±0,006 |
|
|
20-й день |
4,31±0,01 |
2,92±0,009 |
|
|
30-й день |
4,14±0,02 |
3,00±0,01 |
|
3.7. Пропердиновая активность сыворотки крови
Большую роль в естественной резистентности организма играет пропердин, который обладает разносторонним биологическим действием, так, например, в присутствии комплемента и ионов магния он способен разрушить сальмонеллы, эшерихии, протей, псевдомонасы, шигеллы, сенную палочку. Полная инактивация пропердина происходит при 58°С в течение 30 мин. За единицу действия пропердина принимают то количество, которая способно инактивировать третий компонент комплемента в 1 мл сыворотки, лишенной пропердина [ И. М. Архангельский, 1976; П. А. Емельяненко, О. И. Грызлова, Г. Н. Печникова и М. Н. Тулупова, 1980; С. И. Плященко, В. Т. Сидоров, А. Ф. Трофимов, 1990].
В результате проведенных исследований (табл. 3.28-3.31) на телятах от рождения до 30-дневного возраста установлено, что организм новорожденных телят обладает существенной пропердиновой активностью по сравнению с животными до выпойки молозива.
Так, на 2, 5-й день после выпойки молозива и на 10-й день после выпойки молока пропердиновая активность увеличилась соответственно в 1,41, 1,77 и в 1,71 раза. На 20 и 30-й дни существенного изменения в количественном содержании пропердина в сыворотки крови новорожденных телят по сравнению со 2-, 5- и 10-ым днем не установили.
В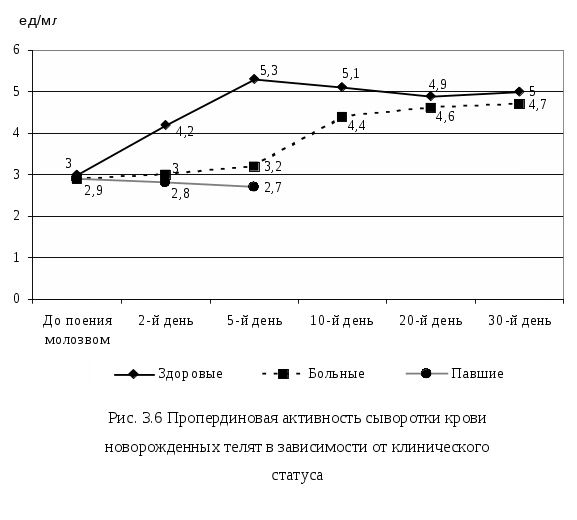
первой подопытной группе (больные
телята) пропердиновая активность после
выпойки молозива, на 2-й день не
изменялась(2,98±0,03 против 2,95±0,04),' на 5-й
день достоверного изменения, также не
устанавливали по сравнению с периодом
до поения молозива (3,2±0,44 против 2,95±0,04)
и только в 10-дневном возрасте пропердиновая
активность увеличилась в 1,5 раза и была
существенно достоверной (4,42±0,04 против
2,95±0,04).
Таблица 3.28
Количественное содержание пропердина в сыворотке крови телят в зависимости от клинического статуса (x±m)
|
Возраст телят |
Здоровые, n=10 |
Больные, n=10 |
Павшие, n=7 |
|
До поения молозива |
2,97±0,04 |
2,95±0,04 |
2,93±0,03 |
|
2-й день |
4,19±0,03 |
2,98±0,03 |
2,78±0,03 |
|
5-й день |
5,25±0,05 |
3,21±0,44 |
2,77±0,025 |
|
10-й день |
5,07±0,04 |
4,42±0,04 |
|
|
20-й день |
4,89±0,02 |
4,63±0,036 |
|
|
30-й день |
5,04±0,04 |
4,72±0,03 |
ВЫВОДЫ
Основной признак при колибактериозе телят - некомпенсированный метаболический ацидоз. Сопутствующими ацидозу были отклонения в уровнях сывороточных хлоридов, кальция, и калия.
Установлена связь между кислотно-щелочным равновесием и катаболизмом. Катаболизм отражает недостаточно функционирующие физиологические механизмы, воздействующие на причинный агент.
Новорожденные телята обладают защитно-приспособительной фагоцитарной реактивностью по отношению к Е. соli и эти бактерии поглощаются фагоцитами животного, еще не получавшего молозива. Уровень фагоцитарной активности и фагоцитарной интенсивности достоверно повышался у телят до 5-дневного возраста.
Бактерицидная активность сыворотки крови зависела от возраста телят и была наиболее низкой у новорожденных животных до приема молозива. Увеличение бактерицидной активности отмечали у здоровых телят в 3-5-дневном возрасте по сравнению с периодом до выпойки молозива.
Коэффициент корреляции между фагоцитарной и бактерицидной активностью у больных телят был равен 0,81, что указывает на высокую степень взаимосвязи между данными показателями.
Новорожденные животные обладают комплементарной, лизоцимной и пропердиновой активностью, еще до выпойки молозива, что указывает на их синтез самим организмом.
Комплементарная активность у новорожденных телят возрастала достоверно (P<0,05) до 30-дневного возраста по сравнению с периодом до выпойки молозива, особенно , в контрольной группе (здоровые), в то время как в группе больных увеличение было недостоверным (Р0,05).
Увеличение лизоцимной активности установили у новорожденных телят в 2-5 и 10 -дневном возрасте; увеличение исследуемого показателя в группе болъных было статистически недостоверным (Р>0,05).
Пропердиновая активность в сыворотке крови достоверно повышалась (Р>0,05) в контрольной группе животных в 2-5 и 10-дневном возрасте, в то время как в группе больные-павшие, содержание пропердина в сыворотке крови в аналогичный период существенно не изменялось (Р>0,05).
РАЗДЕЛ 4 ЭФФЕКТИВНОСТЬ ЛЕКАРСТВЕННЫХ ПРЕПАРАТОВ ПРИ КОЛИБАКТЕРИОЗЕ НОВОРОЖДЕННЫХ ТЕЛЯТ
4.1. Эффективность применения колостральной сыворотки и лактоглобулина для коррекции иммунодефицита неонатальных телят
Сыворотка молозива коров обладает высокой антибактериальной активностью и защищает организм новорожденного от внедрения бактерий, других чужеродных клеток и токсинов. К антибактериальным факторам сыворотки молозива относятся иммуноглобулины (антитела), которые имеют большое значение для невосприимчивости новорожденных телят к инфекционным болезням. Поскольку плацента коров непроницаема для антител крови,·новорожденные фактически лишены защиты от микроорганизмов. В постнатальный период они получают антитела в виде иммуноглобулинов молозива, которые в первые часы жизни в неселективном виде проходят через стенки их кишечника в кровь.
Колостральную сыворотку получали путем добавления 10 мл 1 %-ного раствора пепсина на 100 мл молозива с последующим выдерживанием смеси в термостате при 38° С в течение 4-6 ч. Сыворотку отделяли фильтрацией через (бумажный фильтр от взвешенных частиц, затем проводили фильтрацию через бактериальный фильтр Зейтца.
Для выделения лактоглобулинов к сыворотке молозива добавляли два или три объема дистиллированной воды в зависимости от содержания белка, затем в смесь при постоянном помешивании добавили тонкой струей насыщенный раствор сернокислого аммония до 38 %-ного насыщения. Выпавший осадок лактоглобулинов отделяли фильтрацией. Лактоглобулины освобождали от сернокислого аммония путем диализа. Впоследствии лактоглобулины разбавляли дистиллированной водой до получения 10 %-ного раствора белка, рН полученного раствора доводили до 7,2, 3 %-ным едким натрием и добавляли в раствор хлорид натрия до 0,85 %-ного насыщения.
После проверки на стерильность и безвредность лактоглобулин использовали с профилактической и лечебной целью.
Работа проведена в учебном хозяйстве «Кетросу» района Анений Ной. В данном хозяйстве анализировали условия содержания и кормления стельных коров и новорожденных телят, наблюдая клиническую картину и течение болезни, анализировали эффективность лечебных мероприятий, производили патологоанатомическое вскрытие и бактериологические исследования с последующей идентификацией выделенных культур.
Для патологоанатомических и бактериологических исследований использовали животных не подвергшихся лечению антибиотиками и химиотерапевтическими препаратам, через 1-4 ч после гибели, а также убитых с диагностической целью.
Высевы производили на МПА, МПБ и агар Эндо в чашках Петри из содержимого крови, сердца, селезенки, почек, печени, желчного пузыря, костного мозга, мезентеральных лимфатических узлов пораженных участков тонкого отдела кишечника.
Для идентификации снимали с агара Эндо по 3 одинаковые, не отличающиеся по цвету и морфологии, колонии.
Видовую принадлежность выделенных культур устанавливали путем изучения культурально-морфологических и тинкториальных свойств, ферментативной активности, а также по патогенности для белых мышей и серологической типизации.
Анализ показал, что в обследованном хозяйстве имеет место нарушения санитарно-гигиенических и ветеринарных правил содержания и кормления коров и новорожденных телят. В хозяйстве в первые 2-3 дня жизни заболевали почти все новорожденные животные с признаками диареи. Летальность варьировала до 50 % и более. Гибель новорожденных наступала в отдельных случаях в первые сутки жизни, но в большинстве случаев в течении первых десяти суток. Клиническая картина болезни проявлялась угнетением, вялостью, пониженным аппетитом, диареей.
Патологоанатомическому и бактериологическому исследованию подвергнуто 5 трупов и фецес больного теленка, У 4 трупов картина патологоанатомического вскрытия соответствовала специфической форме колибактериоза, а в одном случае изменения характеризовались поражением желудочно-кишечного тракта и мезентеральных лимфатических узлов.
Из органов 5 трупов и фецеса больного теленка выделена 21 культура. Вирулентность выделенных культур была различной и она варьировала в пределах одного трупа и даже одного органа.
Из большинства паренхиматозных органов, или мезентеральных лимфатических узлов и пораженных участков тонкого участка кишечника выделена чистая культура Е. coli энтеропатогенных серогрупп (О78:К80; О119:К69; О137:К79). Серологическую типизацию по О-антигену проводили при помощи О-диагностических сывороток, приготовленных Армавирской биофабрикой. Определение К-антигена проводили при помощи ОК-сывороток Московского института вакцин и сывороток имени И. И. Мечникова.
Для коррекции иммуноглобулинового дефицита (меньше 20 мг/мл по цинк-сульфатному тесту) вводили внутривенно по 50 мл лактоглобулина или по 100 мл колостральной сыворотки через 24-36 ч после первой выпойки молозива (не позже 2 ч после рождения) телятам.
С целью профилактики и лечения больных животных колостральную сыворотку и лактоглобулины применяли телятам 1-2 -дневного возраста. Одновременно исключали одну выпойку молозива, заменяя ее подогретым до 37-38° С изотоническим раствором хлорида натрия в количестве 1000 мл и вводили внутримышечно или подкожно по 0,5 мл лактоглобулина или по 1 мл колостральной сыворотки на 1 кг живой массы теленка. Препараты перед применением подогревали на водяной бане до 37-38°С.
При последующем кормлении телятам однократно вводили по 300-350 мл молозива разбавленного изотоническим раствором хлорида натрия (800-1000мл), постепенно увеличивая в каждое последующее кормление количество молозива на 150-200 мл до полной суточной дозы.
Препараты инъецировали повторно через 24 ч. Лечение телят продолжали до полного исчезновения клинических признаков заболевания (улучшение общего состояния, восстановления приема молозива др.). Выздоровление телят, как правило, наступало после 2-3 разового применения колостральной сыворотки или лактоглобулина. Препараты не оказывали побочного действия и не вызывали осложнений. Противопоказаний к их применению нет.
Наряду с применением колостральной сыворотки и лактоглобулина, в хозяйстве проводили комплекс ветеринарно-санитарных мероприятий, предусмотренных при выращивании телят молозивного периода [B. C. Шипилов, В. П. Шишков, В. Г. Зароза, В. П. Карев, Г. Д. Смоленская, 1987].
Проведение указанных мероприятий приводило, как правило, к ликвидации заболевания.
Наставление по применению колостральной сыворотки и лактоглобулина с цепью профилактики колибактериоза телят одобрено научно-техническим советом Министерства сельского хозяйства и продовольствия Республики Молдова (протокол №3 от 3.Х.91 г.).
4.2. Эффективность лактоглобулина и ретинола применяемых с целью коррекции иммунодефицита новорожденных телят.
Лечение новорожденных телят при иммунодефицитном состоянии без применения иммунокорректирующих препаратов очень затруднительно в связи с тем, что телята рождаются гипогаммаглобулинемичными, крайне важна быстрая абсорбция колостральных иммунных лактоглобулинов, что связано с своевременным поглощением молозива. Эффективность абсорбции иммунных лактоглобулинов связано с количеством проглоченного молозива, возраста телят и влияния присутствия коров-матерей в течение первых 24 ч жизни.
Для идентификации иммунодефицита телят применяют простой, но точный тест — цинк-сульфатный.
Известны способы лечения колибактериоза у телят, основанные на использовании препаратов, содержащих антибиотики, сульфаниламиды и нитрофураны.
Однако они обладают целым рядом недостатков. Прежде всего они характеризуются весьма относительной лечебной эффективностью. Микроорганизмы быстро адаптируются к антибиотикам, становятся полирезистентными. Тестирование эффективности терапевтических препаратов против колибактериоза у телят зависит от определения концентрации сывороточных иммуноглобулинов, так как при низкой концентрации лечение, как правило, не эффективно.
Наиболее близким предлагаемому является способ лечения колибактериоза у телят с использованием лактоглобулина путем выпаивания в дозе 150-200 мл [R. Aschaffenburg, S. Bartlett, S. K. Kon, J. M. Roy and D. M. Walker, 1951a]. Однако данный способ не обеспечивает достижения терапевтического эффекта.
Задачей является повышение эффективности лечения за счет коррекции иммунодефицита и поддержания нормального состояния эпителия слизистых оболочек пищеварительного тракта новорожденных телят.
Это достигается тем, что в способе лечения телят от колибактериоза, предусматривающем использование лактоглобулина дополнительно вводят ретинол (ретинола ацетат) внутримышечно в дозе 100000-150000 МЕ 1 раз в сутки в течение 2-3 суток, причем лактоглобулин вводят внутривенно в дозе 75-80 мл 1 раз в сутки в течение 1-2 суток.
Совместное действие обоих препаратов обеспечивает высокую эффективность лечения. При этом повышается неспецифическая иммунологическая реактивность организма, способствующая подавлению жизнедеятельности возбудителя болезни. В свою очередь, исключение использования антибиотиков способствует сохранению эубактериоза организма и предотвращает развитие аллергических реакций у людей.
Способ осуществляется следующим образом. Вводят ретинол (ретинола ацетат) внутримышечно в дозе 100000-150000 МЕ 1 раз в сутки в течение 2-3 суток, причем лактоглобулин вводят внутривенно в дозе 75-80 мл 1 раз в сутки в течение 1-2 суток.
Пример осуществления способа. На молочной ферме учхоза «Кетросу» с поголовьем 220 коров заболели колибактериозом 64 теленка. Для лечения заболевших животных был использован предлагаемый способ, в соответствии с которым инъецировали внутривенно лактоглобулин в дозе 75-80 мл 1 раз сутки а течение 1-2 суток, а также вводили ретинол (ретинола ацетат) внутримышечно в дозе 100000-150000 МЕ 1 раз в сутки а течение 2-3 суток. В результате проведенного лечения выздоровело 62 теленка.
Для определения оптимальных параметров способа был поставлен ряд опытов, результаты которых сведены в таблицы 4.1-4.5.
Таблица 4.1.
Определение разовой дозы лактоглобулина
|
Группа |
Количество голов |
Доза в мл |
Кратность введения |
Выздоровело, % |
|
1 |
20 |
60 |
1 |
0 |
|
2 |
20 |
65 |
1 |
0 |
|
3 |
20 |
70 |
1 |
20 |
|
4 |
20 |
75 |
1 |
40 |
|
5 |
20 |
80 |
1 |
40 |
|
6 |
20 |
85 |
1 |
40 |
Как следует из таблицы, введение лактоглобулина в дозе менее 75 мл не эффективно или мало эффективно. При повышении дозы более 80 мл дальнейшего увеличения эффективности лечения не происходит. Таким образом, оптимальной является разовая доза внутривенного введения лактоглобулина в пределах 75-80 мл.
Таблица 4.2
Определение кратности введения лактоглобулина
|
Группа |
Количество голов |
Доза в мл |
Продолжительность курса |
Выздоровело, % |
|
1 |
20 |
75 |
1 |
75 |
|
2 |
20 |
75 |
2 |
80 |
|
3 |
20 |
75 |
3 |
80 |
|
4 |
20 |
75 |
4 |
80 |
Таким образом, оптимальным является введение лактоглобулина в течение 1-2 суток. Дальнейшее введение лактоглобулина не приводит к росту эффективности лечения.
Таблица 4.3
Определение оптимальной дозы ретинола в комплексе с лактоглобулином.
|
Группа |
Количество голов |
Доза лактоглобулина в мл |
Кратность введения |
Доза в МЕ |
Выздоровело, % |
|
1 |
20 |
75 |
2 |
50000 |
80 |
|
2 |
20 |
75 |
2 |
75000 |
80 |
|
3 |
20 |
75 |
2 |
100000 |
85 |
|
4 |
20 |
75 |
2 |
125000 |
85 |
|
5 |
20 |
75 |
2 |
150000 |
85 |
|
6 |
20 |
75 |
2 |
175000 |
85 |
Оптимальной является доза ретинола применяемого в комплексе с лактоглобулином в количестве 100000-150000 МЕ на однократное введение. Меньшее количество препарата не обеспечивает достаточно высокой эффективности лечения, а повышение дозы не приводит к увеличению процента выздоровления заболевших животных.
Таблица 4.4
Определение кратности введения ретинола.
|
Группа |
Количество голов |
Доза лактоглобулина в мл |
Кратность введения лактоглобулина |
Доза ретинола в МЕ |
Кратность введения ретинола |
Выздоровело, % |
|
1 |
20 |
75 |
2 |
125000 |
1 |
80 |
|
2 |
20 |
75 |
2 |
125000 |
2 |
95 |
|
3 |
20 |
75 |
2 |
125000 |
3 |
95 |
|
4 |
20 |
75 |
2 |
125000 |
4 |
95 |
Как следует из таблицы, ретинол целесообразно вводить 1 раз в сутки в течение 2-3 суток. Однократное его введение не обеспечивает достаточно высокой эффективности лечения. При продолжении введения ретинола более 3-х суток дальнейшего роста эффективности лечения не происходит. Таким образом, как следует из проведенных исследований, оптимальная терапевтическая эффективность лечения колибактериоза достигается при внутримышечном введении телятам ретинола в дозе 100000-150000 МЕ 1 раз в сутки в течение 2-3 суток в сочетании с внутривенным введением лактоглобулина в дозе 75-80 мл 1 раз в сутки в течение 1-2 суток. Такой способ лечения колибактериоза у телят обеспечивает достижение терапевтической эффективности порядка 95,0 %.
Таблица 4.5
Сравнительная терапевтическая эффективность препаратов.
|
Способ лечения |
Количество голов |
Доза препарата |
Кратность введения |
Выздоровело в % |
|
1. Предлагаемый комплекс состоит из: а) лактоглобулина; б) ретинола. |
20 |
75-80мл 100000-150000 МЕ |
1 раз в сутки в течение 1-2 суток |
95,0 |
|
2. Антибиотик (неомицин) |
20 |
5000 ЕД на 1 кг живой массы |
2 раза в сутки в течение 2-3 суток |
60,0 |
|
3. Прототип (лактоглобулин) |
20 |
150-200 мл |
Однократное выпаивание |
20,0 |
4.3. Применение хлорпромазина и Т-активина для лечения телят, больных колибактериозом
Лечение новорожденных телят очень затруднительно, в связи с резистентностью энтеротоксических штаммов Е. coli к большинству доступных химиотерапевтических средств.
Известны способы лечения колибактериоза у телят, основанные на использовании препаратов, содержащих антибиотики и сульфаниламиды.
Однако известные способы обладают целым рядом недостатков. Прежде всего, они характеризуются весьма относительной лечебной эффективностью часто наблюдаются рецидивы заболевания. Возбудитель колибактериоза быстро приобретает резистентность к большинству доступных химиотерапевтетеских средств.
Наиболее близким предлагаемому является способ лечения колибактериоза у новорожденных поросят с использованием препаратов производных фенотиазина, обладающих сильным антисекреторным действием и ограничивающих чрезмерную активность циклического АМФ.
Однако данный способ не обеспечивает достижения терапевтической эффективности.
Задачей является повышение эффективности лечения за счет увеличения бактерицидного действия на возбудителя колибактериоза и повышения неспецифической резистентности организма.
Это достигается тем, что в способе лечения телят от колибактериоза, предусматривающем применение хлорпромазина (аминазина) дополнительно вводят подкожно 0,01 %-ный раствор Т-активина в дозе 0,8-1,0 мл ежедневно в течение 2-3 суток, а аминазин вводят внутримышечно в форме 2,5 %-ного раствора в дозе 0,6-0,8 мл на 1 кг живой массы 1 раз в сутки в течение 2-3 суток,
Совместное введение обоих препаратов обеспечивает высокую эффективность лечения, предотвращающего рецидивы заболевания. При этом повышается неспецифическая резистентность организма, способствующая скорейшему ингибированию возбудителя болезни. Кроме того, исключение использования антибиотиков способствует сохранению эубиоза предотвращает развитие аллергии у людей.
Способ осуществлялся следующим образом. Вводят подкожно 0,01 % раствор Т-активина в дозе 0,8-1,0 мл ежедневно в течение 2-3 суток, а аминазин вводят внутримышечно в форме 2,5 %-ного раствора в дозе 0,6-0,8 мл на 1 кг живой массы 1 раз в сутки в течение 2-3 суток.
Пример осуществления способа. На молочной ферме учхоза «Кетросу» с поголовьем 220 голов заболели колибактериозом 62 новорожденных теленка. Для лечения заболевания животных был использован предлагаемый способ, в соответствии с которым вводили больным телятам аминазин 2,5 %-ной концентрации на 0,5-ном растворе новокаина внутримышечно в дозе 0,6-0,8 мл на 1 кг живой массы 1 раз в сутки в течение 3 суток. Одновременно с этим подкожно вводили 0,01 %-ный. раствор Т-активина в дозе 0,8-1,0 мл 1 раз в сутки в течение 3 суток. В результате проведенного лечения выздоровело 58 телят. Для определения оптимальных параметров способа был поставлен ряд опытов, результаты которых сведены в табл. 4.6-4.10.
Таблица 4.6
Определение разовой дозы аминазина
|
Группа |
Количество голов |
Доза в мл |
Продолжительность курса |
Выздоровело, % |
|
1 |
15 |
0,4 |
1 |
0 |
|
2 |
15 |
0,5 |
1 |
0 |
|
3 |
15 |
0,6 |
1 |
20 |
|
4 |
15 |
0,7 |
1 |
40 |
|
5 |
15 |
0,8 |
1 |
40 |
|
6 |
15 |
0,9 |
1 |
40 |
Как следует из таблицы, введение аминазина в дозе менее 0,6 мл неэффективно или малоэффективно при повышении дозы более 0,8 мл дальнейшего увеличения эффективности лечения не происходит. Таким образом, оптимальной является разовая доза внутримышечного введения 2,5 %-ного раствора аминазина в пределах 0,6-0,8 мл на 1 кг живой массы.
Таблица 4.7
Определение кратности введения Т-активина.
|
Группа |
Количество голов |
Доза аминазина |
Кратность введения аминазина |
Доза Т-активина |
Кратность введения Т-активина |
Выздоровело, % |
|
1 |
15 |
0,6 |
3 |
1,0 |
1 |
80 |
|
2 |
15 |
0,6 |
3 |
1,0 |
2 |
93,4 |
|
3 |
15 |
0,6 |
3 |
1,0 |
3 |
93,4 |
|
4 |
15 |
0,6 |
3 |
1,0 |
4 |
93,4 |
Как следует из табл. 4.7, Т-активин целесообразно вводить 1 раз в сутки в течение 2-3 суток. Однократное его введение не обеспечивает достаточно высокой эффективности лечения. При продолжении введения Т-активина более 3-х суток дальнейшего роста эффективности леченая не происходит.
Таблица 4.8
Определение кратности введения аминазина
|
Группа |
Количество голов |
Доза в мл |
Продолжительность курса лечения (сутки) |
Выздоровело, % |
|
1 |
15 |
0,6 |
1 |
0 |
|
2 |
15 |
0,6 |
2 |
0 |
|
3 |
15 |
0,6 |
3 |
20 |
|
4 |
15 |
0,6 |
4 |
40 |
Таким образом, при введении аминазина менее 2-х раз не достигается высокая эффективность лечения. Оптимальным является введение аминазина в течение 2-3 суток. Дальнейшее введение аминазина не приводит к росту эффективности лечения.
Таблица 4.9
Определение оптимальной дозы Т-активина в комплексе с аминазином
|
Группа |
Количество голов |
Доза аминазина |
Кратность введения |
Доза в мл |
Выздоровело % |
|
1 |
15 |
0,6 |
3 |
0,7 |
60 |
|
2 |
15 |
0,6 |
3 |
0,8 |
80 |
|
3 |
15 |
0,6 |
3 |
0,9 |
80 |
|
4 |
15 |
0,6 |
3 |
1,0 |
80 |
|
5 |
15 |
0,6 |
3 |
1,1 |
80 |
Оптимальной является доза Т-активина в количестве 0,8-1,0 мл на одно введение. Меньшее количество препарата не обеспечивало достаточно высокой эффективности лечения, а повышение дозы не приводит к увеличению процента выздоровления заболевших животных.
Таблица 4.10
Сравнительная терапевтическая эффективность препаратов
|
Способ лечения |
Количество голов |
Доза препарата |
Кратность введения |
Выздоровело % |
|
1. Предлагаемый комплекс состоит из: а) аминазина; б) Т-активина. |
15 |
0,6-0,8 мл 0,8-1 мл |
1 раз в сутки в течение 2-3 суток |
93,4 |
|
2. Антибиотик (неомицин) |
15 |
5000 ЕД на 1 кг живой массы |
2 раза в сутки в течение 2-3 суток |
40,0 |
|
3. Прототип (хлопромазин, син. аминазина) |
15 |
0,6 мл |
Однократное введение |
33,3 |
Таким образом, как следует из проведенных исследований, оптимальная терапевтическая эффективность лечения колибактериоза, достигается при внутримышечном введении аминазина в дозе 0,6-0,8 мл на 1 кг живой массы 1 раз в сутки и Т-активина в дозе 0,8-1,0 мл подкожно 1 раз в сутки в течение 2-3 суток. Такое осуществление способа лечения колибактериоза у телят обеспечивает достижение терапевтической эффективности порядка 93,4 %.
ВЫВОДЫ
Для коррекции иммунодефицита (уровень иммуноглобулинов меньше 20 мг/мл по цинк-сульфатному тесту) новорожденным телятам вводить внутривенно по 50 мл лактоглобулина или по 100 мл колостральной сыворотки.
Оптимальная терапевтическая эффективность лечения колибактериоза достигается при внутримышечном введении телятам ретинола в дозе 100000-150000 ME 1 раз в сутки в течение 2 - 3 суток в сочетании с внутривенным введением лактоглобулина в дозе 75-80 мл 1 раз в сутки в течение 1 - 2 суток.
Внутримышечное введение хлорпромазина в дозе 0,6-0,8 мл на 1 кг живой массы 1 раз в сутки и Т-активина в дозе 0,8-1,0 мл подкожно 1 раз в сутки в течение 2 - 3 суток обеспечивает достижение терапевтической эффективности порядка 93,4%.
5. Обсуждение результатов исследований.
Выполненная работа посвящена исследованию состоянию естественной резистентности и иммунологической реактивности у новорожденных телят при колибактериозе.
Постановка данного вопроса вызвана тем, что эта инфекция еще до сих пор сравнительно широко распространена в хозяйствах республики и наносит большой экономический ущерб, вследствие гибели телят, затрат труда и средств на лечение больных, а также за счет вынужденного выращивания слабого, недоразвитого после переболевания молодняка. Все это отрицательно сказывается на воспроизводствe стада, как одного из важнейших факторов роста поголовья и повышения продуктивности животных.
В комплексе задач по профилактике и лечению колибактериоза важное место занимает специфическая и неспецифическая иммунологическая реактивность, и ее коррекция посредством различных иммуномодуляторов и антисекреторных факторов.
Учитывая, что молозиво является единственным источником иммуноглобулинов, а, следовательно, и иммунологической защиты, т.е. специфической резистентности, необходимо создавать такие условия содержания и кормления стельных коров, чтобы добиться увеличения выхода молозива с высоким уровнем иммуноглобулинов и широкой противомикробной специфичностью. Основная часть иммуноглобулинов поступает в секрет молочной железы из крови в неизменном состоянии, аккумулируясь в молозиве за 3-9 дней до отела. Основным иммуноглобулином молозива коров является IgG. Иммуноглобулины классов М и А частично синтезируются плазматическими клетками молочной железы. По данным E. W. Fisher, A. A. Martinez, Z. Traianin, 1975; В. М. Чекишев, В. М. Васильев, А. И. Кабанцев, 1983; H. Balbierz, M. Nicolaijczuk, J. Zeilinski, 1983; Н. В. Матузенко, Е. В. Андреев, А. И. Собко, 1990; Д. Н. Масюк, 1997, уровень IgG в молозиве первого удоя составляет 73,4-122,2 мг/мл, IgM и IgA – соответственно 6,0 и 8,0 мг/мл.
По нашим данным, содержание иммуноглобулинов различных классов значительно варьирует, особенно заметно превалирует концентрация IgG в секрете молочной железы и в сыворотке крови по сравнению с IgМ и IgА. Количественное содержание IgG в молозиве составляло 49,5±0,21 мг/мл, тогда как концентрация IgМ и IgА равнялась соответственно 4,16±0,16 мг/мл и 3,93±0,28 мг/мл.
Содержание IgG в сыворотке крови равнялось 18,29±0,27 мг/мл, в то время как концентрация IgМ и IgА составляла соответственно 2,54±0,07 мг/мл и 0,49±0,02 мг/мл.
При изучении количественного содержания иммунных глобулинов в сыворотке молока установили значительное превышение IgG (0,59±0,03 мг/мл) в сопоставлении с уровнем IgМ (0,05±0,01 мг/мл) и IgА (0,13±0,01 мг/мл).
Процентное отношение содержания иммунных глобулинов в сыворотке крови и секрете молочной железы коров к общему уровню иммуноглобулинов составило соответственно в сыворотке крови IgG — 85,78 %; IgМ — 11,92 и IgА — 2,3 %. В сыворотке молозива эти показатели равнялись: IgG — 85,96 %; IgМ — 7,25 и IgА — 6,82 %.
Процентное отношение содержания иммунных глобулинов в сыворотке молока к общему уровню составило: IgG 76,62; IgМ — 6,50 и IgА — 16,88.
Установлено [Ю. Н. Федоров, 1996], что однократная дача молозива с содержанием 80 мг/мл телятям через 6, 12, 36 и 48 ч после рождения дает средние показатели иммуноглобулинов в сыворотке телят соответственно 66; 47; 12; 7 и 6 % от 100 % рассчитанного предложенного количества иммуноглобулинов.
Следовательно, чем раньше новорожденный теленок получит первое молозиво, тем более высокий уровень иммуноглобулинов поступает в кровоток.
Иммунологический статус, а, следовательно, своевременность и адекватность получения молозива у новорожденных телят контролировали в условиях хозяйства. С этой целью использовали цинк-сульфатный и натрий сульфитный тесты. Наиболее простым и информативным был пробирочный тест с сульфатом натрия. Данный метод давал возможность определять своевременность получения телятами молозива после рождения, принимать меры направленные на повышение иммунологического статуса. Это достигалось как дополнительной дачей молозива так и пероральным введением колостральной сыворотки и лактоглобулина, согласно методическим указаниям, утвержденным НТС МСХ и продовольствия Республики Молдова, Кишинев, 1994.
На значения молозива для новорожденных впервые указали Th. Smith and R. B. Little, 1922, которые предполагали, что телята приобретают защиту против белого поноса, получением из молозива материнских антител и демонстрировали защиту морских свинок протективными веществами молозива, против определенных штаммов Е. coli.
Однако, протективное действие молозива можно представить не как универсальное, а как лимитирующее и специфическое, так как определенные штаммы Е. coli против которых молозиво становится неэффективным, постепенно становятся доминантными.
При определении уровня иммуноглобулинов в сыворотке крови новорожденных телят установили, что концентрация колостральных иммунных глобулинов в крови телят до приема молозива составила 0,18±0,08 мг/мл. Этот уровень, главным образом, связан с IgG. Через 24 ч после выпойки молозива содержание иммуноглобулинов достигало 23,94±1,71 мг/мл или увеличилось в 133 раза.
Новорожденные телята с содержанием иммуноглобулинов 23,94±1,71 мг/мл (доверительный интервал 19,19-28,69 мг/мл) переболевали легкой формой диареи на третьи сутки. Даже такая высокая концентрация иммуноглобулинов не обеспечивает иммунологический статус новорожденных телят, что связано со значительной инфицированостью внешней среды и снижением вследствие этого защитного уровня молозива.
Содержание иммуноглобулинов через 24 ч в сыворотке крови телят сильно варьировало в составило от 19,85 до 29,11 мг/мл. Такая разница объясняется количеством проглоченного теленком молозива.
Способность новорожденных животных адаптироваться к изменениям внешней среды лимитировано и изменение условий, не влияющих на взрослых животных, может плохо отразиться на состоянии здоровья телят. Известно, что стресс является способствующим фактором возникновения диареи у телят. Причиной стресса могут быть различные условия внешней среды, порой самые безобидные и поэтому весьма трудно их предотвратить. Даже перемещение является причиной стресса, который приводит к увеличению уровня заражения инфекционными агентами. Низкая температура, сквозняки, влажная подстилка способствуют возникновению и распространению инфекции. Следовательно, иммунологический статус, необходимый для защиты организма зависит от количества молозива, проглоченного теленком в первые минуты жизни и условий внешней среды.
При концентрации иммуноглобулинов 15,37±0,19 мг/мг (доверительный интервал 14,82-15,90 мг/мл) у животных отмечали септическую форму диареи, депрессию, дегидратацию. Возникновение болезни характеризовалось профузным поносом, усиленной перистальтикой кишечника, запавшими глазами, телята не могли стоять и погибали в 7-10-дневном возрасте. Существенное уменьшение концентрации иммунных глобулинов ассоциировалось с тяжелой формой дегидратации и диареей.
Анализируя результаты абсорбции колостральных иммуноглобулинов новорожденными телятами, необходимо отметить, что всасывание иммуноглобулинов происходит в кишечнике и через лимфатические пути они попадают в кровь. Иммуноглобулины появляются в лимфе через 1-2 ч после поступления их в двенадцатиперстную кишку животных. Однако, факторы, обеспечивающие всасывание нерасщепленных молекул белка, как и причины, по которым оно становится невозможным, до настоящего времени изучены недостаточно.
Некоторые авторы [J. E. Butler, C. A. Kiddy, C. Maxwele, M. B. Hylton, A. Asofsky, 1971; J. E. Butler, C. Maxwele, 1972; P. Dobbelaar, I. P. T. M. Noordhuizen and Van K. A. S. Keulen, 1987] указывают на существовании специальных внутриклеточных рецепторов узнавания, обеспечивающих передачу белков в крови. Исследования механизмов транспорта и катаболизма иммуноглобулинов in vitro показывает, что на первых этапах всасывания формируется комплекс между иммуноглобулинами и специфическими рецепторами энтероцитов. Во взаимодействии с рецепторами клеток кишечника участвует Fc-фрагмент иммуноглобулинов. По-видимому, связывание иммуноглобулинов с клетками предохраняет их от катаболизма и способствует транспорту в крови.
Дальнейшее изучение колострального иммунитета новорожденных и выживаемости животных в неонатальный период связано с разработкой ряда вопросов выяснения механизма абсорбции иммуноглобулинов у новорожденных животных и роли иммуноглобулинов различных классов в создании колострального иммунитета; изучение клеточного колострального иммунитета; определения наиболее важных факторов, оказывающих влияние на содержание иммуноглобулинов в молозиве и сыворотке крови; унификация методов исследования содержания иммуноглобулинов с целью получения сопоставимых данных.
Изучая активность антител против К-антигена Е. coli в молозиве и в сыворотке крови телят 1-2 -дневного возраста, установили, что она была меньшей, чем активность против О-антигенов. Различная концентрация антител (титр) в сыворотке новорожденных телят против К- и О-антигенов Е. coli указывает, что кормление молозивом может стимулировать или подавлять синтез антител к определенным антигенам в одинаковой степени независимо от места их локализации в бактериальных клетках[B. Kaijser, S. Ahlstedt, 1977].
Иммуноглобулины в молозиво извлекаются селективной секрецией из сыворотки крови коров. Следовательно, специфичность антител в молозиве определяется специфичностью антител в сыворотке крови коров. Кроме того, уровень антител в сыворотке крови коров зависит от воздействия на иммунокомпетентные клетки специфических антигенов-иммуногенов, то есть активность антител молозива коров является рефлекторным отражением специфичности антигена. Вероятно, если определенные серотипы Е. coli имеются во внешней среде (коровник, родильное отделение), коровы впоследствии продуцируют специфические антитела и передают их через молозиво телятам. это имеет место в том случае, когда инвазионные и вирулентные серотипы Е. coli вызывают антигенную стимуляцию иммунокомпетентных клеток большую, чем не вирулентные штаммы.
Высший средний титр колостральных антител против К-антигена Е. coli был установлен против штамма Е. coli О137:К79. Идентификация антител в сыворотке молозива и в сыворотке крови телят против серотипов Е. coli О78:К80 и О119:К69 в более низких титрах и низшей частоте, может постулироваться их незначительной вирулентностью, инвазивнностью или тем, что антитела обладают перекрестной реакцией защиты и не специфичны для этих серотипов.
Результаты абсолютного теста сыворотки молозива и сыворотки крови телят указывают на перекрестную абсорбцию антител против Е. coli К- антигенов исследуемых серотипов. Перекрестная абсорбция была, особенно, очевидной с 2-типами В антигенов, так как абсорбция с 1 антигеном идентифицирует антитела против 2 антигенов (О78:К80 и О119:К69). Эти данные подтверждают значительную перекрестную реактивность антител к К-антигенам. Такая кросс-реактивность антител обеспечивает защиту неонатальных телят против инфекции с более чем 1 штаммом Е. coli. Следовательно, антитела с перекрестной активностью против К-антигенов Е. coli в естественных условиях способствует повышению резистентности неонатальных телят против различных штаммов.
Корреляционные таблицы биохимических показателей крови новорожденных телят указывает, что определенные параметры имеют очень сходные поведение (например, 1 и 4 бикарбонатная концентрация изменяется, непосредственно, в зависимости от рН крови, феномен хорошо известный). На другие параметры передается их обратная связь, как например, частичное соприкосновение с противоположной связью СО>2> и О>2>.
Кроме того, корреляционный анализ показывает на сопротивление между парциальным давлением О>2> в венозной крови и показателями кислотно-щелочного равновесия, в частности парциального давления СО>2> в порядке нейтрализации метаболического ацидоза в нормальном противодействии уменьшающим количества О>2> в тканях. Следовательно, снижение парциального давления СО>2> указывает на уменьшение количества О>2> в тканях.
Данные исследования показывают, что выжевшие телята имеют преимущество в количественном отношении изучаемых величин перед павшими животными.
Концентрация калия (мера катаболизма) имеет тенденцию к увеличению, в то время как концентрация натрия в крови уменьшается.
При тестировании естественной резистентности организма развивающихся животных принципиально важным является вопрос о подборе тест-микроба. Данные эпизоотологического анализа подтверждают, что во время внутриутробного развития инфекционную патологию у крупного рогатого скота вызывает преимущественно грамотрицательная микрофлора, поэтому для определения противомикробной активности факторов естественной резистентности (фагоцитоз, лизоцимная, бактерицидная активность), использовали в качестве тест культуры эшерихии. Так как, по значимости в патологии крупного рогатого скота, экологической пластичности, изученности свойств, доступности для поддержания и использования требованиям, предъявляемым к тест культурам, эшерихии являются наиболее подходящей моделью.
При тестировании фагоцитарной активности лейкоцитов телят подбирали соотношение тест-микроба и фагоцитирующих элементов, не допуская перегрузки последних, что особенно важно для новорожденных животных. Во избежание паралича фагоцитов, вызываемого патогенными факторами, в качестве тест микроба использовали культуру не вирулентного штамма.
При определении бактерицидной активности особое внимание уделяли подбору экспозиции контактирования тест- микроба с исследуемой пробой и тест штамма эшерихий, колонии, которых для полноценного выявления противомикробных свойств сыворотки должны быть в гладкой, а не в вариантных формах.
Лизоцимную активность определяли с учетом замедленной активности фермента крупного рогатого скота, тщательной подготовки материала к исследованию, что позволяло избегать ошибок, связанных с действием на тест-микробы других метрических факторов исследуемой пробы.
Тестирование комплементарной активности сыворотки крови было основано на подборе индикаторной системы, чувствительной к комплементу крупного рогатого скота, который не вызывает лизиса эритроцитов в классической гемолитической системе, вероятно, из-за лимитирующего содержания отдельных компонентов [П. А. Емельяненко, О. И. Грызлова, Г. Н. Печникова и М. Н. Тулупова, 1980].
Для определения пропердина использовали модифицированный метод, как более доступный из-за удешевления источника комплемента и уменьшения объема проб исследуемого материала [О. Н. Грызлова, П. А. Емельяненко, В. Н. Денисенко, 1978].
Анализируя показатели неспецифической иммунологической реактивности новорожденных телят, необходимо отметить, что уровень фагоцитарной активности и фагоцитарной интенсивности в крови телят в послемолозивный период статистически достоверно повышался до 5-дневного возраста в контрольной группе (здоровые) +7,55 и 0,38 (Р<0,001) и в первой опытной (больные) +2,89 и +0,23 (Р<0,001). Во второй опытной группе (больные-павшие) не отмечали повышения фагоцитарных показателей.
Значение бактерицидной активности зависит от возраста телят, наиболее низкий уровень отмечен у новорожденных до приема молозива (21,83±0,55 %). Достоверное повышение бактерицидной активности установлено у здоровых телят на 2-, 5-й дни жизни по сравнению с периодом до выпойки молозива. У больных телят отмечали на 10-й день достоверное снижение бактерицидной активности по сравнению с 5-дневным возрастом.
Коэффициент корреляции между фагоцитарной и бактерицидной активностью у больных телят равен 0,81, в 30-дневном возрасте этот показатель был равен 0,74, что указывает на высокую степень взаимосвязи.
Данные изучения комлементарной активности у новорожденных телят свидетельствуют о том, что этот показатель в контрольной группе (здоровые) животных возрастает достоверно (Р<0,05) до 30-дневного возраста по сравнению с периодом до поения молозивом. Так, активность комплемента у здоровых телят после выпойки молозива увеличилась на 2-, 5-, 10-й дни соответственно в 1,35, 1,38 и 1,46 раза, в то время как у больных в аналогичный период это увеличение составило 1,19 , 1,13 и 1,14 раза. У больных-павших животных на 2-, 5-й дни жизни увеличение равнялось в 1,10 и 1,11 раза.
Следовательно, увеличение комплементарной активности в 1,10 и 1,11 раза не было адекватным значению комплемента, обеспечивающему сохранность животных.
Лизоцимная активность у новорожденных телят после поения молозивом увеличилась у здоровых телят на 2-й день в 2,77 раза, на 5-й день в 2,52 раза и на 10-й в 2,9 раза; у больных животных это увеличение равнялось соответственно в 1,97, 1,41 и 1,89 раза. Увеличение лизоцимной активности в 1,3 раза на 2-й день и в 1,25 раза на 5-й день во второй опытной группе (больные-павшие) не предохраняло телят от гибели.
Анализируя показатели пропердиновой активности в сыворотке крови новорожденных животных после выпойки молозива, необходимо отметить, что уровень пропердина в сыворотке крови телят увеличился на 2-, 5- 10-й дни жизни соответственно в 1,41, 1,77 и в 1,71 раза, у больных телят увеличение пропердиновой активности отмечали в 1,5 раза только в 10-дневном возрасте, во второй опытной группе (больные-павшие) содержание пропердина в сыворотке крови на 2-й, 5-й дни после поения молозивом существенно не изменилось: 2,78±0,03 и 2,77±0,025 против 2,93±0,03 ед/мл до поения молозивом.
Данные изучения коррекции иммунодефицита неонатальных телят свидетельствует о том, что применение ретинола внутримышечно в дозе 100000-150000 МЕ 1 раз в сутки в течение 2-3 суток и лактоглобулина внутривенно в дозе 75-80 мл 1 раз в сутки в течение 1-2 суток, обеспечивает иммунокоррекцию порядка 95,0%, данные подтверждены патентом за №345, Республики Молдова от 31.12.1995.
Анализируя данные лечения телят, больных колибактериозом, необходимо отметить, что применение хлорпромазина внутримышечно в форме 2,5 %-ного раствора в дозе 0,6-0,8 мл на 1 кг живой массы 1 раз в сутки в течение 2-3 суток и Т-активина 0,01 %-ный раствор подкожно в дозе 0,8-1,0 мл ежедневно в течение 2-3 суток обеспечивает достижение терапевтической эффективности порядка 93,4 %. Результаты подтверждены патентом за №409, Республики Молдова от 29.02.1996.
Все это свидетельствует о перспективности использования в сельскохозяйственном производстве витамина А и лактоглобулина для коррекции иммунодефицита новорожденных телят, а для лечения колибактериоза телят-хпорпромазина и Т-активина.
6. ВЫВОДЫ.
Колибактериоз новорожденных телят в хозяйствах Республики Молдова занимает одно из ведущих мест. Протекает остро с признаками диареи, интоксикации, септицемии, расстройства сердечно-сосудистой и центральной нервной системы. В возникновении, развитии и исходе заболевания первостепенную роль играет уровень содержания иммноглобулинов в крови животных.
Концентрация иммунных глобулинов в сыворотке молозива коров первого, второго и третьего дня лактации составляет соответственно 57,02±0,86 мг/мл; 26,42±0,06 мг/мл и 11,56±0,42 мг/мл. Уровень содержания иммунных глобулинов в сыворотке крови новорожденных телят через 24 ч после рождения и в сыворотке молозива коров-матерей первого дня лактации составляет соответственно 23,096±1,00 мг/мл и 59,00±0,59 мг/мл (контрольная группа); 18,1±0,11 мг/мл и 57,02±0,86 мг/мл (энтеритная форма колибактериоза); 15,06±0,12 мг/мл и 54,53±0,57 мг/мл (септическая форма колибактериоза).
Количественная оценка абсорбции колостральных антител характеризует иммунный статус новорожденных телят и составляет соответственно 39,21±2,04 % (контрольная группа), 31,78±0,11 % (энтеритная форма колибактериоза) и 27,0±0,23 % (септическая форма колибактериоза). Установлена прямая зависимость между концентрацией иммуноглобулинов в сыворотке крови новорожденных животных и их резистентностью, что свидетельствует о прямой зависимости клинического статуса от сывороточной концентрации иммунных глобулинов и их катаболизма. Увеличение уровня иммуноглобулинов М и А в сыворотке крови клинически здоровых животных соответственно в 5,74 и 3,44 раза по сравнению с павшими указывает на их защитную функцию.
Концентрация IgG, IgМ и IgА в сыворотке крови коров составляет соответственно 18,24±0,27 мг/мл, 2,54±0,07 мг/мл и 0,49±0,02 мг/мл. Количественное содержание иммунных глобулинов в колостральной сыворотке существенно превалирует по сравнению с сывороткой крови и равняется IgG 49,5±0,21 мг/мл, IgМ 4,16±0,17 мг/мл, IgА 3,93±0,28 мг/мл. В сыворотке молока уровень иммунных глобулинов значительно меньший в сравнении с колостральной сывороткой и сывороткой крови коров, причем содержание IgА (0,13±0,01 мг/мл) в 2,6 раза больше, чем IgМ (0,05±0,01 мг/мл), что является отличительной особенностью перераспределения иммуноглобулинов в сыворотке крови и секрете молочной железы.
Содержание иммуноглобулинов в сыворотке крови и копрофильтратах у телят в зависимости от клинического статуса имеет существенное различие. Так, уровень катаболизма IgG, IgМ и IgА в группе больных телят составил соответственно 26,01; 60,00 и 51,69 %, а в группе павших телят это значение равнялось: 24,86; 12,91 и 69,77 %. Уровень катаболизма общих иммуноглобулинов в группе больных (выживших) телят был в 1,12 раза выше, чем в группе больных (павших), а IgМ в 4,64 раза.
Активность антител против К-антигенов Е. coli в молозиве и в сыворотке крови телят 1-2 дневного возраста была низшей, чем активность против О антигенов. Титр К-антител в сыворотке крови неонатальных телят зависит от концентрации К антител в молозиве коров. Различная концентрация антител (титр) в сыворотке крови новорожденных телят против К и О антигенов Е. coli указывает, что кормление молозивом может стимулировать или ингибировать синтез антител к определенным микроорганизмам в одинаковой степени независимо от места их локализации в бактериальных клетках.
«К» антитела-агглютинины молозива коров являются фактором, защищающим белых мышей против экспериментального колибактериоза, эффективность проб молозива против определенных серотипов Е. coli можно тестировать агглютинационным методом. Титр О-антител в сыворотке молозива коров и в сыворотке крови телят равнялся соответственно: 1:20-1:160 и 1:10-1:40. При серологической типизации изолированных штаммов Е. coli по К-антигену, обнаруживали фимбрии К99 и К88ас. Большинство тестированных сывороток крови телят не содержали К-антитела.
Наиболее важным фактором, влияющим на выращивание новорожденных телят является абсорбция колостральных антител в соответствующем физиологическом количестве. Иммунизация стельных коров поливалентной гидроокисиалюминиевой формол-тиомерсаловой вакциной против колибактериоза (эшерихиоза) телят не ингибирует серотипы Е. coli О8 и О101, входящие в состав вакцины и выделенные от павших телят.
При статистической обработке результатов исследования с применением ЭВМ «НАИРИ» крови новорожденных телят, установлено, что использование трех показателей легко измеряемых величин (мочевина, гематокрит, хлориды) позволяет прогнозировать заболевание и, следовательно, профилактировать с применением регидратационной терапии, возмещением кислотности и потери ионов натрия.
Уровень фагоцитарной активности и фагоцитарной интенсивности в крови телят в послемолочный период статистически достоверно повышался до 5-дневного возраста в контрольной группе (здоровые) соответственно +7,55 и +0,33 (Р<0,001) и в первой опытной (больные) +2,89 и +0,23 (Р<0,001). Во второй опытной группе (павшие) не отмечали повышение фагоцитарных показателей.
Бактерицидная активность сыворотки крови зависит от возраста телят, наиболее низкий уровень отмечен у новорожденных до приема молозива (21,23±0,55 %). Повышение бактерицидной активности установлено у здоровых телят на 2-, 5-й дни жизни по сравнению с периодом до выпойки молозива. На 10-й день у больных телят отмечали достоверное снижение бактерицидной активности по сравнению с 5-дневным возрастом. Коэффициент корреляции между фагоцитарной и бактерицидной активностью у больных телят равен 0,81, что указывает на высокую степень взаимосвязи.
Комплементарная активность у новорожденных телят (здоровых) возрастает достоверно (Р<0,05) до 30-дневного возраста по сравнению с периодом до выпойки молозива. Так, активность комплемента у здоровых телят после выпойки молозива увеличилась на 2-, 5-, 10-й дни соответственно в 1,35, 1,38 и 1,46. В то время как у больных в аналогичный период это увеличение составило 1,19, 1,13 и 1,14 раза. У павших животных на 2-, 5-й дни жизни увеличение равнялось 1,10 и 1,11 раза, что не обеспечивало сохранность животных.
Новорожденные телята обладают достаточно выраженной лизоцимной активностью, которая после выпойки молозива увеличилась у здоровых телят на 2-й день в 2,77 раза, на 5-й день в 2,82 раза и на 10-й день в 2,9 раза, у больных это увеличение соответственно равняется 1,97, 1,41 и 1,89 раза. Увеличение лизоцимной активности в 1,3 раза на 2-й день и в 1,35 раза на 5-й день во второй опытной группе (больные-павшие) не предохраняло телят от гибели.
Увеличение пропердиновой активности в сыворотке крови новорожденных животных после выпойки молозива установили на 2-й, 5-й и 10-й дни соответственно в 1,41, 1,77 и в 1,71 раза, у больных телят увеличение пропердиновой активности отмечали в 1,5 раза только в 10-дневном возрасте, во второй опытной группе (больные-павшие) содержание пропердина в сыворотке крови на 2-й, 5 й дни после поения молозивом существенно не изменялось 2,78±0,03 и 2,77±0,025 против 2,93±0,03 до поения молозивом.
Применение ретинола внутримышечно в дозе 100000-150000 МЕ 1 раз в сутки в течение 2-3 суток и лактоглобулина внутривенно в дозе 75-80 мл 1 раз в сутки в течение 1-2 суток обеспечивает иммунокоррекцию до 95,0 %.
Применение хлорпромазина внутримышечно в форме 2,5%-ного раствора в дозе 0,6-0,8 мл на 1 кг живой массы 1 раз в сутки в течение 2-3 суток и 0,01%-ного раствора Т активина подкожно в дозе 0,8-1,0 мл ежедневно в течение 2-3 суток обеспечивает терапевтическую эффективность до 93,4 %.
7. РЕКОМЕНДАЦИИ ПРОИЗВОДСТВУ.
Для обеспечения максимальной сохранности здоровых, жизнеспособных телят, рекомендуется применение колостральной сыворотки и лактоглобулина для коррекции иммунодефицита неонатальных телят (Методические рекомендации, одобренные НТС МСХ и продовольствия Республики Молдова, Кишинев, 1992).
Для выявления «К» антител-агглютининов против К-антигенов Е. coli рекомендуется экспресс-индикация их уровня в колостральной сыворотке коров и сыворотке крови новорожденных телят (Методические рекомендации, одобренные НТС МСХ и продовольствия Республики Молдова, Кишинев, 1994).
Для коррекции иммунодефицита новорожденных телят рекомендуется применение ретинола и лактоглобулина (Патент №345 от 31.12.1995, Республики Молдова).
Кислотно-основной баланс рекомендуется изучать по трем основным показателям: гематокриту, мочевине, хлоридам.
Для лечения больных телят колибактериозом рекомендуется использовать хлорпромазин и Т-активин (Патент №409 от 29.02.1996, Республики Молдова).
Результаты экспериментальных исследований, изложенные в настоящей диссертации, целесообразно включить в программу повышения квалификации ветеринарных и зооинженерных специалистов и в раздел «Инфекция и иммунитет» для студентов ветеринарных и зооинженерных факультетов аграрных университетов и колледжей.