Цитология, эмбриология, общая гистология
Луганский национальный аграрный университет
Цитология, эмбриология, общая гистология
(курс лекций)
Луганск - 2005
Цитология, эмбриология, общая гистология
Курс лекций составлен заведующим кафедрой биологии животных, доктором биологических наук, профессором Г.Д. Кацы.
Издание 2-е, переработанное и дополненное.
Лекции подготовлены для студентов зообиотехнологического и факультета ветеринарной медицины Луганского национального аграрного университета. Искренне благодарю аспиранта кафедры биологии животных Крыцю Я.П. и заведующую лабораторией Есауленко В.П. за помощь при подготовке материала к изданию.
Введение в гистологию
1. Предмет гистологии и её место в системе биологических и ветеринарных наук.
2. История и методы микроскопических исследований.
3. Клеточная теория, основные положения.
1. Специфика сельскохозяйственного производства обусловлена тем: что несмотря на возрастание роли технических факторов: главными орудиями и средствами производства остаются биологические объекты. По охвату объектов изучения и по своей глубине ветеринария представляет: как говорил академик К.И.Скрябин, интереснейшую область человеческого знания: в которой исследуется и охраняется такое множество представителей животного царства.
Цитология, гистология и эмбриология, наряду с физиологией, биохимией и другими науками формирует фундамент современной ветеринарии.
Гистология (греч. histos-ткань, logos-учение)- наука о развитии, строении и жизнедеятельности тканей животных организмов. Современная гистология изучает структуры организма животных и человека в связи с происходящими в них процессами, раскрывает соотношения между функцией и структурой и т.д.
Гистологию делят на 3 основных раздела: цитологию, или учение о клетке; эмбриологию, или учение о зародыше и гистологию общую и частную, или учение о тканях, о микроскопическом строении органов, их клеточном и тканевом составе.
Гистология тесно связана с рядом биологических и ветеринарных наук - общей и сравнительной анатомией, физиологией, патологической физиологией и патологической анатомией, а также некоторыми клиническими дисциплинами (внутренние болезни, акушерство и гинекология и др.).
Будущим врачам необходимо хорошее знание строения клеток и тканей органов, являющихся структурной основой всех видов жизнедеятельности организма. Значимость гистологии, цитологии и эмбриологии для врачей возрастает ещё потому, что для современной ветеринарной медицины характерно широкое применение цитологических и гистологических методов при проведении анализов крови, костного мозга, биопсии органов и пр.
2. Понятие ткань впервые было введено в биологию блестящим молодым французским ученым анатомом и физиологом Ксавье Биша (Bichat, 1771-1802), на которого произвела такое сильное впечатление разнообразная текстура различных слоев и структур, обнаруженных им при анатомических исследованиях, что он написал книгу о тканях организма, дав в ней название более чем 20 их видам.
Термин “”гистология” не принадлежит Биша, хотя его и можно считать первым гистологом. Термин “гистология” через 17 лет после смерти Биша предложил немецкий исследователь Мейер.
Ткань есть филогенетически обусловленная элементарная система, объединенная общей стуктурой, функцией и развитием (А.А. Заварзин).
Успехи гистологии с момента зарождения и по настоящее время прежде всего связаны с развитием техники, оптики и методов микроскопирования. Историю гистологии можно разделить на три периода: 1-й - домикроскопический (продолжительность около 2000 лет), 2-й - микроскопический (около 300 лет), 3-й - электронно-микроскопический (около 40 лет).
В современной гистологии, цитологии и эмбриологии применяются разнообразные методы исследования, позволяющие всесторонне изучать процессы развития, строения и функции клеток, тканей и органов.
Объектами исследования служат живые и мертвые (фиксированные) клетки и ткани, их изображения, полученные в световых и электронных микроскопах или на телевизионном экране. Существует ряд методов, позволяющих проводить анализ указанных объектов:
1) методы исследования живых клеток и тканей: а) прижизненное исследование клеток в организме (in vivo) - с помощью методов вживления прозрачных камер в организм животных, методом трансплантации;
б) исследование живых структур в культуре клеток и тканей (in vitro) - недостатки: утрачивается взаимосвязь с другими клетками и тканями, действие комплекса нейрогуморальных факторов регуляции и другое;
в) витальное и суправитальное окрашивание, то есть прижизненное окрашивание и окрашивание живых клеток, выделенных из организма.
2) исследование мертвых клеток и тканей; основным объектом исследования здесь являются гистологические препараты, приготовляемые из фиксированных структур.
Процесс изготовления гистопрепарата для световой и электронной микроскопии включает следующие основные этапы: 1) взятие материала и его фиксация, 2) уплотнение материала, 3) приготовление срезов, 4) окрашивание или контрастирование цвета. Для световой микроскопии необходим ешё один этап - заключение срезов в бальзам или другие прозрачные среды (5).
3) исследование химического состава и метаболизма клеток и тканей:
- цито- и гистохимические методы,
- метод радиоавтографии, в основе которого лежит использование радиоактивных элементов (например, фосфора-32Р, углерода -14С, серы-35S, водорода-3Н) или меченных им соединений.
- метод дифференциального центрифугирования - метод основан на применении центрифуг, дающих от 20 до 150 тыс. оборотов в минуту. При этом отделяются и осаждаются различные компоненты клеток и определяется их химический состав. - интерферометрия - метод позволяет оценить сухую массу и концентрацию плотных веществ в живой и фиксированной клетках. - количественные гистохимические методы - цитоспектрофотометрия - метод количественного изучения внутриклеточных веществ по их абсорбционным свойствам. Цитоспектрофлюориметрия - метод изучения внутриклеточных веществ по спектрам их флюорисценции.
4) методы иммунофлюорисцентного анализа. Они применяются для изучения процессов дифференцировки клеток, выявления в них специфических химических соединений и структур. Они основаны на реакциях антиген-антитело.
Методы микроскопирования гистологических препаратов:
- световая микроскопия: а) ультрафиолетовая, б) флюоресцентная (люминисцентная).
- электронная микроскопия: а) просвечивающая, б) сканирование (считывание). Первая дает лишь плоскостное изображение, вторая - пространственное; главным достоинством последнего (растрового) является большая глубина резкости (в 100-1000 раз больше, чем у световых микроскопов), широкий диапазон непрерывного изменения увеличения (от десятков до десятков тысяч раз) и высокая разрешающая способность.
3. Организм высших животных состоит из микроскопических элементов - клеток и ряда их производных - волокон, аморфного вещества.
Значение клетки в многоклеточном организме определяется тем, что через неё передается наследственная информация, с неё начинается развитие многоклеточных животных; благодаря деятельности клеток образуются неклеточные структуры и основное вещество, которые вместе с клетками образуют ткани и органы, выполняющие специфические функции в сложном организме. Создателем клеточной теории следует считать Дютроше (1824, 1837) и Шванна (1839).
Дютроше (1776-1847) - зоолог, ботаник, морфолог, физиолог. В 1824 г. он опубликовал свою книгу “”Анатомические и физиологические исследования о тонком строении животных и растений, а также о их подвижности”.
Созданию клеточной теории предшествовали следующие открытия. В 1610 году 46-летний проф. матетатики Падуанского университета Г.Галилей сконструировал микроскоп. В 1665 г. Роберт Гук открыл клетку при увеличении 100 х. Его современник, Феличе Фонтана говорил: “”...Посмотреть в микроскоп может каждый, но лишь немногие могут судить о виденном”. “”Микрография” Гука включала 54 наблюдения, в т.ч.”Наблюдение 18. О схематизме или строении пробки или о клетках и порах в некоторых других рыхлых телах”.
Из истории. Компания живших в Лондоне молодых людей (студентов) в 1645 г. стала собираться каждый день после занятий, чтобы обсуждать проблемы экспериментальной философии. Среди них были Роберт Бойль (18 лет), Р.Гук (17 лет), Рэн (23 года) и др. Так зародилась Британская академия, затем Лондонское Королевское общество (Карл II был её почетным членом).
Животную клетку открыл Антон ван Левенгук (1673-1695). Жил он в Делфте и торговал сукном. Свои микроскопы довел до 275 х. Петру I показывал кровообращение в хвосте личинки угря.
В настоящее время клеточная теория гласит: 1) клетка является наименьшей единицей живого, 2) клетки разных организмов сходны по своему строению, 3) размножение клеток происходит путем деления исходной клетки, 4) многоклеточные организмы представляют собой сложные ансамбли клеток и их производных, объединенные в целостные интегрированные системы тканей и органов, подчиненные и связанные между собой межклеточными, гуморальными и нервными формами регуляции.
Клетка - элементарная единица живого
1. Состав и физико-химические свойства живого вещества.
2. Типы клеток. Теории происхождения эукариотической клетки.
3. Клеточные мембраны, их молекулярный состав и функции.
1. Типичную клетку с ядром, цитоплазмой и всеми содержащимися в ней органеллами еще нельзя считать наименьшей единицей живого вещества, или протоплазмы (греч. “протос” -первый, “плазма” -образование). Существуют и более примитивные или более просто организованные единицы жизни - так называемые прокариотические организмы (греч. “карион” - ядро), к которым относится большинство вирусов, бактерии и некоторые водоросли; у них в отличие от клеток высшего типа с настоящим ядром (эукариотические клетки) отсутствует ядерная оболочка и ядерное вещество смешивается или непосредственно соприкасается с остальной протоплазмой.
В состав живого вещества входят белки, нуклеиновые кислоты (ДНК и РНК), полисахариды и липиды. Химические компоненты клетки можно разделить на неорганические (вода и минеральные соли) и органические (белки, углеводы, нуклеиновые кислоты, липиды и т.д.).
Цитоплазма растительной и животной клетки содержит 75-85 % воды, 10-20 % белка, 2-3 % липидов, 1 % углеводов и 1 % неорганических веществ.
ДНК - это молекула (её содержится 0,4 %), которая содержит генетическую информацию, направляющую синтез специфических клеточных белков. На одну молекулу ДНК приходится около 44 молекул РНК, 700 молекул белка и 7000 молекул липидов.
Первичная структура РНК подобна структуре ДНК, за исключением того, что РНК содержит рибозу и вместо тимина урацил. В настоящее время установлено, что существуют различающиеся молекулярным весом и другими свойствами три типа РНК: рибосомная, информационная и транспортная. Эти три типа РНК синтезируются в ядре и участвуют в синтезе белка.
2. Шаттон (1925) разделил все живые организмы на два типа (клистера) - прокариоты и эукариоты. Они дивергировали в докембрии (600-4500 млн. лет назад). Существуют две концепции происхождения эукариотической клетки: экзогенная (симбиотическая) и эндогенная. Первая основана на признании принципа объединения разных прокариотных организмов друг с другом. Эндогенная концепция основана на принципе прямой филиации, т.е. последовательного эволюционного преобразования прокариотных организмов в эукариотные.
В организме млекопитающих гистологи насчитывают около 150 типов клеток, и большинство из них приспособлено к выполнению какой-то одной определенной задачи. Форма и строение клетки зависят от выполняемой ею функции.
Функции клеток: раздражимость, сократимость, секреция, дыхание, проводимость, поглощение и усвоение, экскреция, рост и размножение.
3. Любую клетку отграничивает плазматическая мембрана. Она настолько тонка, что её невозможно различить под световым микроскопом. Плазматическая мембрана, легко поврежденная микроиглой, способна к восстановлению, но при более грубом повреждении, особенно в отсутствие ионов кальция, цитоплазма вытекает через прокол наружу и клетка погибает.
Согласно современной теории, плазматическая мембрана состоит из бислоя полярных липидов и встроенными в него молекулами глобулярных белков. Благодаря этим слоям мембрана, обладает эластичностью и относительной механической прочностью. Плазматическая мембрана большинства типов клеток состоит из трёх слоёв шириной примерно 2,5 нм каждый. Подобная структура, называемая “элементарной мембраной”, обнаружена и в большинстве внутриклеточных мембран. Биохимический анализ показал, что липиды и белки содержаться в них в отношении 1.0 : 1.7. Белковый компонент, названный строматином, представляет собой кислый фибриллярный белок с высоким молекулярным весом. Основную массу липидных компонентов образуют фосфолипиды, преимущественно лецитин и кефалин.
Плазмолемма - оболочка клетки, выполняющая отграничительную, транспортную и рецепторную функции. Она обеспечивает механическую связь клеток и межклеточные взаимодействия, содержит клеточные рецепторы гормонов и других сигналов окружающих клетку среды, осуществляет транспорт веществ в клетку из клетки как по градиенту концентраций - пассивный перенос, так и с затратами энергии против градиента концентраций - активный перенос.
В состав оболочки входят плазматическая мембрана, немембранный комплекс - гликокалекс и субмембранный опорно-сократительный аппарат.
В гликокалексе содержится около 1 % углеводов, молекулы которых образуют длинные ветвящиеся цепи полисахаридов, связанные с белками мембраны. Находящиеся в гликокалексе белки - ферменты участвуют в конечном внеклеточном расщеплении веществ. Продукты этих реакций в виде мономеров поступают в клетку. При активном переносе транспорт веществ в клетку осуществляется или поступлением молекул в виде раствора - пиноцитоз, или захватом крупных частиц - фагоцитоз.
В соответствии с функциональными и морфологическими особенностями тканей оболочка клеток образует характерные для них аппараты межклеточных контактов. Основные их формы: простой контакт (или зона слипания), плотный (замыкающий) и щелевой контакт. Разновидностью плотного контакта являются десмосомы.
Биологичекие мембраны действуют как диффузные барьеры. Благодаря своей избирательной проницаемости для ионов К+, Nа+, Cl- и т.п., а также высокомолекулярных соединений они разграничивают внутри- и межклеточные зоны реакций и создают электрические градиенты и градиенты концентрации веществ. Это делает возможным существование упорядоченных биологических структур со специфическими функциями.
Проникновение веществ в клетку называется эндоцитозом. Но существует и экзоцитоз. Например, от аппарата Гольджи отшнуровываются секреторные пузырьки, мигрирующие по направлению к клеточной мембране и выбрасывающие наружу своё содержимое. При этом мембрана пузырька сливается с гомологичной ей клеточной мембраной.
На основании электронно-микроскопических данных можно предположить, что плазмолемма является продуктом аппарата Гольджи. От этой органеллы в виде непрерывно отделяющихся пузырьков постоянно идет транспорт мембранного материала (“”поток мембран”), восстанавливающего использованные участки плазмолеммы и обеспечивающего её рост после деления клетки.
Мембрана является носителем видоспецифических и специфических для клетки поверхностных свойств, связанных с характерным распределением на ней гликозаминогликанов и белков. Их молекулы могут также покрывать поверхность клеток в виде тончайших пленок и образовывать межклеточный матрикс между соседними клетками. Контактные свойства клеток и иммунные реакции определяются этими компонентами мембран.
У многих клеток, особенно у специализированных для всасывания (кишечный эпителий), на наружной стороне имеются волосовидные выросты - микроворсинки. Образуемая или “щеточная каемка” несет ферменты, принимает участие в расщеплении веществ и транспортных процессах. На базальной стороне клеток, специализированных на интенсивное пропускание жидкости (при осморегуляции), например, в эпителии почечных канальцев и мальпигиевых сосудов, мембрана образует множественные впячивания, составляющие базальный лабиринт. Продукт клеточной секреции, базальная мембрана, часто отграничивает эпителий от глубже лежащих клеточных слоёв.
Особые мембранные структуры возникают в местах соприкосновения соседних клеток. Там имеются области, где мембраны так тесно прилегают друг к другу, что не остаётся места для межклеточного вещества (плотный контакт). В других участках возникают сложные контактные органеллы - десмосомы. Они и другие контактные структуры служат для механического соединения и главное - обеспечивают химическую и электрическую интеграцию соседних клеток, облегчая межклеточный ионный транспорт благодаря своему низкому электрическому сопротивлению.
Строение животной клетки
1. Цитоплазма и органеллы, их функция.
2. Ядро, его строение и функции.
3. Типы деления, фазы клеточного цикла.
1. Цитоплазма, отделенная от окружающей среды плазмолеммой, включает в себя гиалоплазму, находящиеся в ней обязательные клеточные компоненты - органеллы, а также различные непостоянные структуры – включения (рис.1).
Гиалоплазма (hyalinos - прозрачный) - основная плазма, или матрикс цитоплазмы, представляет собой очень важную часть клетки, её истинную внутреннюю среду.
В электронном микроскопе матрикс имеет вид гомогенного и тонкозернистого вещества с низкой электронной плотностью. Гиалоплазма является сложной коллоидной системой, включающей в себя различные биополимеры: белки, нуклеиновые кислоты, полисахариды и др. Эта система способна переходить из золеобразного (жидкого) состояния в гелеобразное и обратно. В состав гиалоплазмы входят главным образом различные глобулярные белки. Они составляют 20-25% общего содержания белков в эукариотической клетке. К важнейшим ферментам гиалоплазмы относятся ферменты метаболизма сахаров, азотистых оснований, аминокислот, липидов и других важных соединений. В гиалоплазме располагаются ферменты активации аминокислот при синтезе белков, транспортные РНК (тРНК). В гиалоплазме при участии рибосом и полирибосом происходит синтез белков, необходимых для собственно клеточных нужд, для поддержания и обеспечения жизни данной клетки.
Органеллы - постоянно присутствующие и обязательные для всех клеток микроструктуры, выполняющие жизненно важные функции.
Различают мембранные органеллы - митохондрии, эндоплазматическую сеть (гранулярная и гладкая), аппарат Гольджи, лизосомы, к категории мембранных органелл относится и плазмолемма; н е м ембранные органеллы: свободные рибосомы и полисомы, микротрубочки, центриоли и филаменты (микрофиламенты). Во многих клетках органеллы могут принимать участие в образовании особых структур, характерных для специализированных клеток. Так, реснички и жгутики образуются за счет центриолей и плазматической мембраны, микроворсинки - это выросты плазматической мембраны с гиалоплазмой и микрофиламентами, акросома спермиев - это производное элементов аппарата Гольджи и пр.
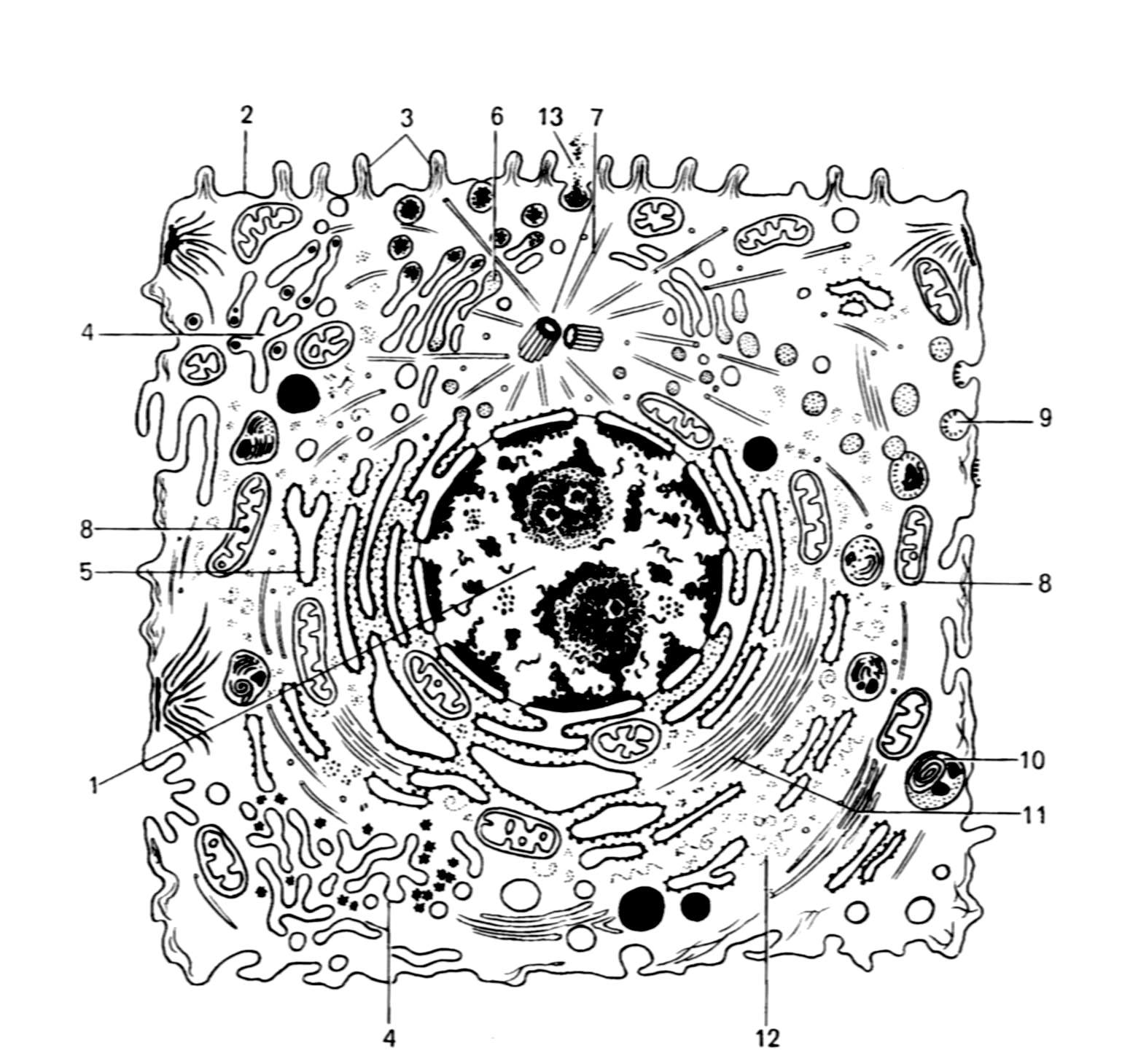
Рис 1. Ультрамикроскопическое строение клетки животных организмов (схема)
1 – ядро; 2 – плазмолемма; 3 – микроворсинки; 4 – агранулярная эндоплазматическая сеть; 5 - гранулярная эндоплазматическая сеть; 6 – аппарат Гольджи; 7 – центриоль и микротрубочки клеточного центра; 8 – митохондрии; 9 – цитоплазматические пузырьки; 10 – лизосомы; 11 – микрофиламенты; 12 – рибосомы; 13 – выделение гранул секрета.
Мембранные органеллы представляют собой одиночные или связанные друг с другом отсеки цитоплазмы, отграниченные мембраной от окружающей их гиалоплазмы, имеющие своё собственное содержимое, отличное по составу, свойствам и функциям:
Митохондрии - органеллы синтеза АТФ. Их основная функция связана с окислением органических соединений и использованием освобождающейся при распаде этих соединений энергии для синтеза молекул АТФ. Митохондрии ещё называют энергетическими станциями клетки, или органеллами клеточного дыхания.
Термин “”митохондрия” был введён Бенда в 1897 году. Митохондрии можно наблюдать в живых клетках, т.к. они обладают достаточно высокой плотностью. В живых клетках митохондрии могут перемещаться, сливаться друг с другом, делиться. Форма и размеры митохондрий животных клеток разнообразны, но в среднем толщина их около 0,5 мкм, а длина - от 1 до 10 мкм. Количество их в клетках сильно варьирует - от единичных элементов до сотен. Так, в клетке печени они составляют более 20% общего объема цитоплазмы. Площадь поверхности всех митохондрий печеночной клетки в 4-5 раз больше поверхности её плазматической мембраны.
Митохондрии ограничены двумя мембранами толщиной около 7 нм. Наружная митохондриальная мембрана ограничивает собственно внутреннее содержимое митохондрии, её матрикс. Характерной чертой внутренних мембран митохондрий является их способность образовывать многочисленные впячивания внутрь митохондрий. Такие впячивания чаще имеют вид плоских гребней, или крист. Нити матрикса митохондрий представляют собой молекулы ДНК, а мелкие гранулы - митохондриальные рибосомы.
Эндоплазматическая сеть была открыта К.Р. Портером в 1945 г. Эта органелла представляет собой совокупность вакуолей, плоских мембранных мешков или трубчатых образований, создающих как бы мембранную сеть внутри цитоплазмы. Различают два типа - гранулярную и гладкую эндоплазматическую сеть.
Гранулярная эндоплазматическая сеть представлена замкнутыми мембранами, отличительной чертой которых является то, что они со стороны гиалоплазмы покрыты рибосомами. Рибосомы участвуют в синтезе белков, выводимых из данной клетки. Кроме того, гранулярная эндоплазматическая сеть принимает участие в синтезе белков-ферментов, необходимых для организации внутриклеточного метаболизма, а также используемых для внутриклеточного пищеварения.
Белки, накапливающиеся в полостях сети, могут, минуя гиалоплазму, транспортироваться в вакуоли комплекса Гольджи, где они часто модифицируются и входят в состав либо лизосом, либо секреторных гранул.
Роль гранулярной эндоплазматической сети заключается в синтезе на её полисомах экспортируемых белков, в их изоляции от содержимого гиалоплазмы внутри мембранных полостей, в транспорте этих белков в другие участки клетки, а также в синтезе структурных компонентов клеточных мембран.
Агранулярная (гладкая) эндоплазматическая сеть также представлена мембранами, образующими мелкие вакуоли и трубки, канальцы, которые могут ветвиться друг с другом. В отличие от гранулярной эндоплазматической сети на мембранах гладкой эндоплазматической сети нет рибосом. Диаметр вакуолей и канальцев обычно около 50-100 нм.
Гладкая эндоплазматическая сеть возникает и развивается за счет гранулярной эндоплазматической сети.
Деятельность гладкой ЭПС связана с метаболизмом липидов и некоторых внутриклеточных полисахаридов. Гладкая ЭПС участвует в заключительных этапах синтеза липидов. Она сильно развита в клетках, секретирующих стероиды в корковом веществе надпочечников и сустентоцитах ( клетки Сертоли) семенников.
В поперечнополосатых мышечных волокнах гладкая ЭПС способна депонировать ионы кальция, необходимые для функции мышечной ткани.
Очень важна роль гладкой ЭПС в дезактивации различных вредных для организма веществ.
Комплекс Гольджи (КГ). В 1898 г. К. Гольджи, используя свойства связывания тяжелых металлов с клеточными структурами, выявил в нервных клетках сетчатые образования, которые он назвал внутренним сетчатым аппаратом.
Он представлен мембранными структурами, собранными вместе в небольшой зоне. Отдельная зона скопления этих мембран называется диктиосомой. Таких зон в клетке может быть несколько. В диктиосоме плотно друг к другу (на расстоянии 20-25 нм) расположены 5-10 плоских цистерн, между которыми располагаются тонкие прослойки гиалоплазмы. Кроме цистерн в зоне КГ наблюдается множество мелких пузырьков (везикул). КГ участвует в сегрегации и накоплении продуктов, синтезированных в цитоплазматической сети, в их химических перестройках, созревании; в цистернах КГ происходит синтез полисахаридов, их комплексирование с белками и, главное, выведение готовых секретов за пределы клетки.
Лизосомы - это разнообразный класс шаровидных структур размером 0,2-0,4 мкм, ограниченных одиночной мембраной.
Характерным признаком лизосом является наличие в них гидролитических ферментов, расщепляющих различные биополимеры. Лизосомы были открыты в 1949 г. де Дьювом.
Пероксисомы - небольшие размером 0,3-1,5 мкм овальной формы тельца, ограниченные мембраной. Они особенно характерны для клеток печени, почек. Ферменты окисления аминокислот образуют перекись водорода, который разрушается ферментом каталаза. Каталаза пероксисом играет важную защитную роль, так как Н2О2 является токсическим веществом для клетки.
Немембранные органеллы
Рибосомы - элементарные аппараты синтеза белковых, полипептидных молекул - обнаруживаются во всех клетках. Рибосомы - это сложные рибонуклеопротеиды, в состав которых входят белки и молекулы РНК. Размер функционирующей рибосомы эукариотических клеток 25 х 20 х 20 нм.
Различают единичные рибосомы и комплексные рибосомы (полисомы). Рибосомы могут располагаться свободно в гиалоплазме и быть связанными с мембранами эндоплазматической сети. Свободные рибосомы образуют белки в основном на собственные нужды клетки, связанные обеспечивают синтез белков “” на экспорт”.
Микротрубочки относятся к фибриллярным компонентам белковой природы. В цитоплазме они могут образовывать временные образования (веретено деления). Микротрубочки входят в состав центриолей, а также являются основными структурными элементами ресничек и жгутиков. Они представляют собой прямые, неветвящиеся длинные полые цилиндры. Их внешний диаметр составляет около 24 нм, внутренний просвет - 15 нм, толщина сетки - 5 нм. Микротрубочки содержат белки - тубулины. Создавая внутриклеточный скелет, микротрубочки могут быть факторами ориентированного движения клетки в целом и её внутриклеточных компонентов, создавая факторы направленных потоков разных веществ.
Центриоли. Термин был предложен Т. Бовери в 1895 г. для обозначения очень мелких телец. Центриоли обычно расположенные в паре - диплосома, окружены зоной более светлой цитоплазмы, от которой отходят радиально тонкие фибриллы (центросфера). Совокупность центриолей и центросферы называют клеточным центром. Эти органеллы в делящихся клетках принимают участие в формировании веретена деления и располагаются на его полюсах. В неделящихся клетках располагаются вблизи КГ.
Основой строения центриолей являются расположенные по окружности 9 триплетов микротрубочек, образующих таким образом полый цилиндр. Его ширина около 0,2 мкм, а длина 0,3-0,5 мкм.
Кроме микротрубочек в состав центриоли входят дополнительные структуры - “”ручки”, соединяющие триплеты. Системы микротрубочек центриоли можно описать формулой: (9 х 3) + 0, подчеркивая отсутствие микротрубочек в её центральной части.
При подготовке клеток к митотическому делению происходит удвоение центриолей.
Полагают, что центриоли участвуют в индукции полимеризации тубулином при образовании микротрубочек. Перед митозом центриоль является одним из центров полимеризации микротрубочек веретена клеточного деления.
Реснички и жгутики. Это специальные органеллы движения. В основании ресничек и жгутика в цитоплазме видны мелкие гранулы - базальные тельца. Длина ресничек 5-10 мкм, жгутиков - до 150 мкм.
Ресничка представляет собой тонкий цилиндрический вырост цитоплазмы с диаметром 200 нм. Он покрыт плазматической мембраной. Внутри расположена аксонема (“осевая нить”), состоящая из микротрубочек.
Аксонема в своём составе имеет 9 дуплетов микротрубочек. Здесь систему микротрубочек реснички опмсывают (9 х 2) + 2.
Свободные клетки, имеющие реснички и жгутики, обладают способностью двигаться. Способ их движения “”скользящие нити”.
К фибриллярным компонентам цитоплазмы относятся микрофиламенты толщиной 5-7 нм и так называемые промежуточные филаменты, микрофибриллы, толщиной около 10 нм.
Микрофиламенты встречаются во всех типах клеток. По строению и функциям они бывают разные, но отличить их морфологически друг от друга трудно. Химический состав их разный. Они могут выполнять функции цитоскелета и участвовать в обеспечении движения внутри клетки.
Промежуточные филаменты тоже белковые структуры. В эпителии в их состав входит кератин. Пучки филаментов образуют тонофибриллы, которые подходят к десмосомам. Роль промежуточных микрофиламентов скорее всего опорно-каркасная.
Включения цитоплазмы. Это необязательные компоненты клетки, возникающие и исчезающие в зависимости от метаболического состояния клеток. Различают включения трофические, секреторные, экскреторные и пигментные. Трофические включения - это нейтральные жиры и гликоген. Пигментные включения могут быть экзогенные (каротин, красители, пылевые частицы и др.) и эндогенные (гемоглобин, меланин и др.). Наличие их в цитоплазме может изменять цвет ткани. Нередко пигментация ткани служит диагностическим признаком.
Ядро обеспечивает две группы общих функций: одну, связанную собственно с хранением и передачей генетической информации, другую - с её реализацией, с обеспечением синтеза белка.
В ядре происходит воспроизведение или редупликация молекул ДНК, что дает возможность при митозе двум дочерним клеткам получить совершенно одинаковые в качественном и количественном отношении объемы генетической информации.
Другой группой клеточных процессов, обеспечиваемых активностью ядра, является создание собственного аппарата белкового синтеза. Это не только синтез, транскрипция на молекулах ДНК разных информационных РНК, но и транскрипция всех видов транспортных и рибосомальных РНК.
Таким образом, ядро является не только вместилищем генетического материала, но и местом, где этот материал функционирует и воспроизводиться.
Ядро неделящейся, интерфазной клетки обычно одно на клетку. Ядро состоит из хроматина, ядрышка, кариоплазмы (нуклеоплазмы) и ядерной оболочки, отделяющей его от цитоплазмы (кариолеммы).
Кариоплазма или ядерный сок - микроскопически бесструктурное вещество ядра. Он содержит различные белки (нуклеопротеиды, гликопротеиды), ферменты и соединения, участвующие в процессах синтеза нуклеиновых кислот, белков и др. веществ, входящих в состав кариоплазмы. Электронно - микроскопически в ядерном соке выявляют рибонуклеопротеидные гранулы 15 нм в диаметре.
В ядерном соке выявлены также гликолитические ферменты, участвующие в синтезе и расщеплении свободных нуклеотидов и их компонентов, энзимы белкового и аминокислотного обмена. Сложные процессы жизнедеятельности ядра обеспечиваются энергией, освобождающейся в процессе гликолиза, ферменты которого содержатся в ядерном соке.
Хроматин. В состав хроматина входит ДНК в комплексе с белком. Такими же свойствами обладают и хромосомы, которые отчетливо видны во время митотического деления клеток. Хроматин интерфазных ядер представляет собой хромосомы, которые теряют в это время свою компактную форму, разрыхляются, деконденсируются. Зоны полной деконденсации называются эухроматином; неполного разрыхления хромосом - гетерохроматином. Максимально конденсирован хроматин во время митотического деления клеток, когда он обнаруживается в виде плотных хромосом.
Ядрышко. Это одно или несколько округлой формы тельца величиной 1-5 мкм, сильно преломляющих свет. Его ещё называют нуклеолой. Ядрышко - самая плотная структура ядра - является производным хромосомы.
В настоящее время известно, что ядрышко - это место образования рибосомальных РНК и полипептидных цепей в цитоплазме.
Ядрышко неоднородно по своему строению: в световом микроскопе можно видеть его тонковолокнистую организацию. В электронном микроскопе выделяют два основных компонента: гранулярный и фибриллярный. Фибриллярный компонент - это рибонуклеопротеидные тяжи предшественников рибосом, гранулы - созревающие субъединицы рибосом.
Ядерная оболочка состоит из внешней ядерной мембраны и внутренней мембраны оболочки, разделенных перинуклеарным пространством. Ядерная оболочка содержит ядерные поры. Мембраны ядерной оболочки в морфологическом отношении не отличаются от остальных внутриклеточных мембран.
Поры имеют диаметр около 80-90 нм. Поперёк поры имеется диафрагма. Размеры пор у данной клетки обычно стабильны. Число пор зависит от метаболической активности клеток: чем интенсивнее синтетические процессы в клетках, тем больше пор на единицу поверхности клеточного ядра.
Хромосомы. Как интерфазные, так и митотические хромосомы состоят из элементарных хромосомных фибрилл - молекул ДНК.
Морфологию митотических хромосом лучше всего изучать в момент их наибольшей конденсации, в метафазе и в начале анафазы. Хромосомы в этом состоянии представляют собой палочковидные структуры разной длины и довольно постоянной толщины. У большинства хромосом удается легко найти зону первичной перетяжки (центромеры), которая делит хромосому на два плеча. Хромосомы с равными или почти равными плечами называют метацентрическими, с плечами неодинаковой длины - субметацентрическими. Палочковидные хромосомы с очень коротким, почти незаметным вторым плечом называют акроцентрическими. В области первичной перетяжки расположен кинетохор. От этой зоны во время митоза отходят микротрубочки клеточного веретена. Некоторые хромосомы имеют, кроме того, вторичные перетяжки, располагающиеся вблизи одного из концов хромосомы и отделяющие маленький участок - спутник хромосом. В этих местах локализована ДНК, ответственная за синтез рибосомальных РНК.
Совокупность числа, размеров и особенностей строения хромосом называют кариотипом данного вида. Кариотип крупного рогатого скота - 60, лошади - 66, свиньи - 40, овцы - 54, человека - 46.
Время существования клетки как таковой, от деления до деления или от деления до смерти называют клеточным циклом (рис.2).
Весь клеточный цикл состоит из 4 отрезков времени: собственно митоза, предсинтетического, синтетического и постсинтетического периодов интерфазы. В период G1 начинается рост клеток за счет накопления клеточных белков, что определяется увеличением количества РНК на клетку. В S - периоде происходит удвоение количества ДНК на ядро и соответственно удваивается число хромосом. Здесь уровень синтеза РНК возрастает соответственно увеличению количества ДНК, достигая своего максимума в G2-периоде. В G2-периоде происходит синтез информационной РНК, необходимой для прохождения митоза. Среди синтезирующихся в это время белков особое место занимают тубулины - белки митотического веретена.

Рис. 2. Жизненный цикл клетки:
М – митоз; G1 - предсинтетический период; S — синтетический период; G2 — постсинтетический период; 1 - старая клетка (2n4c); 2- молодые клетки (2n2c)
Преемственность хромосомного набора обеспечивается клеточным делением, которое называется митозом. Во время этого процесса происходит полная перестройка ядра. Митоз состоит из последовательного ряда стадий, сменяющихся в определенном порядке: профазы, метафазы, анафазы и телофазы. В процессе митоза ядро соматической клетки делится таким образом, что каждая из двух дочерних клеток получает точно такой же набор хромосом, какой имела материнская.
Способность клеток к воспроизведению - важнейшее свойство живой материи. Благодаря этой способности обеспечивается непрерывная преемственность клеточных поколений, сохранение клеточной организации в эволюции живого, совершается рост и регенерация.
По разным причинам (нарушение веретена деления, нерасхождение хроматид и др.) во многих органах и тканях встречаются клетки с крупными ядрами или многоядерные клетки. Это результат соматической полиплоидии. Такое явление называется эндорепродукцией. Чаще полиплоидия встречается у беспозвоночных животных. У некоторых из них распространено и явление политении - построение хромосомы из многих молекул ДНК.
Полиплоидные и политенные клетки не вступают в митоз и могут делиться только амитозом. Смысл данного явления в том, что как полиплоидия - увеличение количества хромосом, так и политения - увеличение количества молекул ДНК в хромосоме приводят к значительному усилению функциональной активности клетки.
Кроме митоза науке известны ешё два типа деления - амитоз (а - без, митоз - нити) или прямое деление и мейоз, который представляет собой процесс уменьшения числа хромосом вдвое путём двух клеточных делений - первого и второго деления мейоза (мейозис - уменьшение). Мейоз характерен для половых клеток.
Гаметогенез, стадии раннего эмбриогенеза
1. Строение половых клеток позвоночных.
2. Сперматогенез и овогенез.
3. Стадии раннего эмбриогенеза.
1. Эмбриология - наука о развитии зародыша. Она изучает индивидуальное развитие животных с момента зарождения (оплодотворение яйцеклетки) до его вылупления или рождения. Эмбриология рассматривает развитие и строение половых клеток и основные этапы эмбриогенеза: оплодотворение, дробление, гаструляцию, закладку осевых органов и органогенез, развитие провизорных (временных) органов.
Достижения современной эмбриологии широко используют в животноводстве, птицеводстве, при разведении рыб; в ветеринарии и медицине при решении многих практических задач, касающихся искусственного осеменения и оплодотворения, технологии ускоренного воспроизводства и селекции; повышения плодовитости с/х животных, размножения животных путем трансплантации эмбрионов, при изучении патологии беременности, при распознавании причин бесплодия и других вопросов акушерства.
По строению половые клетки сходны с соматическими клетками. Они также состоят из ядра и цитоплазмы, построенной из органелл и включений.
Отличительные свойства зрелых гаметоцитов - низкий уровень процессов ассимиляции и диссимиляции, неспособность к делению, содержание в ядрах гаплоидного (половины) числа хромосом.
Половые клетки самцов (спермии) у всех позвоночных имеют жгутиковую форму (рис.3). Образуются они в семенниках в большом количестве. В одной порции выделенного семени (эякулята) содержатся десятки миллионов и даже миллиардов спермиев.
Спермии с/х животных обладают подвижностью. Как размер, так и форма спермиев у разных животных сильно варьирует. Они состоят из головки, шейки и хвостового отдела. Спермии гетерогенны, т. к. в их ядрах содержатся разные типы половых хромосом. Половина спермиев имеет Х-хромосому, другая половина - Y хромосому. Половые хромосомы несут генетическую информацию, определяющую половые признаки самца. От остальных хромосом (аутосом) они отличаются большим содержанием гетерохроматина, размером и строением.
Спермии обладают минимальным запасом питательных веществ, которые очень быстро расходуются при движении клетки. Если не произойдет слияния спермия с яйцеклеткой, то в половых путях самки он обычно погибает через 24-36 часов.
Продлить жизнь спермиев можно замораживанием. Губительно влияют на спермии хинин, алкоголь, никотин и другие наркотические вещества.
Строение яйцеклеток. Размер яйцеклетки гораздо больше спермия. Диаметр овоцитов варьирует от 100 мкм до нескольких мм. Яйцеклетки позвоночных овальной формы, неподвижны, состоят из ядра и цитоплазмы (рис. 4). Ядро содержит гаплоидный набор хромосом. Яйцеклетки млекопитающих относят к гомогаметным, т. к. в их ядре имеется только Х-хромосома. В цитоплазме содержатся свободные рибосомы, эндоплазматическая сеть, комплекс Гольджи, митохондрии, желток и другие компоненты. Овоциты обладают полярностью. В связи с чем в них различают два полюса: апикальный и базальный. Периферический слой цитоплазмы яйцеклетки называют кортикальным слоем (cortex - кора). Он лишен полностью желтка, содержит множество митохондрий.
Яйцеклетки покрыты оболочками. Различают первичную, вторичную и третичную оболочки. Первичная оболочка - это плазмолемма. Вторичная оболочка (прозрачная или блестящая) является производной фолликулярных клеток яичника. Третичные оболочки формируются в яйцеводе у птиц: белок, подскорлуповая и скорлуповая оболочки яйца. По количеству желтка различают яйцеклетки с малым количеством - олиголецитальные (oligos - мало, lecytos - желток), со средним количеством - мезолецитальные (mesos - средний) и с большим количеством - полилецитальные (poli - много).
По месту расположения желтка в цитоплазме различают яйцеклетки с равномерным распределением желтка - изолецитальные, или гомолецитальные, и с локализацией желтка у одного полюса - телолецитальные ( telos - край, конец). Олиголецитальные и изолецитальные яйцеклетки - у ланцетника и млекопитающих, мезолецитальные и телолецитальные - у амфибий, некоторых рыб, полилецитальные и телолецитальные - у многих рыб, пресмыкающихся, птиц.
2. Родоначальниками половых клеток являются первичные половые клетки - гаметобласты (гонобласты). Выявляются они в стенке желточного мешка вблизи кровеносных сосудов. Гонобласты интенсивно делятся митозом и с током крови или по ходу кровеносных сосудов мигрируют в зачатки половых желез, где окружаются поддерживающими (фолликулярными) клетками. Последние выполняют трофическую функцию. Затем, в связи с развитием пола животного, половые клетки приобретают свойства, характерные для спермиев и яйцеклеток.
Развитие спермиев (сперматогенез) протекает в семенниках половозрелого животного. В сперматогенезе различают 4 периода: размножение, рост, созревание и формирование.
Период размножения. Клетки называются сперматогониями. Они имеют небольшие размеры, диплоидное число хромосом. Клетки интенсивно делятся митозом. Делящиеся клетки являются стволовыми клетками и пополняют запас сперматогоний.
Период роста. Клетки называются первичными сперматоцитами. У них сохраняется диплоидное число хромосом. Увеличивается размер клетки и происходят сложные изменения в перераспределении наследственного материала в ядре, в связи с чем различают четыре стадии: лептотенную, зиготенную, пахитенную, диплотенную
Период созревания. Это процесс развития сперматид с половинным числом хромосом.
В процессе созревания из каждого первичного сперматоцита возникает 4 сперматиды с одинарным числом хромосом. В них хорошо развиты митохондрии, комплекс Гольджи, центросома, расположены вблизи ядра. Другие органеллы, а также включения почти отсутствуют. Сперматиды не способны делиться.
Период формирования. Сперматида приобретает морфологические свойства, характерные для спермия. Комплекс Гольджи преобразуется в акросому, в виде чехлика охватывающую ядро сперматиды. Акросома богата ферментом гиалуронидазой. К противоположному от ядра полюсу перемещается центросома, в которой различают проксимальную и дистальную центриоли. Проксемальная центриоль остается в шейке спермия, а дистальная – идет на построение хвостика.
Развитие яйцеклеток, овогенез - сложный и очень длительный процесс. Он начинается в период эмбриогенеза и завершается в органах половой системы половозрелой самки. Слагается овогенез из трех периодов: размножения, роста, созревания.
Период размножения протекает в период внутриутробного развития и завершается в течение первых месяцев после рождения. Клетки называются овогониями, имеют диплоидное число хромосом.
В период роста клетки называются первичными ооцитами. Изменения в ядрах аналогичны первичным сперматоцитам. Затем в ооците начинается интенсивный синтез и накопление желтка: стадия превителлогенеза и стадия вителлогенеза. Вторичная оболочка ооцита состоит из одного слоя фолликулярных клеток. Превителлогенез обычно длится до наступления половой зрелости самки. Период созревания состоит из быстро следующих друг за другом делений созревания, в ходе которых диплоидная клетка становится гаплоидной. Этот процесс обычно протекает в яйцеводе после овуляции.
Первое деление созревания завершается образованием двух неравноценных структур - вторичного ооцита и первого направительного или редукционного тельца . В ходе второго деления также образуется одна зрелая яйцеклетка и второе направительное тельце. Первое тельце также делится. Следовательно, из одного первичного ооцита в процессе созревания возникает только одна зрелая яйцеклетка и три направительных тельца последние вскоре гибнут.
Все яйцеклетки генетически однородны, т. к. имеют только Х-хромосому.
3. Оплодотворение - слияние половых гамет и образование нового одноклеточного организма (зиготы). От зрелой яйцеклетки она отличается удвоенной массой ДНК, диплоидным числом хромосом. Оплодотворение у млекопитающих внутреннее, происходит оно в яйцеводе при пассивном её передвижении по направлению к матке. Движение спермиев в половых путях самки осуществляется благодаря функции аппарата движения этой клетки (хемотаксиса и реотаксиса), перистальтическим сокращениям стенки матки, движению ресничек, покрывающих внутреннюю поверхность яйцевода. При сближении половых клеток ферменты акросомы головки спермия разрушают слой фолликулярных клеток, вторичную оболочку яйцеклетки. В момент прикосновения спермия к плазмолемме яйцеклетки на её поверхности образуется выпячивание цитоплазмы - бугорок оплодотворения. В ооцит проникают головка и шейка. У млекопитающих в оплодотворении участвует только один спермий - поэтому процесс называется моноспермией: ХY - самец, XX - самка.
У птиц, рептилий наблюдается полиспермия. У птиц все спермии имеют Z - хромосому, а яйцеклетки Z или W - хромосому.
После проникновения спермия в яйцеклетку вокруг последней формируется оболочка оплодотворения, препятствующая проникновению в ооцит других спермиев ядра половых клеток называются: мужской пронуклеус, женский пронуклеус. Процесс их соединения называется синкарионом. Центриоль, привнесенная спермием, делится и расходится, образуется ахроматиновое веретено. Начинается дробление. Дробление - дальнейший процесс развития одноклеточной зиготы, в ходе которого образуется многоклеточная бластула, которая состоит из стенки - бластодермы и полости - бластоцеля. В процессе митотического деления зиготы образуются новые клетки - бластомеры.
Характер дробления у хордовых различен и в значительной степени обусловлен типом яйцеклетки. Дробление может быть полным (голобластическим) или частичным (меробластическим). При первом типе принимает участие весь материал зиготы, при втором - только та ее зона, которая лишена желтка.
Полное дробление классифицируют на равномерное и неравномерное. Первое характерно для олиго изолецитальных яиц (ланцетник, аскарида и др.). В оплодотворенной яйцеклетке различают два полюса: верхний - анимальный и нижний - вегететивный. После оплодотворения желток перемещается к вегетативному полюсу.
Дробление завершается образованием бластулы, форма которой напоминает шар, заполненный жидкостью. Стенка шара образована клетками бластодермы. Таким образом, при полном равномерном дроблении материал всей зиготы участвует в дроблении и после каждого деления число клеток увеличивается вдвое.
Полное неравномерное дробление характерно для мезолецитальных (среднее количество желтка) и телолецитальных яйцеклеток. Это амфибии. Тип бластулы у них – целобластула.
Частичное, или меробластическое (дискоидальное) дробление распространено у рыб, птиц и характерно для полилецитальных и телолецитальных яиц (тип бластулы называется дискобластулой).
Гаструляция. При дальнейшем развитии бластулы в процессе деления, роста , дифференцировки клеток и их перемещений формируется сначала двух-, а затем трехслойный зародыш. Его слоями являются эктодерма, энтодерма и мезодерма.
Типы гаструляции: 1) инвагинация, 2) эпиболия (обрастание), 3) иммиграция (вселение), 4) деляминация (расслоение).
Закладка осевых органов. Из указанных зародышевых листков образуются осевые органы: зачаток нервной системы ( нервная трубка), хорда и кишечная трубка.
В процессе развития мезодермы у всех позвоночных образуется хорда, сегментированная мезодерма, или сомиты (спинные сегменты), и несегментированная мезодерма, или спланхнотом. Последний состоит из двух листков: наружного - париетального и внутреннего - висцерального. Пространство между этими листками называется вторичной полостью тела.
В сомитах различают три зачатка: дерматом, миотом, склеротом. Нефрогонадотом.
При дифференцировке зародышевых листков образуется эмбриональная ткань - мезенхима. Она развивается из клеток, выселившихся главным образом из мезодермы и эктодермы. Мезенхима - это источник развития соединительной ткани, гладких мышц, сосудов и других тканей организма животного. Процессы дробления у различных представителей хордовых очень своеобразны и зависят от проморфологии яиц, в особенности от количества и распределения желтка. Процессы гаструляции также весьма варьируют в пределах Chordata.
Так, гаструляция у ланцетника типично инвагинационная, начинается она впячиванием презумптивной энтодермы. Вслед за энтодермой инвагинирует в бластоцель материал хорды, а через боковую и вентральную губы бластопора погружается мезодерма. Передняя (или дорсальная) губа бластопора состоит из материала будущей нервной системы, а изнутри из клеток будущей хорды. Как только энтодермальный пласт приходит в контакт с внутренней стороны эктодермального пласта, начинаются процессы, приводящие к формированию зачатков осевых органов.
Процесс гаструляции у костистых рыб начинается тогда, когда многослойный бластодиск покрывает только небольшую часть желтка яйца, а заканчивается вместе с полным обрастанием всего ”желточного шара”. Это означает, что гаструляция включает и разрастание бластодиска.
Клеточный материал всех трех зародышевых пластов по переднему и боковым краям бластодиска начинают нарастать на желток. Таким образом формируется так называемый желточный мешок.
Желточный мешок как часть зародыша выполняет многообразные функции:
1) это орган с трофической функцией, т. к. дифференцирующийся энтодермальный пласт продуцирует ферменты, помогающие расщеплять вещества желтка, а в дифференцирующемся мезодермальном пласте образуются кровеносные сосуды, находящиеся в связи с сосудистой системой собственно зародыша.
2) желточный мешок - орган дыхания. Газообмен зародыша с внешней средой происходит через стенки сосудов мешка и эктодермальный эпителий.
3) “кровяная мезенхима” является клеточной основой кроветворения. Желточный мешок - первый кроветворный орган зародыша.
Лягушки, тритоны и морские ежи являются главнейшими объектами экспериментальных эмбриологических исследований в ХХ веке.
Инвагинация у амфибий не может происходить так, как у ланцетника, потому что вегетативное полушарие яйца очень перегружено желтком.
Первый заметный признак начинающейся гаструляции у лягушек - это появление бластопора, т. е. вдавления или щели в середине серого серпа.
Достойно особого внимания поведение клеточного материала нервной системы и эпидермиса кожи. В конце концов, будущий эпидермис и материал нервной системы покрывает всю поверхность зародыша. Презумптивный эпидермис кожи перемещается и истончается во всех направлениях. Совокупность клеток презумптивной нервной системы перемещается почти исключительно в меридиональных направлениях. Пласт клеток будущей нервной системы в поперечном направлении сокращается, презумптивная область нервной системы оказывается вытянутой в анимально-вегетативном направлении.
Обобщим известное нам о судьбе каждого из зародышевых листков.
Производные эктодермы. Из клеток, составляющих наружный пласт, размножаясь и дифференцируясь, формируются: наружный эпителий, кожные железы, поверхностный слой зубов, роговых чешуй и т. п. Кстати сказать, почти всегда каждый орган развивается из клеточных элементов двух, а то и всех трех зародышевых листков. Например, кожа млекопитающих развивается из эктодермы и мезодермы.
Обширная часть первичной эктодермы “погружается” внутрь, под наружный эпителий, и дает начало всей нервной системе.
Производные энтодермы. Внутренний зародышевый пласт развивается в эпителий средней кишки и ее пищеварительные железы. Эпителий дыхательной системы развивается из переднего отдела кишечника. Но в его происхождении участвует клеточный материал так называемой прехордальной пластинки.
Производные мезодермы. Из неё развиваются все мышечные ткани, все виды соединительной, хрящевой, костной тканей, каналы выделительных органов, перитонеум полости тела, кровеносная система, часть тканей яичников и семенников.
У большинства животных средний пласт появляется не только в виде совокупности клеток, образующих компактный эпителиальный слой, т. е. собственно мезодерму, но в виде рыхлого комплекса разрозненных, амебоподобных клеток. Эта часть мезодермы называется мезенхимой. Собственно мезодерма и мезенхима отличаются друг от друга по своему происхождению, между ними нет прямой связи, они не гомологичны. Мезенхима большей частью эктодермального происхождения, начало же мезодерме дает энтодерма. У позвоночных, однако, мезенхима имеет общее с остальной мезодермой происхождение.
У всех животных, которым свойственно иметь целом (вторичная полость тела), начало полым целомическим мешкам дает мезодерма. Целомические мешки формируются симметрично по бокам кишечника. Стенка каждого целомического мешка, обращенная в сторону кишечника называется спланхноплеврой. Стенка же, обращенная в сторону эктодермы зародыша, называется соматоплеврой.
Таким образом, в ходе развития зародыша формируются различные полости, имеющие важное морфогенетическое значение. Сначала появляется полость Бэра, превращающаяся в первичную полость тела - бластоцель, затем возникает гастроцель (или гастральная полость), наконец у многих животных - целом. При образовании гастроцеля и целома бластоцель все более уменьшается, так что от бывшей первичной полости тела остаются лишь щели в промежутках между стенками кишки и целома. Эти щели превращаются в полости кровеносной системы. Гастроцель со временем превращается в полость средней кишки.
Особенности эмбриогенеза млекопитающих и птиц
1. Внезародышевые органы.
2. Плацента млекопитающих.
3. Стадии пренатального периода онтогенеза жвачных, свиней и птиц.
1. У зародышей рептилий и птиц также формируется желточный мешок. В этом участвуют все зародышевые пласты. В течение 2-го и 3-го дней развития куриного зародыша во внутренней части area opaca развивается сеть кровеносных сосудов. Их появление неразрывно связано с возникновением эмбрионального кроветворения. Таким образом, одна из функций желточного мешка зародышей птиц - эмбриональное кроветворение. В самом зародыше лишь впоследствии образуются кроветворные органы - печень, селезенка, костный мозг.
Сердце зародыша начинает функционировать (сокращаться) в конце второго дня, с этого времени возникает кровоток.
У зародышей птиц кроме желточного мешка образуются ещё три провизорных органа, которые принято называть зародышевыми оболочками, - амнион, сероза и аллантоис. Эти органы можно рассматривать как выработавшиеся в процессе эволюции адаптации эмбрионов.
Амнион и сероза возникают в теснейшей взаимосвязи. Амнион в виде поперечной складки, нарастая, изгибается над передним концом головы зародыша и покрывает его, как капюшон. В дальнейшем боковые участки амниотических складок нарастают по обеим сторонам собственно зародыша и срастаются. Амниотические складки состоят из эктодермы и париетального листка мезодермы.
Сопряженно со стенкой амниотической полости развивается еще одно важное провизорное образование – сероза , или серозная оболочка. Она состоит из эктодермального листка, ”смотрящего” на зародыш, и мезодермального, ”смотрящего” наружу. Наружная оболочка разрастается по всей поверхности под скорлупой. Это и есть сероза.
Амнион и сероза являются, конечно, ”оболочками”, т. к. действительно покрывают и объединяют собственно эмбрион от внешней среды. Однако это - органы, части зародыша с весьма важными функциями. Амниотическая жидкость создает водную среду для эмбрионов животных, в ходе эволюции ставших сухопутными. Она предохраняет развивающийся зародыш от высыхания, от сотрясений, от прилипания к оболочке яйца. Интересно отметить, что роль амниотической жидкости у млекопитающих отметил ещё Леонардо да Винчи.
Серозная оболочка принимает участие в дыхании и резорбции остатков белковой оболочки (под действием выделяемых хорионом ферментов).
Развивается ещё один провизорный орган - аллантоис, выполняющий сначала функцию зародышевого мочевого пузыря. Он появляется как вентральный вырост энтодермы задней кишки. У куриного зародыша этот выступ появляется уже на 3 день развития. В середине эмбрионального развития птиц аллантоис разрастается под хорионом по всей поверхности зародыша с желточным мешком.
В самом конце эмбрионального развития птиц (и рептилий) провизорные органы зародыша постепенно прекращают свои функции, они редуцируются, зародыш начинает дышать воздухом, имеющимся внутри яйца (в воздушной камере), пробивает скорлупу, освобождается от яйцевых оболочек и оказывается во внешней среде.
Внезародышевыми органами млекопитающих является желточный мешок, амнион, аллантоис, хорион и плацента (рис. 5).
2. У млекопитающих связь эмбриона с материнским организмом обеспечивается формированием специального органа - плаценты (детского места). Источником его развития является алланто-хорион. Плаценты по своему строению разделяются на несколько типов. В основе классификации положено два принципа: а) характер распределения ворсинок хориона и 2) способ их связи со слизистой матки (рис. 6).
По форме различают несколько типов плаценты:
1) Диффузная плацента (эпителиохориальная) - вторичные сосочки ее развиваются по всей поверхности хориона. Ворсинки хориона проникают в железы стенки матки, не разрушая при этом ткани матки. Питание зародыша осуществляется посредством маточных желез, секретирующих маточное молочко, которое всасывается в кровеносные сосуды ворсинок хориона. При родах ворсинки хориона выдвигаются из маточных желез без разрушения тканей. Такая плацента характерна для свиньи, лошади, верблюда, сумчатых, китообразных, бегемота.

Рис. 5. Схема развития желточного мешка и зародышевых оболочек у млекопитающих (шесть последовательных стадий):
А — процесс обрастания полости плодного пузыря энтодермой (1) и мезодермой (2); Б — образование замкнутого энтодермального пузырька (4); В — начало образования амниотической складки (5) и кишечного желобка (6); Г — обособление тела зародыша (7); желточный мешок (8); Д — смыкание амниотических складок (9); начало образования развития аллантоиса (10); Е — замкнутая амниотическая полость (11); развитый аллантоис (12); ворсинки хориона (13); париетальный листок мезодермы (14); висцеральный листок мезодермы (15); эктодерма (3).
2) Котиледонная плацента (десмохориальная) - ворсинки хориона расположены кустиками - котиледонами. Они соединяются с утолщениями стенки матки, которые именуются карункулами. Комплекс котиледон-карункул называется плацентомом. Такая плацента свойственна жвачным.
3) Поясная плацента (эндотелиохориальная) - ворсинки в виде широкого пояса окружают плодный пузырь и располагаются в соединительнотканном слое стенки матки, контактируя с эндотелиальным слоем стенки кровеносных сосудов.
4) Дискоидальная плацента (гемохориальная) - зона контакта ворсинок хориона и стенки матки имеет форму диска. Ворсинки хориона погружаются в заполненные кровью лакуны, лежащие в соединительнотканном слое стенки матки. Такая плацента встречается у приматов.
3. Работники животноводства своей практической деятельностью разводят и выращивают животных. Это сложные биологические процессы, и, чтобы сознательно управлять или искать пути их совершенствования, зооинженер и ветеринарный врач должны знать основные закономерности развития животных на протяжении их индивидуальной жизни. Нам уже известно, что цепь изменений, которую переживает организм от момента своего возникновения до естественной смерти, называется онтогенезом. Он слагается из качественно различных периодов. Однако периодизация онтогенеза разработана ещё недостаточно. Одни ученые считают, что онтогенетическое развитие организма начинается с развития половых клеток, другие - с образования зиготы.

А Б В Г
Рис. 6. Типы гистологического строения плацент:
А — эпителиохориальная; Б — десмохориальная; В — эндотелиохориальная: Г — гемохориальная; I — зародышевая часть; II — материнская часть; 1 — эпителий: 2 — соединительная ткань и 3 — эндотелий кровеносного сосуда ворсинки хориона; 4 — эпителий; 5 — соединительная ткань и 6 — кровеносные сосуды и лакуны слизистой оболочки матки.
После возникновения зиготы последующий онтогенез с/х животных разделяется на внутриутробное и послеутробное развитие.
Продолжительность подпериодов внутриутробного развития с/х животных, сутки (по Г.А. Шмидту).
|
Вид животного |
Подпериоды внутриутробного развития |
||
|
зародышевый |
предплодный |
плодный |
|
|
Кролик |
1-12 |
13-18 |
19-30 |
|
Свинья |
1-22 |
23-38 |
39-114 |
|
Овца |
1-28 |
29-46 |
47-150 |
|
Крупный рогатый скот |
1-34 |
35-60 |
61-284 |
В эмбриогенезе животных благодаря их родству имеются некоторые принципиально сходные черты: 1) образование зиготы, 2) дробление, 3) образование зародышевых листков, 4) дифференцировка зародышевых листков, приводящая к образованию тканей и органов.
Общая гистология. Эпителиальные ткани
1. Развитие тканей.
2. Классификация эпителиальных тканей.
3. Железы и критерии их классификации.
1. Организм животных построен из клеток и неклеточных структур, специализированных на выполнении определенных функций. Популяции клеток, разные по функции, отличаются строением и специфичностью синтеза внутриклеточных белков.
В процессе развития первоначально однородные клетки приобрели различия в обмене веществ, строении, функции. Этот процесс называется дифференциацией. При этом реализуется генетическая информация, исходящая от ДНК клеточного ядра, которая проявляется в конкретных условиях. Приспособление клеток к этим условиям называется адаптацией.
Дифференцировка и адаптация обусловливают развитие между клетками и их популяциями качественно новых взаимосвязей и отношений. При этом в значительной степени возрастает значение целостности организма, т. е. интеграция. Так, каждая стадия эмбриогенеза - это не просто увеличение числа клеток, а новое состояние целостности.
Интеграция - это объединение клеточных популяций в более сложные функционирующие системы - ткани, органы. Нарушить её можно вирусами, бактериями, лучами Рентгена, гормонами и др. факторами. В этих случаях биологическая система выходит из-под контроля, что может послужить причиной развития злокачественных опухолей и других патологий.
Морфофункциональные и генетические различия, возникшие в процессе филогенеза, позволили клеткам и неклеточным структурам объединиться в так называемые гистологические ткани.
Тканью называется исторически сложившаяся система клеток и неклеточных структур, характеризующаяся общим строением, функцией и происхождением.
Различают четыре основных типа тканей: эпителиальные, соединительные или опорно-трофические, мышечные и нервная. Существуют и другие классификации.
2. Эпителиальные ткани осуществляют связь организма с внешней средой. Они выполняют покровную и железистую (секреторную) функции. Эпителий расположен в кожном покрове, выстилает слизистые оболочки всех внутренних органов; у него функции всасывания, выделения. Большинство желез организма построено из эпителиальной ткани.
В развитии эпителиальной ткани принимают участие все зародышевые листки.
Все эпителии построены из эпителиальных клеток - эпителиоцитов. Соединяясь прочно друг с другом с помощью десмосом, поясков замыкания, поясков склеивания и путем интердигитации эпителиоциты образуют клеточный пласт, функционирующий и регенерирующий. Обычно пласты расположены на базальной мембране, которая, в свою очередь лежит на рыхлой соединительной ткани, питающей эпителий (рис. 7).
Эпителиальные ткани характеризуются полярной дифференциацией, которая сводится к разному строению или слоев эпителиального пласта, или полюсов эпителиоцитов. Например, на апикальном полюсе плазмолемма образует всасывающую каемку или мерцательные реснички, а в базальном полюсе находится ядро и большинство органелл.
В зависимости от места расположения и выполняемой функции различают два типа эпителиев: покровные и железистые.
В основу наиболее распространенной классификации покровных эпителиев положены форма клеток и количество слоев в эпителиальном пласте, поэтому она именуется морфологической.
3. Эпителии, вырабатывающие секреты, называются железистыми, а его клетки - секреторными клетками, или секреторными гландулоцитами. Из секреторных клеток построены железы, которые могут быть оформлены в виде самостоятельного органа или является только его частью.
Различают эндокринные и экзокринные железы. Морфологически разница в наличии выводного протока у вторых. Экзокринные железы могут быть одно- и многоклеточными. Пример: бокаловидная клетка в простом столбчатом каёмчатом эпителии. По характеру ветвления выводного протока различают простые и сложные. У простых желез неветвящийся выводной проток, у сложных - ветвящийся. Концевые отделы у простых желез разветвляются и неразветвляются, у сложных - разветвляются.
По форме концевых отделов экзокринные железы классифицируют на альвеолярные, трубчатые и трубчато-альвеолярные. Клетки концевого отдела именуются гландулоцитами.
По способу образования секрета железы делят на голокринные, апокринные и мерокринные. Это сальные, затем потовые и молочные, железы желудка соответственно.
Регенерация. Покровные эпителии занимают пограничное положение. Они часто повреждаются, поэтому характеризуются высокой регенерационной способностью. Регенерация осуществляется главным образом митотическим способом. Клетки эпителиального пласта быстро изнашиваются, стареют и гибнут. Их восстановление называется физиологической регенерацией. Восстановление эпителиальных клеток, утраченных по причине травмы, называется репаративной регенерацией.
В однослойных эпителиях регенерационной способностью обладают все клетки, в многослойных - стволовые. В железистом эпителии при голокриновой секреции такой способностью обладают стволовые клетки, находящиеся на базальной мембране. В мерокринных и апокринных железах восстановление эпителиоцитов протекает главным образом, путем внутриклеточной регенерации.

Рис. 7. Схема различных типов эпителия
А. Однослойный плоский.
Б. Однослойный кубический.
В. Однослойный цилиндрический.
Г. Многорядный цилиндрический мерцательный.
Д. Переходный.
Е. Многослойный плоский неороговевающий.
Ж. Многослойный плоский ороговевающий.
Опорно-трофические ткани. кровь и лимфа
1. Кровь. Клетки крови.
2. Лимфа.
3. Гемоцитопоэз.
4. Эмбриональный гемоцитопоэз.
С этой темы начинаем изучение группы родственных тканей, именуемых как соединительные. Сюда входят: собственно соединительная ткань, клетки крови и кроветворных тканей, скелетные ткани ( хрящевая и костная ), соединительные ткани со специальными свойствами.
Проявление единства перечисленных выше видов ткани является происхождение их из общего эмбрионального источника - мезенхимы.
Мезенхима - совокупность эмбриональных сетевидно связанных отростчатых клеток, заполняющих промежутки между зародышевыми листками и зачатками органов. В теле зародыша мезенхима возникает главным образом из клеток определенных участков мезодермы - дерматомов, склеротомов и спланхнотомов. Клетки мезенхимы быстро делятся митозом. В различных её участках возникают многочисленные мезенхимные производные - кровяные островки с их эндотелием и клетками крови, клетки соединительных тканей и гладкой мышечной ткани и др.
1. Внутрисосудистая кровь - подвижная тканевая система с жидким межклеточным веществом плазмой и форменными элементами - эритроцитами, лейкоцитами и кровяными пластинками.
Постоянно циркулируя в замкнутой системе кровообращения, кровь объединяет работу всех систем организма и поддерживает многие физиологические показатели внутренней среды организма на определенном, оптимальном для осуществления обменных процессов уровне. Кровь выполняет в организме разносторонние жизненно важные функции: дыхательную, трофическую, защитную, регуляторную, выделительную и другие.
Несмотря на подвижность и изменяемость крови, её показатели в каждый момент соответствуют функциональному состоянию организма, поэтому исследование крови является одним из важнейших диагностических методов.
Плазма - жидкая составная часть крови, содержит 90-92% воды и 8-10% сухих веществ, в т. ч. 9% органических и 1% минеральных веществ. Основные органические вещества плазмы крови - белки (альбумины, различные фракции глобулинов и фибриноген). Иммунные белки (антитела), а большинство их содержится в гамма-глобулиновой фракции, называют иммуноглобулинами. Альбумины обеспечивают перенос различных веществ - свободных жирных кислот, билирубина и др. Фибриноген принимает участие в процессах свертывания крови.
Эритроциты являются основным типом клеток крови, т. к. их в 500-1000 раз больше, чем лейкоцитов. В 1мм 3 крови крупного рогатого скота содержится 5,0-7,5млн, лошади - 6-9 млн., овцы - 7-12 млн., козы - 12-18 млн., свинья - 6-7,5 млн., куры - 3-4 млн. эритроцитов.
Утратив в процессе развития ядро, зрелые эритроциты у млекопитающих являются безъядерными клетками и имеют форму двояковогнутого диска со средним диаметром круга 5-7 мкм. Эритроциты крови верблюда и ламы овальные. Дисковидная форма увеличивает общую поверхность эритроцита в 1,64 раза.
Между количеством эритроцитов и их величиной существует обратная зависимость.
Эритроциты покрыты оболочкой - плазмолеммой(толщиной 6 нм), содержащей 44% липидов, 47% белков и 7% углеводов. Мембрана эритроцитов легко проницаема для газов, анионов, ионов Na.
Внутреннее коллоидное содержимое эритроцитов на 34% состоит из гемоглобина - уникального сложного окрашенного соединения - хромопротеида, в небелковой части которого (геме) имеется двухвалентное железо, способное образовывать особые непрочные связи с молекулой кислорода. Именно благодаря гемоглобину осуществляется дыхательная функция эритроцитов. Оксигемоглобин = гемоглобин+О2.
Наличие гемоглобина в эритроцитах обуславливает выраженную оксифилию их при окраске мазка крови по Романовскому- Гимзе (эозин + азур II). Эритроциты при этом окрашиваются в красный цвет эозином. При некоторых формах анемий центральная бледно окрашенная часть эритроцитов увеличена - гипохромные эритроциты. При суправитальном окрашивании крови бриллиантовым крезиловым синим можно обнаружить молодые формы эритроцитов, содержащие зернисто-сетчатые структуры. Такие клетки называют ретикулоцитами, они являются непосредственными предшественниками зрелых эритроцитов. Подсчет ретикулоцитов используется для получения информации о скорости образования эритроцитов.
Период жизни эритроцита 100-130 дней ( у кроликов 45-60 дней). Эритроциты обладают свойством противостоять различным разрушительным воздействиям - осмотическим, механическим и др. При изменениях концентрации солей в окружающей среде мембрана эритроцита перестаёт удерживать гемоглобин, и он выходит в окружающую жидкость - явление гемолиза. Выход гемоглобина может происходить в организме при действии змеиного яда, токсинов. Гемолиз развивается также при переливании несовместимой по группе крови. Практически важно при введении в кровь животным жидкостей осуществлять контроль за тем, чтобы вводимый раствор был изотоническим.
У эритроцитов по сравнению с плазмой и лейкоцитами крови относительно большая плотность. Если кровь обработать противосвертывающими веществами и поместить в сосуд, то отмечают оседание эритроцитов. Скорость оседания эритроцитов (СОЭ) у животных разного возраста, пола и вида неодинакова. Высокая СОЭ у лошадей и, наоборот, низкая у крупного рогатого скота. СОЭ имеет диагностическое и прогностическое значение.
Лейкоциты - разнообразные по морфологическим признакам и функциям клетки сосудистой крови. В организме животных они выполняют многообразные функции, направленные прежде всего на защиту организма от чужеродного влияния путем фагоцитарной активности, участия в формировании гуморального и клеточного иммунитета, а также в восстановительных процессах при тканевых повреждениях. В 1 мм3 крови у крупного рогатого скота их насчитывается 4,5-12 тыс., у лошадей - 7-12 тыс., овец - 6-14 тыс., свиней - 8-16 тыс., кур - 20-40 тыс. Увеличение количества лейкоцитов - лейкоцитоз - характерный признак для многих патологических процессов.
Образовавшись в кроветворных органах и поступив в кровь, лейкоциты лишь непродолжительное время пребывают в сосудистом русле, затем мигрируют в вокруг сосудистую соединительную ткань и органы, где осуществляют свою основную функцию.
Особенность у лейкоцитов та, что они обладают подвижностью за счет образующихся псевдоподий. В лейкоцитах различают ядро и цитоплазму, содержащую различные органеллы и включения. Классификация лейкоцитов основана на способности окрашиваться красителями и зернистости.
Лейкоциты зернистые (гранулоциты): нейтрофилы (25-70%), эозинофилы (2-12%), базофилы (0,5-2%).
Лейкоциты незернистые (агранулоциты): лимфоциты (40-65) и моноциты (1-8%).
Определенное процентное соотношение между отдельными видами лейкоцитов называется лейкоцитарной формулой - лейкограммой.
Увеличение в лейкограмме процента нейтрофилов типично для гнойно-воспалительных процессов. У зрелых нейтрофилов ядро состоит из нескольких сегментов, соединенных тонкими перемычками.
По сравнению с нейтрофилами эозинофилы менее подвижны и обладают меньшей фагоцитарной активностью. Они участвуют в ограничении воспалительного процесса. Эозинофилы являются важнейшими эффекторными клетками в противопаразитарном иммунитете.
На поверхности базофилов расположены специальные рецепторы, с помощью которых связываются иммуноглобулины Е. Они участвуют в иммунологических реакциях аллергического типа.
Циркулирующие в крови моноциты являются предшественниками тканевых и органных макрофагов. После пребывания в сосудистой крови (12-36 ч) моноциты мигрируют через эндотелий капилляров и венул в ткани и превращаются в подвижные макрофаги.
Лимфоциты - важнейшие клетки, участвующие в разнообразных иммунологических реакциях организма. Большое количество лимфоцитов находится в лимфе.
Различают два основных класса лимфоцитов: Т- и В-лимфоциты. Первые развиваются из костномозговых клеток в корковой части долек тимуса. В плазмолемме имеют антигенные маркеры и многочисленные рецепторы, с помощью которых происходит распознавание чужеродных антигенов и иммунных комплексов.
В-лимфоциты образуются из стволовых предшественников в фабрициевой сумке (Bursa). Местом развития их считают миелоидную ткань костного мозга.
Эффекторными клетками в системе Т-лимфоцитов являются три основные субпопуляции: Т-киллеры (цитотоксические лимфоциты), Т-хелперы (помощники) и Т-супрессоры (угнетающие). Эффекторными клетками В-лимфоцитов являются плазмобласты и зрелые плазмоциты, способные в повышенном количестве продуцировать иммуноглобулины.
Кровяные пластинки - безъядерные элементы сосудистой крови млекопитающих. Это небольшие цитоплазматические фрагменты мегакариоцитов красного костного мозга. В 1 мм3 крови их насчитывается 250-350 тыс. кровяных пластинок. У птиц сходные по функции клетки называются тромбоцитами.
Кровяные пластинки имеют важнейшее знание в обеспечении основных этапов остановки кровотечения - гемостаза.
2. Лимфа - почти прозрачная желтоватая жидкость, находящаяся в полости лимфатических капилляров и сосудов. Образование её обусловлено переходом составных частей плазмы крови из кровеносных капилляров в тканевую жидкость. В образовании лимфы существенное значение имеют взаимоотношение гидростатического и осмотического давления крови и тканевой жидкости, проницаемость стенки кровеносных капилляров и т. д.
Лимфа состоит из жидкой части - лимфоплазмы и форменных элементов. Лимфоплазма отличается от плазмы крови меньшим содержанием белков. В лимфе содержится фибриноген, поэтому она также способна к свертыванию. Главные форменные элементы лимфы - лимфоциты. Состав лимфы в различных сосудах лимфатической системы неодинаков. Различают периферическую лимфу (до лимфоузлов) , промежуточную (после лимфоузлов) и центральную (лимфа грудного и правого лимфатического протоков), наиболее богатая клеточными элементами.
3. Кроветворение (гемоцитопоэз) - многостадийный процесс последовательных клеточных превращений, приводящих к образованию зрелых клеток периферической сосудистой крови.
В постэмбриональный период у животных развитие клеток крови осуществляется в двух специализированных интенсивно обновляющихся тканях - миелоидной и лимфоидной.
В настоящее время наиболее признанной является схема кроветворения, предложенная И.Л. Чертковым и А.И. Воробьевым (1981), в соответствии с которой весь гемоцитопоэз разделен на 6 этапов (рис. 8).
Родоначальником всех клеток крови (по А.А. Максимову) является полипотентная стволовая клетка (колониеобразующая единица в селезенке и КОЕс). Во взрослом организме наибольшее количество стволовых клеток находится в красном костном мозге (на 100000 клеток костного мозга приходится около 50 стволовых), из которого они мигрируют в тимус, селезенку.
Развитие эритроцитов (эритроцитопоэз) в красном костном мозге протекает по схеме: стволовая клетка (СК) - полустволовые клетки (КОЕ - ГЭММ, КОЕ- ГЭ, КОЕ - МГЦЭ) - унипотентные предшественники эритропоэза (БОЕ - Э,КОЕ - Э) - эритробласт - пронормоцит - нормоцит базофильный - нормоцит полихроматофильный - нормоцит оксифильный - ретикулоцит - эритроцит.
Развитие гранулоцитов: стволовая клетка красного костного мозга, полустволовые (КОЕ - ГЭММ, КОЕ - ГМ, КОЕ - ГЭ), унипотентные предшественники (КОЕ - Б, КОЕ - Эо, КОЕ - Гн), которые через стадии распознаваемых клеточных форм превращаются в зрелые сегментоядерные гранулоциты трех разновидностей - нейтрофилы, эозинофилы и базофилы.
Развитие лимфоцитов - один из наиболее сложных процессов дифференцировки стволовых кроветворных клеток.
С участием различных органов поэтапно осуществляется формирование двух тесно связанных при функционировании линий клеток - Т- и В-лимфоцитов.
Развитие кровяных пластинок происходит в красном костном мозге и связано с развитием в нем особых гигантских клеток - мегакариоцитов. Мегакариоцитопоэз состоит из следующих стадий: СК - полустволовые клетки (КОЕ -ГЭММ и КОЕ - МГЦЭ) - унипотентные предшественники, (КОЕ - МГЦ) - мегакариобласт - промегакариоцит - мегакариоцит.
4. На самых ранних этапах онтогенеза клетки крови образуются за пределами зародыша, в мезенхиме желточного мешка, где формируются скопления - кровяные островки. Центральные клетки островков округляются и преобразуются в стволовые кроветворные клетки. Периферические клетки островков растягиваются в полоски, связанные между собой клетки и образуют эндотелиальную выстилку первичных кровеносных сосудов (сосудистая сеть желточного мешка). Часть стволовых клеток превращается в крупные базофильные бластные клетки - первичные кровяные клетки. Большая часть этих клеток, интенсивно размножаясь все сильнее окрашивается кислыми красителями. Это происходит в связи с синтезом и накоплением в цитоплазме гемоглобина, а в ядре конденсированного хроматина. Такие клетки называют первичными эритробластами. В некоторых первичных эритробластах распадается и исчезает ядро. Образующаяся генерация ядерных и безъядерных первичных эритроцитов разнообразна по размерам, однако чаще всего встречаются крупные клетки - мегалобласты и мегалоциты. Мегалобластический тип кроветворения характерен для эмбрионального периода.
Часть первичных кровяных клеток преобразуется в популяцию вторичных эритроцитов, а вне сосудов развивается небольшое количество гранулоцитов - нейтрофилов и эозинофилов, т. е. происходит миелопоэз.
Возникшие в желточном мешке стволовые клетки с кровью переносятся в органы организма. После закладки печени она становится универсальным органом кроветворения (развиваются вторичные эритроциты, зернистые лейкоциты и мегакариоциты). К концу внутриутробного периода кроветворение в печени прекращается.
На 7-8 неделе эмбрионального развития (у крупного рогатого скота) из стволовых клеток в развивающемся тимусе дифференцируются лимфоциты тимуса и мигрирующие из него Т-лимфоциты. Последние заселяют Т-зоны селезенки и лимфатических узлов. В начале своего развития селезенка также является органом, в котором образуются все виды форменных элементов крови.
На последних стадиях эмбрионального развития у животных основные кроветворные функции начинает выполнять красный костный мозг; в нем образуются эритроциты, гранулоциты, кровяные пластинки, часть лимфоцитов (В-л). В постэмбриональный период красный костный мозг становится органом универсального гемопоэза.
Во время эмбрионального эритроцитопоэза идет характерный процесс смены генераций эритроцитов, отличающихся морфологией и типом образующегося гемоглобина. Популяция первичных эритроцитов образует эмбриональный тип гемоглобина (Нв - F). на последующих стадиях эритроциты в печени и селезенке содержат плодный ( фетальный) тип гемоглобина (Нв-Г). В красном костном мозге образуются дефинитивный тип эритроцитов с третьим типом гемоглобина (Нв-А и Нв-А 2). Разные типы гемоглобинов отличаются составом аминокислот в белковой части.
клетка эмбриогенез ткань гистология цитология
Собственно соединительная ткань
1. Рыхлая и плотная соединительная ткань.
2. Соединительная ткань со специальными свойствами: ретикулярная, жировая, пигментная.
1. Широко распространенные в организме животных ткани с сильно развитой в межклеточном веществе системой волокон, благодаря которым эти ткани выполняют разносторонние механические и формообразующие функции - формируют комплекс перегородок, трабекул или прослоек внутри органов, входят в состав многочисленных оболочек, образуют капсулы, связки, фасции, сухожилия.
В зависимости от количественного соотношения между компонентами межклеточного вещества - волокнами и основным веществом и в соответствии с типом волокон различают три вида соединительных тканей: рыхлую соединительную ткань, плотную соединительную ткань и ретикулярную ткань.
Основными клетками, создающими вещества, необходимые для построения волокон в рыхлой и плотной соединительной ткани, являются фибробласты, в ретикулярной ткани - ретикулярные клетки. Рыхлая соединительная ткань отличается особенно большим разнообразием клеточного состава.
Рыхлая соединительная ткань является наиболее распространенной. Она сопровождает все кровеносные и лимфатические сосуды, формирует многочисленные прослойки внутри органов и т. д. Состоит она из разнообразных клеток, основного вещества и системы коллагеновых и эластических волокон. В составе данной ткани различают более оседлые клетки (фибробласты - фиброциты, липоциты), подвижные (гистиоциты - макрофаги, тканевые базофилы, плазмоциты)- рис.9.
Главные функции этой соединительной ткани: трофическая, защитная и пластическая.
Разновидности клеток: Адвентициальные клетки - малодифференцированы, способны к митотическому делению и превращению в фибробласты, миофибробласты и липоциты. Фибробласты - основные клетки, принимающие непосредственное участие в формировании межклеточных структур. В ходе зародышевого развития фибробласты возникают непосредственно из мезенхимных клеток. Различают три разновидности фибробластов: малодифференцированные (функция: синтез и секреция гликозаминогликанов); зрелые (функция: синтез проколлагена, проэластина, ферментных белков и гликозаминогликанов, особенно - синтез белка коллагеновых волокон); миофибробласты, способствующие закрытию раны. Фиброциты утрачивают способность к делению, снижают синтетическую активность. Гистиоциты (макрофаги) относятся к системе мононуклеарных фагоцитов (СМФ). Об этой системе речь будет идти в следующей лекции. Тканевые базофилы (лаброциты, тучные клетки), располагаясь вблизи мелких кровеносных сосудов, они одни из первых клеток реагируют на проникновение антигенов из крови.
Плазмоциды - в функциональном отношении - эффекторные клетки иммунологических реакций гуморального типа. Это высокоспециализированные клетки организма, синтезирующие и выделяющие основную массу разнообразных антител (иммуноглобулинов).
Межклеточное вещество рыхлой соединительной ткани составляет значительную её часть. Представлено оно коллагеновыми и эластическими волокнами и основным (аморфным) веществом.
Аморфное вещество - продукт синтеза клеток соединительной ткани (преимущественно фибробластов) и поступлением веществ из крови, прозрачная, слегка желтоватая, способная менять свою консистенцию, что существенно отражается на его свойствах.
В его состав входят гликозаминогликаны (полисахариды), протеогликаны, гликопротеиды, вода и неорганические соли. Важнейшим химическим высокополимерным веществом в этом комплексе является несульфатированная разновидность гликозаминогликанов - гиалуроновая кислота.
Коллагеновые волокна состоят из фибрилл, образованных молекулами белка тропоколлагена. Последние являются своеобразными мономерами. Образование фибрилл - результат характерной группировки мономеров в продольном и поперечном направлении.
В зависимости от аминокислотного состава и формы объединения цепей в тройную спираль различают четыре основных типа коллагена, имеющих различную локализацию в организме. Коллаген I типа содержится в соединительной ткани кожи, сухожилий и в костях. Коллаген II типа - в гиалиновом и волокнистом хрящах. Коллаген II? типа - в коже зародышей, стенке кровеносных сосудов, связках. Коллаген IV типа - в базальных мембранах.
Выделяют два способа образования коллагеновых волокон: внутриклеточный и внеклеточный синтез.
Эластические волокна - это гомогенные нити, формирующие сеть. Не объединяются в пучки, обладают малой прочностью. Различают более прозрачную аморфную центральную часть, состоящую из белка эластина, и периферическую, состоящую из микрофибрилл гликопротеидной природы, имеющих форму трубочек. Эластические волокна образуются, благодаря синтетической и секреторной функции фибробластов. Считается, что вначале в непосредственной близости от фибробластов образуется каркас из микрофибрилл, а затем усиливается образование аморфной части из предшественника эластина - проэластина. Молекулы проэластина под влиянием ферментов укорачиваются и превращаются в молекулы тропоэластина. Последние при образовании эластина соединяются между собой с помощью десмозина, отсутствующего в других белках. Преобладают эластические волокна в затылочно-шейной связке, брюшной желтой фасции.
Плотная соединительная ткань. Эта ткань характеризуется количественным преобладанием волокон над основным веществом и клетками. В зависимости от взаимного расположения волокон и образованных из низ пучков сетей различают две основные разновидности плотной соединительной ткани: неоформленную (дерма) и оформленную (связки, сухожилия).
2. Ретикулярная ткань состоит из отростчатых ретикулярных клеток и ретикулярных волокон (рис.10). Ретикулярная ткань образует строму кроветворных органов, где в комплексе с макрофагами создает микроокружение, обеспечивающее размножение, дифференциацию и миграцию разных форменных элементов крови.
Ретикулярные клетки развиваются из мезенхимоцитов и имеют сходство с фибробластами, хондробластами и др. Ретикулярные волокна - производные ретикулярных клеток и представляют тонкие ветвящиеся волокна, образующие сеть. В их составе различные по диаметру фибриллы, заключенные в межфибриллярное вещество. Фибриллы состоят из коллагена III типа.
Жировая ткань образуется жировыми клетками (липоцитами). Последние специализированы на синтезе и накоплении в цитоплазме запасных липидов, главным образом, триглицеридов. Липоциты широко распространены в рыхлой соединительной ткани. В эмбриогенезе жировые клетки возникают из клеток мезенхимы.
Предшественниками для образования новых жировых клеток в постэмбриональный период являются адвентициальные клетки, сопровождающие кровеносные капилляры.
Различают две разновидности липоцитов и собственно два типа жировой ткани: белую и бурую. Белая жировая ткань содержится в организме животных неодинаково в зависимости от вида и породы. Её много в жировых депо. Общее количество ее в организме животных различных видов, пород , пола, возраста, упитанности колеблется от 1 до 30% к жировой массе. Жир как источник энергии (1 г жира =39 кДж), депо воды, амортизатор.

Рис. 11. Строение белой жировой ткани (схема по Ю.И.Афанасьеву)
А — адипоциты с удаленным жиром в световом оптическом микроскопе; Б — ультрамикроскопическое строение адипоцитов. 1 — ядро жировой клетки; 2 — крупные капли липидов; 3 — нервные волокна; 4 — гемокапилляры; 5 — митохондрии.

Рис. 12. Строение бурой жировой ткани (схема по Ю.И.Афанасьеву)
А — адипоциты с удаленным жиром в световом оптическом микроскопе; Б — ультрамикроскопическое строение адипоцитов. 1 — ядро адипоцита; 2 — мелко раздробленные липиды; 3 — многочисленные митохондрии; 4 — гемокапилляры; 5 — нервное волокно.
Бурая жировая ткань в значительном количестве имеется у грызунов и животных, впадающих в зимнюю спячку; а также у новорожденных других видов. Клетки, окисляясь, образуют тепло, идущее на терморегуляцию.
Пигментные клетки (пигментоциты) имеют в цитоплазме много темно-коричневых или черных зерен пигмента из группы меланинов.
Иммунная система и клеточные взаимодействия в иммунных реакциях
1. Понятие об антигенах и антителах, их разновидности.
2 Понятие о клеточном и гуморальном иммунитете.
3 Генез и взаимодействие Т- и В-лимфоцитов.
4 Мононуклеарная система макрофагов.
1. В промышленном животноводстве в условиях концентрации и интенсивной эксплуатации поголовья, стрессовых воздействий техногенных и других факторов окружающей среды значительно возрастает роль профилактики заболеваний животных, особенно молодняка, обусловленных воздействием разнообразных агентов заразной и незаразной природы на фоне понижения естественных защитных способностей организма.
В связи с этим большое значение приобретает проблема контроля физиологического и иммунологического состояния животных с целью своевременного повышения их общей и специфической устойчивости (Цымбал А.М., Конаржевский К.Е. и др., 1984).
Иммунитет (immunitatis - освобождение от чего-либо) - это защита организма от всего генетически чужеродного - микробов, вирусов, от чужеродных клеток. или генетически измененных собственных клеток.
Иммунная система объединяет органы и ткани, в которых происходит образование и взаимодействие клеток - иммуноцитов, выполняющих функцию распознавания генетически чужеродных субстанций (антигенов) и осуществляющих специфическую реакцию.
Антигены - это сложные органические вещества, способные при поступлении в организм человека и животных вызывать специфический иммунный ответ. Свойствами антигенов обладают бактерии, вирусы, паразиты, чужеродные клетки, а также продукты жизнедеятельности чужеродных клеток - белки, полисахариды, полипептиды.
Антитела - это сложные белки, находящиеся в иммуноглобулиновой фракции плазмы крови животных, синтезируемые плазматическими клетками под воздействием различных антигенов. Изучено несколько классов иммуноглобулинов (Y, M, A, E, D).
При первой встрече с антигеном (первичный ответ) лимфоциты стимулируются и подвергаются трансформации в бластные формы, которые способны к пролиферации и дифференцировке в иммуноциты. Дифференцировка приводит к появлению двух типов клеток - эффекторных и клеток памяти. Первые непосредственно участвуют в ликвидации чужеродного материала. К эффекторным клеткам относят активированные лимфоциты и плазматические клетки. Клетки памяти - это лимфоциты, возвращающиеся в неактивное состояние, но несущие информацию (память) о встрече с конкретным антигеном. При повторном введении данного антигена они способны обеспечивать быстрый иммунный ответ (вторичный ответ) вследствие усиленной пролиферации лимфоцитов и образования иммуноцитов.
2. В зависимости от механизма уничтожения антигена различают клеточный иммунитет и гуморальный иммунитет.
При клеточном иммунитете эффекторными (двигательными) клетками являются цитотоксические Т-лимфоциты, или лимфоциты-киллеры (убийцы), которые непосредственно учавствуют в уничтожении чужеродных клеток других органов или патологических собственных клеток (например, опухолевых) и выделяют литические вещества.
При гуморальном иммунитете эффекторными клетками являются плазматические клетки, которые синтезируют и выделяют в кровь антитела.
В становлении клеточного и гуморального иммунитета в организме человека и животных большую роль играют клеточные элементы лимфоидной ткани, в частности Т- и В-лимфоциты. Сведения о популяциях этих клеток в крови крупного рогатого скота немногочисленны. По данным Корчана Н.И. (1984), телята рождаются с относительно зрелой системой В-лимфоцитов и недостаточно развитой системой В-лимфоцитов и регуляторными взаимоотношениями между этими клетками. Лишь к 10-15 дню жизни показатели систем этих клеток приближаются к показателям у взрослых животных.
Иммунная система в организме взрослого животного представлена: красным костным мозгом - источником стволовых клеток для иммуноцитов, центральными органами лимфоцитопоэза (тимус), периферическими органами лимфоцитопоэза (селезенка, лимфатические узлы, скопление лимфоидной ткани в органах), лимфоцитами крови и лимфы, а также популяциями лимфоцитов и плазмоцитов, проникающими во все соединительные и эпителиальные ткани. Все органы иммунной системы функционируют как единое целое благодаря нейрогуморальным механизмам регуляции, а также постоянно совершающимся процессам миграции и рециркуляции клеток по кровеносной и лимфатической системам. Главными клетками, осуществляющими контроль и иммунологическую защиту в организме, являются лимфоциты, а также плазматические клетки и макрофаги.
3. Различают две основные разновидности лимфоцитов: В-лимфоциты и Т-лимфоциты. Стволовые клетки и клетки-предшественники В-лимфоцитов образуются в костном мозге. У млекопитающих здесь же происходит дифференцировка В-лимфоцитов, характеризующаяся появлением у клеток иммуноглобулиновых рецепторов. Далее такие дифференцированные В-лимфоциты поступают в периферические лимфоидные органы: селезенку, лимфатические узлы, лимфатические узелки пищеварительного тракта. В этих органах при действии антигенов происходит пролиферация и дальнейшая специализация В-лимфоцитов с образованием эффекторных клеток и В-клеток памяти.
Т-лимфоциты также развиваются из стволовых клеток костно-мозгового происхождения. Последние переносятся с током крови в тимус, превращаются в бласты, которые делятся и дифференцируются в двух направлениях. Одни бласты образуют популяцию лимфоцитов, обладающих специальными рецепторами, воспринимающими чужеродные антигены. Дифференцировка этих клеток происходит под влиянием индуктора дифференцировки, вырабатываемого и выделяемого эпителиальными элементами тимуса. Образующиеся в результате Т-лимфоциты (антигенреактивные лимфоциты) заселяют специальные Т-зоны (тимус-зависимые) в периферических лимфоидных органах. Там под влиянием антигенов они могут подвергаться трансформации в Т-бласты, пролиферировать и дифференцироваться в эффекторные клетки, участвующие в трансплантационном (Т-киллеры) и гуморальном иммунитете (Т-хелперы и Т-супрессоры), а также в Т-клетки памяти. Другая часть потомков Т-бластов дифференцируются с образованием клеток, несущих рецепторы к антигенам собственного организма. Эти клетки подвергаются разрушению.
Таким образом, необходимо различать антигеннезависимую и антигензависимую пролиферацию, дифференцировку и специализацию В- и Т-лимфоцитов.
В случае формирования клеточного иммунитета при действии тканевых антигенов дифференцировка Т-лимфобластов приводит к появлению цитотоксических лимфоцитов (Т-киллеры) и Т-клеток памяти. Цитотоксические лимфоциты способны разрушать чужеродные клетки (клетки-мишени) или посредством выделяемых ими особых веществ-медиаторов (лимфокинов).
При формировании гуморального иммунитета большинство растворимых и других антигенов также оказывают стимулирующее влияние на Т-лимфоциты; при этом формируются Т-хелперы, которые выделяют медиаторы (лимфокины), взаимодействующие с В-лимфоцитами и вызывающие их трансформацию в В-бласты, специализирующиеся в секретирующие антитела плазматической клетки. Пролиферация стимулированных антигеном Т-лимфоцитов приводит также к увеличению числа клеток, которые превращаются в неактивные малые лимфоциты, сохраняющие в течение нескольких лет информацию о данном антигене и поэтому называющиеся Т-клетками памяти.
Т-хелпер обусловливает специализацию В-лимфоцитов в направлении формирования антителобразующих плазмоцитов, которые обеспечивают “гуморальный иммунитет”, вырабатывая и выделяя в кровь иммуноглобулины. Одновременно В-лимфоцит получает антигенную информацию от макрофага, который захватывает антиген, перерабатывает его и передает В-лимфоциту. На поверхности В-лимфоцита расположено большее количество иммуноглобулиновых рецепторов (50-150 тыс).
Таким образом, для обеспечения иммунологических реакций необходима кооперация деятельности трех основных типов клеток: В-лимфоцитов, макрофагов и Т-лимфоцитов (рис.13).
4. Макрофаги играют важную роль как в
естественном, так и в приобретенном
иммунитете организма. Участие макрофагов
в естественном иммунитете проявляется
в их способности к фагоцитозу. Их роль
в приобретенном иммунитете заключается
в пассивной передаче антигена
иммунокомпетентным клеткам (Т- и
В-лимфоцитам), в индукции специфического
ответа на антигены.
Выделяемая макрофагами большая часть переработанного материала антигенов оказывает стимулирующее влияние на пролиферацию и дифференцировку клонов Т- и В-лимфоцитов.
В В-зонах лимфатических узлов и селезенки имеются специализированные макрофаги (дендритные клетки), на поверхности многочисленных отростков которых сохраняются многие антигены, попадающие в организм и передающиеся соответствующим клонам В-лимфоцитов. В Т-зонах лимфатических фолликулов расположены интердигитирующие клетки, влияющие на дифференцировку клонов Т-лимфоцитов.
Таким образом, макрофаги принимают непосредственное участие в кооперативном взаимодействии клеток (Т- и В-лимфоцитов) в иммунных реакциях организма.
Различают два типа миграции клеток иммунной системы: медленный и быстрый. Первый более типичен для В-лимфоцитов, второй - для Т-лимфоцитов. Процессы миграции и рециркуляции клеток иммунной системы обеспечивают поддержание иммунного гомеостаза.
Смотрите также учебное пособие «Методы оценки защитных систем организма млекопитающих» (Кацы Г.Д., Коюда Л.И. - Луганск.-2003.- с.42-68).
Скелетные ткани: хрящевая и костная
1. Развитие, строение и разновидности хрящевой ткани.
2. Развитие, строение и разновидности костной ткани.
1. Хрящевая ткань - специализированный вид соединительной ткани, выполняющий опорную функцию. В эмбриогенезе она развивается из мезенхимы и формирует скелет зародыша, который в последующем в большей части замещается костью. Хрящевая ткань, за исключением суставных поверхностей, покрыта плотной соединительной тканью - надхрящницей, содержащей сосуды, питающие хрящ и его камбиальные ( хондрогенные) клетки.
Хрящ состоит из клеток хондроцитов и межклеточного вещества. В соответствии с характеристикой межклеточного вещества различают три вида хрящей: гиалиновый, эластический и волокнистый.
В процессе эмбрионального развития зародыша мезенхима, интенсивно развиваясь, образует островки плотно прилегающих друг к другу клеток протохондральной ткани. Ее клетки характеризуются высокими значениями ядерно-цитоплазматических отношений, мелкими плотными митохондриями, обилием свободных рибосом, слабым развитием гранулярной ЭПС и т. д. В процессе развития из этих клеток формируется первичная хрящевая (прехондральная) ткань.
По мере накопления межклеточного вещества клетки развивающегося хряща изолируются в отдельных полостях (лакунах) и дифференцируются в зрелые хрящевые клетки - хондроциты.
Дальнейший рост хрящевой ткани обеспечивается продолжающимся делением хондроцитов и формированием между дочерними клетками межклеточного вещества. Образование последнего со временем замедляется. Дочерние клетки, оставаясь в одной лакуне, образуют изогенные группы клеток (Isos -равный, genesis - происхождение).
По мере дифференцировки хрящевой ткани интенсивность размножения клеток падает, ядра пиктонизируются, ядрышковый аппарат редуцируется.
Гиалиновый хрящ. Во взрослом организме гиалиновый хрящ входит в состав ребер, грудины, покрывает суставные поверхности и т. д. (рис.14).
Клетки хряща - хондроциты - различных его зон имеют свои особенности. Так, непосредственно под надхрящницей локализованы незрелые хрящевые клетки - хондробласты. Они овальной формы, цитоплазма богата РНК. В более глубоких зонах хряща хондроциты округляются, образуют характерные “изогенные группы”.
Межклеточное вещество гиалинового хряща содержит до 70% сухого веса фибриллярного белка коллагена и до 30% аморфного вещества, в состав которого входят гликозаминогликаны, протеогликаны, липиды и неколлагеновые белки.
Ориентация волокон межклеточного вещества определяется характерными для каждого хряща закономерностями механической напряженности.
Коллагеновые фибриллы хряща в отличие от коллагеновых волокон других видов соединительной ткани тонкие и не превышают 10 нм в диаметре.
Обмен веществ хряща обеспечивается циркуляцией тканевой жидкости межклеточного вещества, которая составляет до 75% общей массы ткани.
Эластичекий хрящ образует скелет наружного уха, хрящей гортани. В его состав, кроме аморфного вещества и коллагеновых фибрилл, входит плотная сеть эластических волокон. Клетки его идентичны клеткам гиалинового хряща. Они также образуют группы и лишь под надхрящницей лежат одиночно (рис.15).
Волокнистый хрящ локализуется в составе межпозвоночных дисков, в области прикрепления сухожилия к костям. Межклеточное вещество содержит грубые пучки коллагеновых волокон. Клетки хряща образуют изогенные группы, вытянутые в цепочки между пучками коллагеновых волокон (рис.16).
Регенерация хряща обеспечивается надхрящницей, клетки которой сохраняют камбиальность- хондрогенные клетки.
2. Костная ткань, как и другие виды соединительной ткани, развивается из мезенхимы и состоит из клеток и межклеточного вещества. Выполняет функцию опоры, защиты и активно участвует в обмене веществ. В губчатом веществе костей скелета локализован красный костный мозг, где осуществляются процессы кроветворения и дифференцировки клеток иммунной защиты организма. Кость депонирует соли кальция, фосфора и др. В совокупности минеральные вещества составляют 65-70% сухой массы ткани.
Костная ткань содержит четыре различных вида клеток: остеогенные клетки, остеобласты, остеоциты и остеокласты.
Остеогенные клетки - клетки ранней стадии специфической дифференцировки мезенхимы в процессе остеогенеза. Они сохраняют потенцию к митотическому делению. Локализуются эти клетки на поверхности костной ткани: в надкостнице, эндоосте, в гаверсовых каналах и других зонах формирования костной ткани. Размножаясь, они пополняют запас остеобластов.
Остеобласты - клетки, продуцирующие органические элементы межклеточного вещества костной ткани: коллаген, гликозаминогликаны, белки и др.
Остеоциты лежат в особых полостях межклеточного вещества - лакунах, соединенных между собой многочисленными костными канальцами.
Остеокласты - крупные, многоядерные клетки. Они находятся на поверхности костной ткани в местах ее резорбции. Клетки поляризованные. Поверхность, обращенная к резорбируемой ткани, имеет гофрированную каемку за счет тонких ветвящихся отростков.
Межклеточное вещество состоит из коллагеновых волокон и аморфного вещества: гликопротеидов, гликозаминогликанов, белков и неорганических соединений. 97% всего кальция организма сосредоточено в костной ткани.
В соответствии со структурной организацией межклеточного вещества различают грубоволокнистую кость и пластинчатую (рис.17). Грубоволокнистая кость характеризуется значительным диаметром пучков коллагеновых фибрилл и разнообразием их ориентации. Она типична для костей ранней стадии онтогенеза животных. В пластинчатой кости коллагеновые фибриллы не образуют пучков. Располагаясь параллельно, они формируют слои - костные пластинки толщиной 3-7 мкм. В пластинках расположены клеточные полости - лакуны и соединяющие их костные канальца, в которых лежат остеоциты и их отростки. По системе лакун и канальцев циркулирует тканевая жидкость, обеспечивающая обмен веществ в ткани.
В зависимости от положения костных пластинок различают губчатую и компактную костную ткань. В губчатом веществе, в частности в эпифизах трубчатых костей, группы костных пластинок располагаются под разными углами друг к другу. Ячеи губчатого вещества кости содержат красный костный мозг.
В компактном веществе группы костных пластинок 4-15 мкм толщиной плотно прилегают друг к другу. В диафизах формируется три слоя: наружная общая система пластинок, остеогенный слой и внутренняя общая система.
Через наружную общую систему из надкостницы проходят прободающие канальца, несущие в кость кровеносные сосуды и грубые пучки коллагеновых волокон.
В остеогенном слое трубчатой кости каналы остеона, содержащие кровеносные сосуды, нервы, в основном, ориентированы продольно. Система трубкообразных костных пластинок, окружающие эти каналы, - остеоны содержат от4 до 20 пластинок. Остеоны отграничены друг от друга цементной линией основного вещества они являются структурной единицей костной ткани (рис.18).
Внутренняя общая система костных пластинок граничит с эндоостом костной полоси и представлена пластинками, ориентированными параллельно поверхности канала.
Различают два вида остеогенеза: непосредственно из мезенхимы («прямой») и путем замещения костью эмбрионального хряща («непрямой») остеогенез – рис. 19,20.
Первый характерен для развития грубоволокнистой кости черепа и нижней челюсти. Процесс начинается с интенсивного развития соединительной ткани и кровеносных сосудов. Мезенхимные клетки, анастомозируя между собой отростками, образуют сеть. Клетки, оттесненные межклеточным веществом на поверхность, дифференцируются в остеобласты, активно участвующие в остеогенезе. В последующем первичная грубоволокнистая костная ткань замещается пластинчатой костью. Кости туловища, конечностей и др. формируются на месте хрящевой ткани. В трубчатых костях этот процесс начинается в области диафиза формированием под надхрящницей сети перекладин грубоволокнистой кости - костной манжетки. Процесс замещения хряща костной тканью называется энхондральным окостенением.
Одновременно с развитием энхондральной кости со стороны надкостницы идет активный процесс перихондрального остеогенеза, формирующий плотный слой периостальной кости, распространяющийся по всей ее длине до эпифизарной пластинки роста. Периостальная кость представляет компактное вещество кости скелета.
Позднее центры окостенения появляются в эпифизах кости. Костная ткань здесь замещает хрящевую. Последняя сохраняется лишь на суставной поверхности и в эпифизарной пластинке роста, отграничивающей эпифиз от диафиза в течение всего периода роста организма до половой зрелости животного.
Надкостница (периост) состоит из двух слоев: внутренний - содержит коллагеновые и эластические волокна, остеобластыЮ остеокласты и кровеносные сосуды. Наружный - образован плотной соединительной тканью. Она непосредственно связана с сухожилиями мышц.
Эндоост - слой соединительной ткани, выстилающий костно-мозговой канал. Он содержит остеобласты и тонкие пучки коллагеновых волокон, переходящих в ткань костного мозга.
Мышечные ткани
1. Гладкие.
2. Сердечная поперечнополосатая.
3. Скелетные поперечнополосатые.
4. Развитие, рост и регенерация мышечных волокон.
1. Ведущая функция мышечных тканей - обеспечение перемещения в пространстве организма в целом и его частей. Все мышечные ткани составляют морфофункциональную группу, а в зависимости от структуры органелл сокращения ее делят на три группы: гладкие, скелетные поперечно-полосатые и сердечные поперечнополосатые мышечные ткани. Единого источника эмбрионального развития у этих тканей нет. Ими являются мезенхима, миотомы сегментированной мезодермы, висцеральный листок спланхнотома и др.
Гладкие мышечные ткани мезенхимного происхождения. Ткань состоит из миоцитов и соединительнотканного компонента. Гладкий миоцит представляет собой веретеновидную клетку длиной 20-500 мкм, толщиной 5-8 мкм. Ядро палочковидной формы находится в ее центральной части. В клетке много митохондрий.
Каждый миоцит окружен базальной мембраной. В ней есть отверстия, в области которых между соседними миоцитами образуются щелевидные соединения (нексусы), обеспечивающие функциональные взаимодействия миоцитов в ткани. В базальную мембрану вплетены многочисленные ретикулярные фибриллы. Вокруг мышечных клеток ретикулярные, эластические и тонкие коллагеновые волокна образуют трехмерную сеть - эндомизий, которая объединяет соседние миоциты.
Физиологическая регенерация гладкой мышечной ткани проявляется обычно в условиях повышенных функциональных нагрузок преимущественно в форме компенсаторной гипертрофии. Наиболее отчетливо это наблюдается в мышечной оболочке матки во время беременности.
Элементами мышечной ткани эпидермального происхождения являются миоэпителиальные клетки, развивающиеся из эктодермы. Они располагаются в потовых, молочных, слюнных и слезных железах, дифференцируясь одновременно с их секреторными эпителиальными клетками из общих предшественников. Сокращаясь, клетки способствуют выведению секрета железы.
Гладкие мышцы образуют мышечные слои во всех полых и трубчатых органах.
2. Источники развития сердечной поперечнополосатой мышечной ткани - симметричные участки висцерального листка спланхнотома. Большинство ее клеток дифференцируются в кардиомиоциты (сердечные миоциты), остальные - в клетки мезотелия эпикарда. И те и другие имеют общие клетки-предшественники. В ходе гистогенеза дифференцируются несколько видов кардиомиоцитов: сократительные, проводящие, переходные и секреторные.
Строение сократительных кардиомиоцитов. Клетки имеют удлинненную форму (100-150 мкм), близкую к цилиндрической. Их концы соединяются друг с другом вставочными дисками. Последние выполняют не только механическую функцию, но и проводящую, обеспечивают электрическую связь между клетками. Ядро овальной формы, располагается в центральной части клетки. В ней много митохондрий. Они образуют цепочки вокруг специальных органелл - миофибрилл. Последние построены из постоянно существующих упорядоченно расположенных нитей актина и миозина - сократительных белков. Для их закрепления служат особые структуры - телофрагма и мезофрагма, построенные из других белков.
Участок миофибриллы между двумя Z -линиями называется саркомером. А-полосы - анизотропные, микрофиламенты толстые, содержат миозин: I-полосы - изотропные, микрофиламенты тонкие, содержат актин; H-полоса располагается посредине А-полосы (рис.21).
Существует несколько теорий механизма сокращения миоцитов:
1) Под влиянием потенциала действия, который распространяется по цитолемме, ионы кальция освобождаются, поступают к миофибриллам и инициируют сократительный акт, являющийся результатом взаимодействия актиновых и миозиновых микрофиламентов; 2) Наиболее распространенной в настоящее время теорией является модель скользящих нитей (Г. Хаксли, 1954). Мы являемся сторонниками последней.
Особенности строения проводящих кардиомиоцитов. Клетки крупнее рабочих кардиомиоцитов (длина около 100 мкм, а толщина около 50 мкм). Цитоплазма содержит все органеллы общего значения. Миофибриллы немногочисленны и лежат по периферии клетки. Эти кардиомиоциты соединяются в волокна друг с другом не только концами, но и боковыми поверхностями. Основная функция проводящих кардиомиоцитов состоит в том, что они воспринимают управляющие сигналы от пейсмекерных элементов и передают информацию к сократительным кардиомиоцитам (рис.22).
В дефинитивном состоянии сердечная мышечная ткань не сохраняет ни стволовых клеток, ни клеток-предшественников, поэтому, если кардиомиоциты гибнут (инфаркт), то они не восстанавливаются.
3. Источником развития элементов скелетной поперечнополосатой мышечной ткани являются клетки миоциты. Одни из них дифференцируются на месте, другие же мигрируют из миотомов в мезенхиму. Первые участвуют в формировании миосимпласта, вторые дифференцируются в миосателлитоциты.
Основным элементом скелетной мышечной ткани является мышечное волокно, образованное миосимпластом и миосателлитоцитами. Волокно окружено сарколеммой. Поскольку симпласт не клетка, термин “цитоплазма” не применяют, а говорят “саркоплазма” (греч. sarcos - мясо). В саркоплазме у полюсов ядер располагаются органеллы общего значения. Специальные органеллы представлены миофибриллами.
Механизм сокращения волокон такой же, как и в кардиомиоцитах.
Большую роль в деятельности мышечных волокон играют включения, в первую очередь миоглобина и гликогена. Гликоген служит основным источником энергии, необходимой как для совершения мышечной работы, так и для поддержания теплового баланса всего организма.

Рис. 22. Ультрамикроскопическое строение трех видов кардиомиоцитов: проводящих (А), промежуточных (Б) и рабочих (В) (схема по Г.С. Катинасу)
1 — базальная мембрана; 2 — ядра клеток; 3 — миофибриллы; 4 — плазмолемма; 5 — соединение рабочих кардиомиоцитов (вставочный диск); соединения промежуточного кардиомиоцита с рабочим и проводящим кардиомиоцитами; 6 — соединение проводящих кардиомиоцитов; 7 — поперечные трубочки-системы (органеллы общего назначения не показаны).
Миосателлитоциты прилежат к поверхности симпласта так, что их плазмолеммы соприкасаются. С одним симпластом связано значительное количество сателлитоцитов. Каждый миосателлитоцит - одноядерная клетка. Ядро мельче, чем ядро миосимпласта, и более округлое. Митохондрии и эндоплазматическая сеть распределены в цитоплазме равномерно, комплекс Гольджи и клеточный центр расположены рядом с ядром. Миосателлитоциты - камбиальные элементы скелетной мышечной ткани.
Мышца как орган. Между мышечными волокнами находятся тонкие прослойки рыхлой соединительной ткани - эндомизий. Его ретикулярные и коллагеновые волокна переплетаются с волокнами сарколеммы, что способствует объединению усилий при сокращении. Мышечные волокна группируются в пучки, между которыми располагаются более толстые прослойки рыхлой соединительной ткани - перимизий. В нем содержатся также и эластические волокна. Соединительная ткань, окружающая мышцу в целом, называется эпимизием.
Васкуляризация. Артерии, вступающие в мышцу, ветвятся в перимизии. Рядом с ними много тканевых базофилов, регулирующих проницаемость сосудистой стенки. Капилляры располагаются в эндомизии. Венулы и вены лежат в перимизии рядом с артериолами и артериями. Здесь же проходят и лимфососуды.
Иннервация. Нервы, вступающие в мышцу содержат как эфферентные, так и афферентные волокна. Отросток нервной клетки, приносящий эфферентный нервный импульс, проникает через базальную мембрану и ветвится между ней и плазмолеммой симпласта, участвуя в образовании двигательной, или моторной бляшки. Нервный импульс освобождает здесь медиаторы, которые вызывают возбуждение, распространяющееся по плазмолемме симпласта.
Итак, каждое мышечное волокно иннервируется самостоятельно и окружено сетью гемокапилляров. Этот комплекс образует морфофункциональную единицу скелетной мышцы - мион; иногда мионом называют само мышечное волокно, что не соответствует Международной гистологической номенклатуре.
4. Клетки, из которых в эмбриогенезе образуются поперечнополосатые мышечные волокна, называются миобластами. После ряда делений эти одноядерные клетки, не содержащие миофибрилл, начинают сливаться между собой, формируя удлиненные многоядерные цилиндрические образования - микротрубочки, в которых в свое время появляются миофибриллы и другие органеллы, характерные для поперечнополосатых мышечных волокон. У млекопитающих большинство этих волокон образуется еще до рождения. В период постнатального роста мышцы должны становиться длиннее и толще для того, чтобы сохранить соразмерность с растущим скелетом. Их окончательная величина зависит от выпадающей на их долю работы. После первого года жизни дальнейший рост мышц всецело обусловлен утолщением отдельных волокон, т. е. представляет собой гипертрофию (гипер - над, сверх и трофи - питание), а не увеличением их числа, что называлось бы гиперплазией (от плазис - образование).
Таким образом, поперечнополосатые мышечные волокна растут в толщину путем увеличения числа содержащихся в них миофибрилл ( и других органелл).
Мышечные волокна удлиняются в результате слияния с клетками-сателлитами. Кроме того, в постнатальном периоде возможно удлинение миофибрилл путем пристройки к их концам новых саркомеров.
Регенерация. Клетки-сателлиты не только обеспечивают один из механизмов роста поперечнополосатых мышечных волокон, но и остаются в течение всей жизни потенциальным источником новых миобластов, слияние которых может приводить к образованию совершенно новых мышечных волокон. Клетки-сателлиты способны делиться и давать начало миобластам после мышечной травмы и при некоторых дистрофических состояниях, когда наблюдаются попытки регенерации новых волокон. Однако даже незначительные дефекты мышечной ткани после тяжелых травм заполняются фиброзной тканью, образуемой фибробластами.
Рост и регенерация гладких мышц. Подобно другим типам мышц, гладкие мышцы отвечают на повышенные функциональные требования компенсаторной гипертрофией, но это не единственная возможная реакция. Например, во время беременности увеличиваются не только размеры гладких мышечных клеток в стенке матки (гипертрофия), но и их число (гиперплазия).
У животных при беременности или после введения гормонов в мышечных клетках матки часто можно видеть фигуры митоза; поэтому общепризнано, что гладкие мышечные клетки сохраняют способность к митотическому делению.
Нервная ткань
1. Развитие ткани.
2. Классификация нервных клеток.
3. Нейроглия, её разновидность.
4. Синапсы, волокна, нервные окончания.
1. Нервная ткань - специализированная ткань, которая формирует основную интегрирующую систему организма - нервную систему. Основная функция - проводимость.
Нервная ткань состоит из нервных клеток - нейронов, выполняющих функцию нервного возбуждения и проведения нервного импульса, и нейроглии, обеспечивающей опорную, трофическую и защитную функции.
Нервная ткань развивается из дорсального утолщения эктодермы - нервной пластинки, которая в процессе развития дифференцируется в нервную трубку, нейральные гребни (валики) и нейральные плакоды.
В последующие периоды эмбриогенеза из нервной трубки образуется головной и спинной мозг. Нейральный гребень формирует чувствительные ганглии, ганглии симпатической нервной системы, меланоциты кожи и т. д. Нейральные плакоды участвуют в формировании органов обоняния, слуха, чувствительных ганглиев.
Нервная трубка состоит из одного слоя призматических клеток. Последние, размножаясь, образуют три слоя: внутренний - эпендимный, средний - мантийный и наружный - краевую вуаль.
В последующем клетки внутреннего слоя продуцируют эпендимные клетки, выстилающие центральный канал спинного мозга. Клетки мантийного слоя дифференцируются на нейробласты, превращающиеся далее в нейроны и спонгиобласты, дающие начало различным видам нейроглии (астроцитам, олигодендроцитам).
2. Нервные клетки (нейроциты, нейроны) различных отделов нервной системы характеризуются разнообразием формы, размеров и функционального значения. В соответствии с функцией нервные клетки делятся на рецепторные (афферентные), ассоциативные и эффекторные (эфферентные).
При большом разнообразии формы нервных клеток общим морфологическим признаком является наличие отростков, обеспечивающих их связь в составе рефлекторных дуг. Длина отростков различна и колеблется от нескольких микрон до 1-1,5 м.
Отростки нервных клеток по функциональному значению делят на два вида. Одни принимают нервное возбуждение и проводят его к перикариону нейрона. Они получили название дендритов. Другой вид отростков проводит импульс от тела клетки и передает его на другой нейроцит или на аксон (axos - ось), или нейрит. Все нервные клетки имеют только один нейрит.
По количеству отростков нервные клетки делят на униполярные - с одним отростком, биполярные и мультиполярные (рис.23).
Ядра нервных клеток крупные, округлые или слегка овальные, располагаются в центре перикариона.
Цитоплазма клеток характеризуется обилием различных органелл, нейрофибрилл, хроматофильного вещества. Поверхность клетки покрыта плазмолеммой, которая характеризуется возбудимостью и способностью проводить возбуждение.

Рис. 23. Типы нервных клеток (схема по Т.Н. Радостиной, Л.С. Румянцевой)
А – униполярный нейрон; Б - псевдоуниполярный нейрон; В – биполярный нейрон; Г – мультиполярный нейрон.
Нейрофибриллы - совокупность волокон, структур цитоплазмы, формирующие в перикарионе густое сплетение.
Хроматофильное (базофильное) вещество выявляется в перикарионе нефроцитов и в их дендритах, но отсутствует в аксонах.
Эпендимоциты выстилают полости центральной нервной системы: желудочки головного мозга и спинно-мозговой канал. Клетки, обращенные в полость нервной трубки, содержат реснички. Их противоположные полюса переходят в длинные отростки, которые поддерживают остов тканей нервной трубки. Эпендимоциты участвуют в секреторной функции, выделяя в кровь различные активные вещества.
Астроциты бывают протоплазматические (коротколучистые) и волокнистые (длиннолучистые). Первые локализованы в сером веществе ЦНС ( центральной нервной системы). Они участвуют в обмене веществ нервной ткани и выполняют разграничительную функцию.
Волокнистые астроциты характерны для белого вещества ЦНС. Они образуют поддерживающий аппарат ЦНС.
Олигодендроциты - большая группа клеток ЦНС и ПНС (периферическая нервная система). Они окружают тела нейронов, входят в состав оболочек нервных волокон и нервных окончаний, участвуют в их обмене веществ.
Микроглия (глиальные макрофаги) - специализированная система макрофагов, выполняющая защитную функцию. Они развиваются из мезенхимы, способны к амебоидному перемещению. Они характерны для белого и серого вещества ЦНС.
4. Отростки нервных клеток в совокупности с покрывающими их клетка2ми нейроглии образуют нервные волокна. Расположенные в них отростки нервных клеток называют осевыми цилиндрами, а покрывающие их клетки олигодендроглии - нейролеммоцитами (шванновские клетки).
Различают миелиновые и безмиелиновые нервные волокна.
Безмиелиновые (безмякотные) нервные волокна характерны для вегетативной нервной системы. Леммоциты плотно прилегают друг к другу, образуя непрерывные тяжи. В волокне содержится несколько осевых цилиндров, т. е. отростков различных нервных клеток. Плазмолемма образует глубокие складки, формирующие сдвоенную мембрану - мезаксон, на котором подвешен осевой цилиндр. При световой микроскопии эти структуры не выявляются, что создает впечатление погружения осевых цилиндров непосредственно в цитоплазму глиальных клеток.
Миелиновые (мякотные) нервные волокна. Их диаметр колеблется от 1 до 20 мкм. Они содержат один осевой цилиндр - дендрит или нейрит нервной клетки, покрытой оболочкой, образованной леммоцитами. В оболочке волокна различают два слоя: внутренний - миелиновый, более толстый и наружный - тонкий, содержащий цитоплазму и ядра леммоцитов.
На границе двух леммоцитов оболочка миелинового волокна истончается, образуется сужение волокна - узловой перехват (перехват Ранвье). Участок нервного волокна между двумя перехватами называется межузловым сегментом. Его оболочка соответствует одному леммоциту.
Нервные окончания различны по их функциональному значению. Различают три вида нервных окончаний: эффекторные, рецепторные и концевые аппараты.
Эффекторные нервные окончания - к их числу относят двигательные нервные окончания поперечнополосатых и гладких мышц и секреторные окончания железистых органов.
Двигательные нервные окончания поперечнополосатых скелетных мышц - моторные бляшки - комплекс взаимосвязанных структур нервной и мышечной тканей.
Чувствительные нервные окончания (рецепторы) - специализированные концевые образования дендритов чувствительных нейронов. Различают две большие группы рецепторов: экстерорецепторы и интерорецепторы. Чувствительные окончания делятся на механорецепторы, хеморецепторы, терморецепторы и др. Они подразделяются на свободные нервные окончания и несвободные. Последние покрыты соединительнотканной капсулой и называются инкапсулированными. К этой группе относят пластинчатые тельца (тельца Фатера-Пачини), осязательные тельца (тельца Мейснера) и др.
Пластинчатые тельца характерны для глубоких слоев кожи и внутренних органов. Осязательные тельца также образованы клетками глии.
Синапсы - специализированный контакт двух нейронов, обеспечивающий одностороннее проведение нервного возбуждения. Морфологически в синапсе различают пресинаптический и постсинаптический полюсы, а между ними щель. Встречаются синапсы с химической и электрической передачей.
По месту контакта различают синапсы: аксосоматические, аксодендрические и аксоаксональные.
Пресинаптический полюс синапса характеризуется наличием синаптических пузырьков, содержащих медиатор (ацетилхолин или норадреналин).
Нервная система представлена чувствительными и двигательными клетками, объединенными межнейрональными синапсами в функционально активные образования - рефлекторные дуги. Простая рефлекторная дуга состоит из двух нейронов - чувствительного и двигательного.
Рефлекторные дуги высших позвоночных животных содержат в своем составе ещё значительное количество ассоциативных нейронов, расположенных между чувствительными и двигательными нейронами.
Нерв - это пучок волокон, окруженных плотной оболочкой периневрия. Мелкие нервы состоят только из одного пучка, окруженного эндоневрием. Число и диаметр нервных волокон в пучке весьма непостоянны. В дистальных участках некоторых нервов имеется больше волокон, чем в более проксимальных. Это объясняется ветвлением волокон.
Кровоснабжение нервов. Нервы обильно снабжены сосудами, образующими много анастомозов. Имеются эпиневральные, межпучковые, периневральные и внутрипучковые артерии и артериолы. Эндоневрий содержит сеть капилляров.
Литература
1. Александровская О.В., Радостина Т.Н., Козлов Н.А. Цитология, гистология и эм бриология.-М:Агропромиздат, 1987.- 448 с.
2. Афанасьев Ю.И., Юрина Н.А. Гистология.- М: Медицина, 1991.- 744 с.
3. Вракин В.Ф., Сидорова М.В. Морфология сельскохозяйственных животных. - М: Агропромиздат, 1991.- 528 с.
4. Глаголев П.А., Ипполитова В.И. Анатомия сельскохозяйственных животных с основами гистологии и эмбриологии.- М: Колос, 1977.- 480 с.
5. Хэм А., Кормак Д. Гистология. -М: Мир, 1982.-Т 1-5.
6. Серавин Л.Н. Происхождение эукариотической клетки //Цитология.-1986/-Т. 28.-№ 6-8.
7. Серавин Л.Н. Основные этапы развития клеточной теории и место клетки среди живых систем //Цитология.-1991.-Т.33.-№ 12/-C. 3-27.