Біофізика рецепції органів зору та слуху
Размещено на http://www.
Біофізика рецепції органів зору та слуху
Біофізичні основи зору
Світло, що потрапило в око, фокусується за допомогою хрусталика на шар світлочутливих кліток сітківки – паличках і колбочках. Палички розташовуються по всій поверхні напівсферичної сітківки і відповідають за чорно-білий чи сутінковий зір, їх приблизно 125 млн. Колбочок значно менше (близько 6,5 млн.), вони в основному сконцентровані в центральній частині сітківки і відповідають при досить яскравому висвітленні за сприйняття кольору.
Будівля сітківки людини і хребетних тварин може показатися парадоксальним, тому що світлочутливі клітки знаходяться в задньому шарі сітківки. Перш ніж світло потрапить на них, воно повинно пройти через кілька шарів нервових кліток. Мало того, самі палички і колбочки орієнтовані до світла своїм внутрішнім сегментом, що не містить зорового пігменту. Однак така організація не знижує істотно чутливості ока до світла, тому що нервові клітки і внутрішні сегменти самих світлочутливих кліток прозорі для видимого світла.
Абсолютний поріг чутливості зору людини (тобто мінімальна величина яскравості світлової плями, що виявляється ще, при відсутності світлового тла й в умовах повної темнової адаптації) дорівнює
2,1 – 5,7 10 -17
Дж на поверхні роговиці ока, що відповідає
58–148 квантам синьо-зеленого світла. З
них тільки 5–15 квантів поглинаються
молекулами зорового пігменту в паличках
сітківки, а інші «губляться».
10 -17
Дж на поверхні роговиці ока, що відповідає
58–148 квантам синьо-зеленого світла. З
них тільки 5–15 квантів поглинаються
молекулами зорового пігменту в паличках
сітківки, а інші «губляться».
Близько 4% світла відбивається від роговиці, близько 50% поглинається оптичними середовищами ока. 80–85% світла, що дійшло до сітківки, проходить крізь неї і поглинається в клітках пігментного епітелію. Завдяки пігментному епітелію, що підстилає сітківку, різко зменшується кількість відбитого і розсіяного від задньої стінки ока світла і тим самим поліпшується гострота зору. Якщо пляма що спостерігається тим, що підлягає обстеженню знаходиться на освітленому тлі, то мінімальну різницю, що ще виявляється, інтенсивності плями і тла називають різницевим порогом. Відношення різницевого порога до яскравості тла називають диференціальним порогом. Диференціальний поріг, відповідно до закону Вебера – Фехнера, при зміні яскравості тла залишається постійним. Це означає, що величина різницевого порога росте прямо пропорційно яскравості світлового тла.
Розглянемо будівлю і функції паличок (див. рис. 1). Зоровий пігмент паличок родопсин зосереджений у зовнішніх сегментах паличок, де він убудований у зорові диски. Диски являють собою замкнуті бімолекулярні ліпідні мембрани, що нагадують собою розплющені повітряні кулі, покладені в стопку. Зовнішній сегмент з'єднаний із внутрішнім сегментом тонкою сполучною ніжкою. В внутрішньому сегменті поруч з ніжкою зосереджена велика кількість мітохондрій, у ньому ж розташовується ядро клітки. Наприкінці внутрішнього сегмента, зверненого до світла, знаходиться синаптичний контакт із нервовим волокном.
Молекули родопсину пронизують ліпідний шар мембран зорових дисків. Через дуже низьку в'язкість ліпідного біслоя молекули пігменту можуть робити швидку латеральну дифузію, а після поглинання кванта світла перетерплювати значні конформаційні перебудови. Низька в'язкість ліпідного шару обумовлена високим змістом поліненасичених жирних кислот.
Зоровий пігмент родопсин – складний білок. Він складається з білка опсина і хромофорної групи – чи альдегіду просто ретиналя. У принципі ретиналь може мати кілька просторових ізомерів, але з опсином зв'язується тільки 11-цис-ретиналь.
Цис-транс-фотоізомеризація
ретиналя.
Обидві найбільш важливі конформації
хромофора (11 – цис- і цілком –
транс-ретиналь) зображені на рис. 1.
Молекула ретиналя містить 6 подвійних
сполучених зв'язків, а тому що обертання
навколо подвійного зв'язку неможливо,
то молекула хромофора плоска. Поглинання
світла ретиналем приводить до
 * –
електронному переходу. В органічних
молекулах
* –
електронному переходу. В органічних
молекулах
 орбіталі (незбуджений стан) єднальні,
тобто утворення подвійного зв'язку
супроводжується виділенням енергії.
Навпроти
орбіталі (незбуджений стан) єднальні,
тобто утворення подвійного зв'язку
супроводжується виділенням енергії.
Навпроти * –
орбіталі (електронно-збуджений стан) є
що розпушують, тобто їхнє перекривання
вимагає витрати енергії. В основному
стані
– орбитали перекриваються і молекула
ретиналя плоска.
* –
орбіталі (електронно-збуджений стан) є
що розпушують, тобто їхнє перекривання
вимагає витрати енергії. В основному
стані
– орбитали перекриваються і молекула
ретиналя плоска.
Після поглинання
фотона
-зв'язок
розривається і дві частини молекули
ретиналя повертаються навколо – зв'язка (рис. 2 і 3). Повернення
молекули в основний стан відбувається
в одній із двох крапок виродження, коли
енергії основного і збудженого станів
рівні (см. рис. 2), після чого молекула
знову стає плоскою. Очевидно, що з такого
стану можливий перехід збудженої
молекули як у цис-, так і в трансформу.
При тривалому опроміненні кожної з форм
у системі установлюються визначені
концентрації обох ізомерів. Іншими
словами, фотореакція цис-транс-ізомеризації
ретиналя фотооборотня, що має два
важливих наслідки. По-перше, під дією
світла не відбувається повного переходу
11-цис- у трансформу, а по-друге, реакції
транс-цис-ізомеризації відіграють
важливу роль не тільки в сприйнятті
світла, але й у регенерації зорового
пігменту (див. нижче).
– зв'язка (рис. 2 і 3). Повернення
молекули в основний стан відбувається
в одній із двох крапок виродження, коли
енергії основного і збудженого станів
рівні (см. рис. 2), після чого молекула
знову стає плоскою. Очевидно, що з такого
стану можливий перехід збудженої
молекули як у цис-, так і в трансформу.
При тривалому опроміненні кожної з форм
у системі установлюються визначені
концентрації обох ізомерів. Іншими
словами, фотореакція цис-транс-ізомеризації
ретиналя фотооборотня, що має два
важливих наслідки. По-перше, під дією
світла не відбувається повного переходу
11-цис- у трансформу, а по-друге, реакції
транс-цис-ізомеризації відіграють
важливу роль не тільки в сприйнятті
світла, але й у регенерації зорового
пігменту (див. нижче).
Фотоперетворення родопсину. 11-Цис-ретиналь точно вписується в центр зв'язування молекули опсина. Після фотохімічного утворення трансу-форми стерична відповідність порушується, що приводить до цілої серії конформаційних перебудов у молекулі білка, що супроводжуються змінами в спектрі поглинання зорового пігменту. Такі перебудови родопсину вперше досліджені Уолдом, що получили Нобелівську премію в 1966 р. У досвідах Уолда родопсин виділяли в темряві, заморожували в рідкому азоті при -196° і опромінювали світлом. При наступному нагріванні зразка до визначених температур спостерігалися зміни спектрів поглинання через послідовні переходи родопсину в його похідні. Останні одержали спеціальні назви (рис. 4). На цьому ж рисунку приведені обмірювані методом імпульсного фотолізу часи конформаційних перетворень різних форм пігменту при кімнатній температурі. При цьому тільки перехід родопсин – батородопсин відбувається під дією світла, а інші стадії – у темряві.
Кожен конформер містить ретиналь у трансі-формі і під дією квантів світла може регенерувати безпосередньо в родопсин. Імовірність такої регенерації зростає зі збільшенням часу життя конформера, тому що збільшується імовірність зіткнення з ним кванта світла. Якщо родопсин освітити коротким спалахом світла і далі помістити в темряву, то можливість світлової регенерації родопсину усувається. У цьому випадку приблизно через 100 с відбувається розпад родопсину на опсин і транс-ретиналь. У такій ситуації регенерація родопсину теж відбудеться, але вже при участі ферментів.
Фермент ретинальізомераза переводить транс-ретиналь у 11-цис-ретиналь, після чого відбувається його приєднання до опсину з утворенням родопсину.
Рецепторні потенціали. Яким образом фотоперетворення родопсину приводять до електричної відповіді рецепторної клітки? Відповідь на це питання дозволяють дати дані, отримані методами відведення потенціалів, і зведення по конформаційним перетвореннях родопсину, представлені вище. Реєстрація електроретинограм (екстраклітинне відведення потенціалів від сітківки) дозволила одержати рецепторні потенціали, зображені на рис. 5. Пізніше аналогічні потенціали безпосередньо на цитоплазматичній мембрані паличок і колбочек були отримані мікроелектродними методами. Відразу після висвітлення палички коротким спалахом світла спостерігається триваючий приблизно 1 мс ранній рецепторний потенціал (РРП), амплітуда якого росте зі збільшенням інтенсивності спалаху, але не перевищує 5 мВ. Потім, приблизно через 1 мс, розвивається пізній рецепторний потенціал (ПРП). На відміну від всіх інших відомих кліток на цитоплазматичній мембрані зовнішніх сегментів паличок потенціал має знак «плюс» усередині і знак «мінус» зовні.
Природа РРП і ПРП зовсім різна. Очевидно, РРП зв'язаний з переміщенням молекул родопсину під час конфірмаційних перебудов, викликаних висвітленням. На його молекулі є фіксовані заряди, положення яких відносно біслоя ліпідів після поглинання квантів світла міняється, що і є причиною РРП. При цьому за ФРП відповідальні конформаційні перетворення родопсину до стадії метародопсин II, тобто саме вони відбуваються за час, порівнянний із тривалістю РРП (див. рис. 4). З моменту утворення метародопсина II починаються процеси зовсім іншої природи, у яких вирішальну роль грає рух іонів.
Особливістю зовнішніх сегментів паличок сітківки є те, що в спокої (у темряві) потенціал на їх цитоплазматичній мембрані має натрієву природу, на відміну від потенціалів спокою нервових і багатьох інших кліток. Це зв'язано з тим, що в стані спокою коефіцієнт проникності для натрію значно перевищує значення таких коефіцієнтів для інших іонів. Завдяки цьому по електрохімічному градієнті усередину рухається постійний потік іонів натрію. Натрій, накопичується в цитоплазмі зовнішнього сегмента, а потім через ніжку, що з'єднує його з внутрішнім сегментом, пасивно дифундує у внутрішній сегмент. Тут іони натрію попадають у цілу енергетичну фабрику (скупчення мітохондрій зображене на рис. 1.), де, використовуючи АТФ, що поставляється мітохондріями, працює фермент АТФ-аза, який активно викачує натрій у зовнішнє середовище. Коли з родопсину під дією світла утвориться метародопсин II, проникність цитоплазматичної мембрани для натрію різко знижується, а для інших іонів не міняється. У цих умовах на перше місце виходить проникність для калію, потенціал на мембрані стає калієвої природи і полярність його міняється.
По останнім даним, інформація про фотовицвітання родопсину в дисках передається натрієвим каналам у цитоплазматичній мембрані за допомогою медіаторів. Роль таких медіаторів приділяється іонам кальцію і циклічним нуклеотидам (цГМФ і в меншому ступені цАМФ).
Колірний зір. При слабкому світлі максимум чутливості ока людини розташований близько 500 нм, що відповідає максимуму поглинання родопсину, що міститься в паличках. На яскравому світлі максимум чутливості зміщається до 550 нм, що відповідає максимуму спектра поглинання пігменту в найбільш розповсюдженому типі колбочок. Пігмент колбочок містить той же самий 11-цис-ретиналь, як і родопсин, але білкова частина пігменту відрізняється, тому пігменти колбочок звуться йодопсинами.
Спектри поглинання зорових пігментів вимірюють трьома способами: 1) зорові пігменти екстрагують із сітківки м'якими детергентами і потім вимірюють сумарний спектр поглинання всіх пігментів; 2) проводиться мікроспектрофотометрія окремих фоторецепторних кліток in situ; 3) у інтактних тварин можна вимірити спектр відображення світла від задньої стінки ока до і після засвітки могутнім спалахом яскравого монохроматичного світла; остання приводить до переважного вицвітання тієї форми пігменту, що поглинає світло даної довжини хвилі. Дані різних методів доповнюють один одного.
При вимірі спектрів поглинання окремих колбочок виявилося, що кожна колбочка містить тільки один вид йодопсина, а типів колбочок всього три. Йодопсини людини мають максимуми поглинання при 445, 535 і 570 нм. Поглинання світла йодопсинами приводить до появи РРП у колбочках. Для виміру спектрів дії РРП у колбочки сітківки вводять мікроелектроди і реєструють РРП, які викликаються короткими яскравими спалахами монохроматичного світла. У сітківці коропа таким способом виявлено три типи колбочок з максимумами спектрів дії (а отже, і спектрів поглинання пігментів) при 462, 529 і 611 нм.
При деяких генетичних захворюваннях порушується синтез білків-йодопсинів, у результаті чого не утвориться той чи інший пігмент колірного зору. Людина втрачає здатність розрізняти кольори. Ця хвороба називається дальтонізмом.
Біофізика слуху
Розглянемо послідовність подій при сприйнятті звуку. Звукова хвиля, пройшовши на зовнішнє вухо, наштовхується на туго натягнуту барабанну перетинку, приводячи її в рух. Барабанна перетинка зв'язана із системою слухових кісточок середнього вуха, що передають звукові коливання на внутрішнє вухо – равлика. Слухові кісточки надають руху овальному вікну, що відокремлює перилімфу внутрішнього вуха від повітряного простору середнього вуха. Рух рідини у вестибулярному і базилярному каналах внутрішнього вуха змушує коливатися базилярну мембрану, за частотою і силою звуку. Рух базилярної мембрани стимулює рецепторні клітки, розташовані в кортієвому органі, у результаті з'являються потенціали дії, передані звуковими слуховими нервами в кору головного мозку.
зір слух звук сприйняття
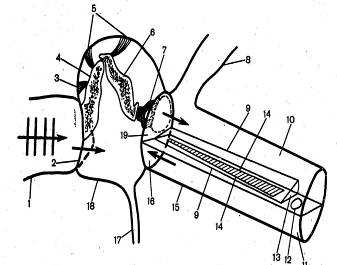
Рис. 6. Схема будівлі органа слуху людини.
1-зовнішній слуховой прохід; 2 – барабанна перетинка; 3 – м'яз, що натягає барабанну перетинку; 4 – молоточек; 5 – зв'язування; 6 – ковадло; 7 – стремінце, що давить на овальне вікно; 8 – вестибулярна частина внутрішнього вуха; 9 – мембрана Рейснера; 10,11 – вестибулярний і барабанний канали (відповідно), заповнені перелімфою; 12 – геликотерма; 13 – равликовий канал, заповнений эндолимфой; 14 – базилярная мембрана; 15 – равлик (розгорнутий); 16 – кругле вікно внутрішнього вуха; 17 – евстахиева труба; 18 – середнє вухо; 19 – овальне вікно внутрішнього вуха.
Розглянемо деякі поняття, використовувані в биоакустике. Інтенсивністю звуку називають кількість енергії, що проходить через одиницю площі поверхні, перпендикулярно до напрямку поширення звукової хвилі.
При рішенні практичних задач простіше вимірити не енергію коливань, а звуковий тиск, за допомогою мікрофона і вило в Паскалях. Звуковий тиск лінійно зв'язаний з амплітудою звукових коливань, і тому його квадрат пропорційний енергії звуку.

Рис. 7. Криві рівної голосності. Нижня крива відповідає порогові слуху чистого тону, верхня – порогові тактильного відчуття.
Поріг чутності чистого тону при частоті 1 кгц прийнято вважати рівним 2 • 10-5 Па, хоча можуть спостерігатися багаторазові індивідуальні коливання порога чутності (у межах 30 дБ). Крім того, поріг чутності значно міняється з частотою (мал. 7). Може також сильно розрізнятися і найвища сприймана частота (від 8 до 25 кгц).
За законом Вебера – Фехнера відчуття голосності [так само як і зорове відчуття] зв'язане з його фізичним роздратуванням, що створює, логарифмічною залежністю. Це стимулювало введення одиниць голосності, що виражаються в децибелах. Тоді нульова крапка шкали голосності відповідає 0 дб при 1 кгц. При інших частотах поріг слуху відрізняється від 0 дб (див. мал. 7).
Зовнішнє вухо людини складається з вушної раковини і зовнішнього слухового проходу, що має довжину 2,7 див і напоминающего органну трубу, закриту з одного кінця барабанною перетинкою. Резонанс спостерігається, якщо довжина резонатора складає 1/4 довжини хвилі. Зовнішній слуховой прохід має тому резонансну частоту близько 3 кгц, що відповідає максимальної чутливості слуху (див. мал. 7). Таким чином, просліджується зв'язок між геометрією зовнішнього вуха і максимальною чутливістю слуху в області 1- 4 кгц.
Середнє вухо є пристроєм, призначеним для трансформації звукових коливань повітря в звукові коливання рідкого середовища внутрішнього вуха, тобто середовища, що має значно велику інерцію, чим повітря. Щоб надати руху інерційній рідині, потрібно одержати виграш у тиску. Це досягається за рахунок двох факторів. По-перше, кісточки середнього вуха (молоточек, ковадло і стремінце) працюють як важіль, що забезпечує виграш у силі в 1,3 рази. По-друге, площа барабанної перетинки людини (0,7 див2) значно більше площі овального вікна внутрішнього вуха (0,032 див2) тому обидві мембрани і єднальні їхні кісточки виконують функцію трансформатора тиску. Очевидно, що сила, що діє на молоточек і обумовлена звуковою хвилею, дорівнює добутку тиску на перетинку на площу контакту перетинки з молоточком (приблизно 5,5 •10-5 м2), відкіля Fc = 5,5 • 10-5 • р>0>. Сила, прикладена до стремінця під час відсутності тертя, дорівнює: F>с> = 1,3 • F>м>. Тиск р>00>, що робиться стремінцем на овальне вікно при площі контакту 3,2 •10-6 м2, може бути знайдене зі співвідношення: р00 = Fc/3,2 • 10-6. Тепер легко обчислити посилення тиску під час відсутності сил тертя: р>00>/р>0> = 22. Бекеші експериментально вимірив цю величину й одержав посилення тиску в 17 разів (чи на 25 дб).
Іншою важливою функцією середнього вуха є захист внутрішнього вуха від надмірних механічних навантажень при впливі дуже голосних звуків. Це досягається за рахунок зміни характеру коливань стремінця в залежності від сили звуку. При граничних значеннях звукового тиску стремінце коливається як поршень. Абсолютні значення зсуву дуже малі і повторюють руху барабанної перетинки. Якщо силу звуку збільшити до 130 дб, то з'являється відчуття болю, а при 155–160 дб – барабанна перетинка руйнується.
Якщо при низьких значеннях звукового тиску поршнеобразние зсуву стремінця практично повторюють руху барабанної перетинки, то при зростанні сили звуку характер коливань стремінця міняється. При середній силі звуку стремінце починає робити коливальні рухи навколо вертикальної осі в одного кінця овального вікна (відкриваподібно і дверей, що закривається,). А при дуже голосному звуці стремінце починає робити обертальні рухи навколо горизонтальної осі овального вікна, так що, коли один кінець стремінця вдавлюється в овальне вікно, іншої рухається в протилежну сторону. Таким чином, запобігаються сильні рухи рідини у внутрішнім вусі. Така зміна характеру рухів стремінця має вирішальне значення для захисту внутрішнього вуха від механічних ушкоджень при різких раптових звуках, наприклад, вибухах, коли не встигає здійснитися будь-який рефлекторний механізм захисту.
Внутрішнє вухо на відміну від зовнішнього і середніх заповнено рідиною. Для слуху важлива тільки равликова частина внутрішнього вуха, що має форму спіралі й утворить у чоловік два з половиною витка (на мал. 6 зображена в розгорнутому виді). Довжина розгорнутого равлика близько 35 мм, а її обсяг – близько 100 мкл (тобто дорівнює обсягу двох крапель води). У равлику розташовуються три рівнобіжних наповнених рідини каналу. Вестибулярний і барабанний канали заповнені перилимфой і з'єднані у вершині равлика маленьким отвором – геликотремой. Ці два канали відділені друг від друга не сполученим з ними равликовим каналом, заповненим эндолимфой і відділеним від вестибулярного каналу дуже тонкою мембраною Рейснера, а від барабанного каналу – базилярной мембраною. На базилярной мембрані – кортиев орган (на мал. 6 не показаний), що містить рецепторні клітки і нервові закінчення.
Характер коливань базилярной мембрани залежить від частоти. При дуже низьких частотах хвилі тиску, передані стремінцем від барабанної перетинки, змушують перилимфу рухатися вперед та назад через геликотрему, викликаючи коливання круглого вікна (див. мал. 6). Такі низькочастотні коливання майже не надають руху базилярную мембрані. При більш високих частотах, наприклад 30 Гц, хвилі тиску через инерционности рідини прагнуть поширитися прямо через базилярную мембрану, приводячи її в рух. Базилярная мембрана дуже неоднорідна по довжині. Від овального вікна до вершини равлика вона уширяется і товщає (ширина росте від 0,04 до 0,5 мм). Поруч зі стремінцем вона вже легше і має приблизно в 100 разів більше значення модуля пружності, чим у вершини. Завдяки неоднорідним механічним властивостям базилярной мембрани хвилі різної частоти надають руху різним її ділянкам. Низькі частоти (менш 100 Гц) викликають коливання найбільш масивної частини мембрани біля геликотреми. Високі частоти (8000 і більш Гц), навпаки, надають руху ділянці мембрани поблизу овального вікна. Для частоти 1600 Гц максимум коливань лежить біля середини равлика. Сприйняття звукових частот визначається локалізацією максимальних коливань базилярной мембрани. Цікаво відзначити, що видалення мембрани Рейснера і кортиева органа не позначається на параметрах коливань базилярной мембрани. Звідси був зроблений висновок, що вібрації овального вікна викликають механічні коливання базилярной мембрани, а інші структури, що прилягають до неї, важливі для перетворення її механічних коливань у відповідні нервові сигнали.
Нервові імпульси в слуховом нерві виникають у такий спосіб. Руху базилярной мембрани викликають деформацію волоскових кліток, що розташовуються в кортиевом органі усередині равликового каналу. Деформація волоскових кліток приводить до часткової деполяризації їх цитоплазматических мембран. Ці зміни потенціалу діють на немиелинизированние дендрити афферентних нейронів, що знаходяться в контакті з бічною поверхнею і підставою волоскових кліток. У результаті збуджуються слуховие нерви. Звук визначеної частоти надає руху і викликає нервові потенціали від визначеної, але досить протяжної частини базилярной мембрани. Очевидно, у центральній нервовій системі мається механізм, що загострює відчуття звукової частоти так, що в діапазоні 60–1000 Гц людське вухо може розрізняти частоти, що відрізняються на 2–3 Гц.
Слуховий апарат людини винятково чуттєвий. Як відзначалося вище, граничні коливання барабанної перетинки складають 10-11 м, викликувані цим коливання базилярної мембрани ще менше по амплітуді. Не дивно тому, що кортиєв орган не має кровоносних судин, щоб пульсації кров'яного тиску, не надавали руху волоскові кліткам і не викликали слухових відчуттів.
Локалізація джерел звуку заснована на двох механізмах. По-перше, при низьких частотах вухо здатне чуйно уловлювати різницю фаз звукової хвилі в левом і правом юшці. По-друге, при високих частотах вухо головним чином реагує на різницю звуку, що досягло лівого і правого вуха. Навколо голови є звукова «тінь» і якщо різниця інтенсивності досягає 1 дБ, те цього вже досить для зразкової локалізації джерел звуку. При високих частотах через звукову «тіні» навколо голови розходження в інтенсивності звуку може досягати 30 дБ, що дозволяє локалізувати джерело з точністю до ±10о. Цікаво, що частота 3000 Гц, дошкульніше всього сприймана вухом людини, неоптимальна для здійснення обох механізмів (по різниці фаз і по «тіні»), тому при цій частоті важко локалізувати джерело звуку.