Симбиоз (работа 1)
Симбиоз или сожительство двух организмов,— одно из интереснейших и до сих пор еще во многом загадочных явлений в биологии, хотя изучение этого вопроса имеет уже почти столетнюю историю. Явление симбиоза впервые было обнаружено швейцарским ученым Швенденером в 1877 г. при изучении лишайников, которые, как выяснилось, представляют собой комплексные организмы, состоящие из водоросли и гриба. Термин “симбиоз” появился в научной литературе позднее. Он был предложен в 1879 г. Д е Пари.
В ряду симбиозов не последнее место занимают симбиозы с участием водорослей. Водоросли способны вступать в симбиотические отношения не только друг с другом, но и с представителями различных систематических групп организмов, как животного, так и растительного царства (бактериями, одноклеточными и многоклеточными животными, грибами, мхами, папоротниками, голосеменными и покрытосеменными растениями). Однако список таких водорослей весьма ограничен. Из обширнейшей группы сине-зеленых водорослей симбиоз с грибами (лишайниковый симбиоз) способны устанавливать представители не более 5—7 родов, из которых чаще других встречаются носток (Nostoc), глеокапса (Gloeocapsa), сцитонема (Scytonema) и стигонема (Stigonema).
Анализ различных симбиозов вскрыл чрезвычайно многообразный характер взаимоотношений между партнерами, разную степень их влияния друг на друга. Одним из простейших случаев является поселение одних организмов на поверхности других.
К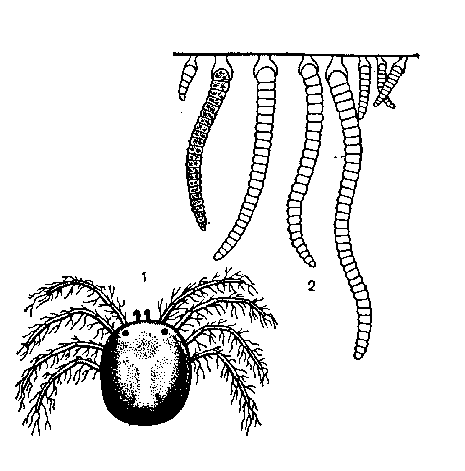
ак
известно, растения, обитающие на других
организмах, но питающиеся самостоятельно,
называют э п и ф и т а м и. К эпифитам
относится и большая группа водорослей.
Особенно часто водоросли эпифитируют
на подводных растениях и водоплавающих
животных, иногда покрывая их плотным
налетом (рис. 46). При
эпифитировании между участниками
устанавливаются очень непрочные и
кратковременные взаимосвязи, которые,
однако, уже можно рассматривать как
симбиотические. Поскольку эпифитирующая
водоросль и хозяин оказывают друг на
друга довольно слабое влияние, эпифитизм
у водорослей принято считать наиболее
примитивной формой симбиоза. Его относят
даже к разряду “безразличных”. С
подобным утверждением полностью
согласиться трудно. Эпифиты действительно
не причиняют прямого вреда организму,
к которому прикрепляются, но косвенный
ущерб при этом все же наносится. Хорошо
известно, например, что обрастающие
водорослями ножки водоплавающих
клещиков, паучков и жучков становятся
менее подвижными, а растения сильно
затеняются расселившимися на них
эпифитами и попадают в условия,
неблагоприятные для фотосинтеза. С
явлением обрастания нередко приходится
сталкиваться при разведении аквариумных
растений, которые могут сильно
угнетаться обитающими на них
водорослями.
К сожалению, явление эпифитизма с биологической точки зрения изучено крайне слабо. Не исключено, что между эпифитом и его хозяином устанавливаются взаимоотношения гораздо более сложные, чем
мы обычно себе представляем.
Помимо поверхностного прикрепления, водоросли могут жить в тканях других организмов—как внеклеточно (в слизи, межклетниках, редко в оболочках мертвых клеток, так и внутриклеточно (в содержимом живых неповрежденных клеток. Такие водоросли по способу обитания относят к группе растений эндофитов.
Внеклеточные и особенно внутриклеточные эндофиты из числа водорослей по сравнению с эпифитами образуют более сложные симбиозы—эндосимбиозы. Для них характерно наличие более или менее тесных, постоянных и прочных связей между партнерами. Эндосимбиозы можно выявить только с помощью специальных цитологических исследований.
Наиболее многочисленную группу составляют эндосимбиозы одноклеточных зеленых и желто-зеленых водорослей с одноклеточными животными. Эти водоросли носят названия соответственно зоохлорелл и зооксантелл. Из многоклеточных животных зеленые и желто-зеленые водоросли образуют эндосимбиозы с пресноводными губками, гидрами и др. . Сине-зеленые водоросли образуют с протозоа и некоторыми другими организмами своеобразную группу эндосимбиозов, получивших название синцианозов; возникающий при этом морфологический комплекс из двух организмов называют цианомом, а сине-зеленые водоросли в нем —цианеллами .
Сопоставление между собой различных эндосимбиозов позволяет наметить последовательные ступени усложнения морфологического и функционального соподчинения партнеров. Так, некоторые эндосимбиозы существуют очень непро
Эпифитизм сине-зеленой водоросли Sokolovia neumaniae на ножках водного клещика Neumania triangulares:
должительное время, а затем распадаются, что является свидетельством их примитивности. Примером этого может служить слизистая колониальная сине-зеленая водоросль воронихиния (Woronichinia naegeliana). Почти в 50% случаев в слизи, окружающей шаровидные колонии этой водоросли, живут другие сине-зеленые водоросли (Lyngbya endophytica и Synechocystis endobiotica. Они интенсивно размножаются там, хотя имеют чрезвычайно бледную, едва заметную окраску. Это, вероятно, обусловлено появлением у них способности утилизировать уже готовые органические соединения, которые в изобилии образуются при распаде слизи.
С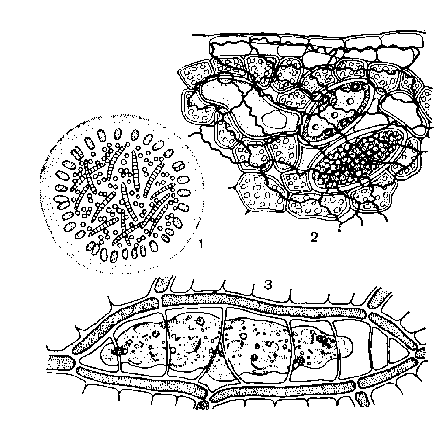
о
временем интенсивное разрастание
водорослей в слизи воронихинии
приводит сначала к подавлению клеток,
а затем к дезорганизации и гибели
всей колонии, а, следовательно, и симбиоза
в целом.
В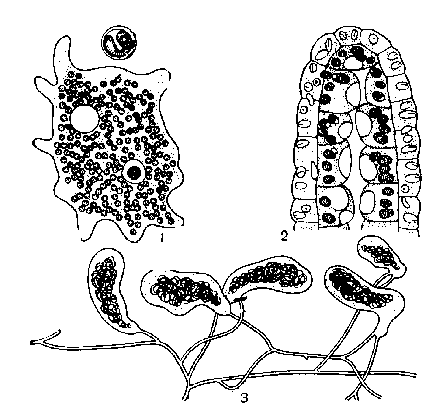
озникает
вопрос: как проникают водоросли в ткани
и клетки других организмов? У некоторых
организмов имеются для этого специальные
приспособления. Так, у мелкого, плавающего
в воде папоротника азоллы
(Azolla) на нижней стороне листьев
располагаются особые полости с узкими
выводными отверстиями, через которые
выделяется наружу слизь. В этих полостях,
независимо от того, в какой географической
точке земного шара растет азолла (в
Америке, Азии, Африке или Австралии),
поселяются колонии строго определенного
вида сине-зеленой водоросли —
анабены (Anahaena azollae). Со
временем полости закрываются и наступает
полная изоляция попавших туда водорослей.
Попытки заражения азоллы представителями
других родов и даже видов сине-зеленых
водорослей успеха не имели. Это
свидетельствует о том, что в процессе
возникновения данного симбиоза между
участниками устанавливается довольно
специфическая физиологическая
взаимозависимость. Этот вывод
подтверждается еще тем,
что вырабатываемые азоллой азотистые
соединения полностью усваиваются
эндосимбиотнрующими здесь экземплярами
анабены, вследствие чего у них отпадает
свойственная свободноживущим
представителям этой сине-зеленой
водоросли функция фиксации атмосферного
азота. В свою очередь, анабена дополнительно
снабжает ткани хозяина кислородом
и другими продуктами своей жизнедеятельности.
Несмотря на существующую у этих симбионтов специализацию физиологических процессов ни один из них не претерпевает сколько-нибудь существенных изменений в своей организации.
Однако так обстоит дело далеко не у всех эндосимбиозов подобного типа. Эндосимбиотический образ жизни водорослей чаще всего приводит к частичной или полной редукции их клеточных оболочек. Например, у живущих в тканях морской губки аплизиллы (Aplysilla) особей сине-зеленой водоросли из рода афанокапса (Aphanocapsa) редукция клеточной оболочки выражается в уменьшении ее толщины. За счет этого снижаются защитные свойства оболочки, но повышается ее проницаемость. Последнее качество, несомненно, улучшает условия транспорта
Внеклеточный симбиоз
Поперечный разрез колонии сине-зеленой водоросли воронихии (крупные клетки по периметру), в слизи которой поселяются другие сине-зеленые водоросли синехоцистис(мелкие клетки и лингбия (удлиненные клетки)
Ткань ряски в межклетниках которой поселяется зеленая водоросль хлорохитрум.
Плазмодий желто-зеленой водоросли миксохлорис в мертвой водоносной клетке сфагнума.
Внутриклеточный симбиоз.
1. Амёба с клеточками зеленой водоросли зоохлореллы внутри, вверху отдельная клетка зоохлореллы при большом увеличении.
2 Продольный разрез через конец шупальца пресноводной зеленой гидры (Hydra viridis) С клетками зоохлореллы в клетках внутреннего слоя гидры.
3. Часть таллома зеленой водоросли геосифон (Geosiphon) разветвленные нити которой оканчиваются крупными пузырями в протоплазме которых живет сине-зеленая водоросль носток.
веществ между клетками губки и эндосимбиотирующей там водоросли.
Эндосимбиозы, относящиеся к разряду внеклеточных, образуют уже довольно устойчивые функциональные и морфологические комплексы. Эта тенденция еще более усиливается у внутриклеточных эндосимбиозов. Механизм проникновения водорослей внутрь клеток других организмов без их повреждения и нарушения нормальной жизнедеятельности остается пока нераскрытым. Отчасти предпосылки для возникновения внутриклеточных эндосимбиозов могут быть заложены в сохранении у клеток некоторых организмов голозойного типа питания. Из всех известных типов питания голозойный тип считается одним из наиболее древних.
У организмов с голозойным типом питания захватываемая добыча, в числе которой оказываются и водоросли, поступает непосредственно внутрь клетки и там переваривается. Однако отдельным захваченным особям, вероятно, в силу стечения благоприятных обстоятельств иногда удается не только сохраниться внутри клеток хозяина в неповрежденном виде, но и выработать приспособления к новым, необычным условиям жизни и начать там размножаться. В результате между организмами устанавливаются отношения нового типа — симбиотические. Вероятно, именно так проникают экземпляры подвижной одноклеточной водоросли эвглены (Euglena gracilis) в эпителиальные клетки задней кишки личинок некоторых видов стрекоз. Клетки эвглены остаются там зелеными на протяжении всего периода совместной жизни. Они, правда, теряют подвижность, но при этом никогда не инцистируются. Очевидно, таким же способом особи одноклеточной зеленой водоросли картерии (Carteria) поселяются в эпидермальных клетках ресничного червя конволюта (Convoluta roscoffensis). Как выяснилось, клетки картерии под влиянием симбиотического образа жизни хотя и претерпевают весьма существенные изменения (полностью редуцируется оболочка, и клетки оказываются окруженными только тонкой плазматической мембраной — плазмалеммой, исчезает стигма, упрощается внутренняя организация жгутиков), но не прекращают фотосинтезировать. В свою очередь, червь приобретает способность питаться за счет продуктов жизнедеятельности водоросли, которые вырабатываются в процессе фотосинтеза. В частности, он может жить в течение 4—5 недель, не получая никакой пищи извне. Однако, когда процесс фотосинтеза прекращается (например, если опыт проводить в темноте), гибнут и водоросль, и червь. Более того, личинки червя, лишенные клеток водоросли, не в состоянии вести самостоятельное существование. Искусственное их заражение водорослями не удается.
Внутриклеточные эндосимбиозы, несомненно, легче устанавливаются с теми организмами, клетки которых не имеют жесткой оболочки на протяжении всего жизненного цикла или по крайней мере на одной из его стадий. Проникновение симбионта внутрь клеток с жесткими оболочками возможно только при условии их частичного или полного разрушения. Последнее может наступить под действием специфических энзимов, вырабатываемых организмом, вступающим в симбиотические отношения. Наблюдаемая в ряде случаев строгая специализация вступающих в симбиоз организмов, вероятно. объясняется именно этим обстоятельством. К сожалению, все попытки обнаружить хотя бы следы подобного рода энзимов пока успехом не увенчались.
Одни внутриклеточные эндосимбиозы. как это происходит у личинок стрекоз, периодически распадаются и вновь возобновляются: другие — непрерывно поддерживаются из поколения в поколение, так как в этих случаях между участниками устанавливаются прочные и продолжительные связи. Последняя группа эндосимбиозов. очевидно, могла возникнуть вследствие утраты той фазы в жизненном цикле организма-хозяина, которая была благоприятна для проникновения симбионта в его клетки. С этого момента, по-видимому, и начинается тесная совместная жизнь двух организмов. В таких случаях переход к симбиотическому способу существования неизбежно сопровождается рядом адаптационных изменений у обоих организмов. Иногда эти изменения морфологически незначительны и симбионт можно узнать (например, носток у геосифона, рис. 48,3), а иногда они настолько существенны, что симбиотирующие водоросли невозможно идентифицировать ни с одной из свободноживущих водорослей.
Так, в вакуолях одного из видов ресничной инфузории парамеции (Paramecium bursaria) неизменно присутствует зеленая одноклеточная водоросль. По морфологии и особенностям поведения ее можно лишь условно отнести к протококковой водоросли из рода хлорелла (Chlorella). Установлено, что клетки водоросли делятся независимо от деления парамеции. Каждая из вновь образующихся дочерних клеток (автоспор) водоросли немедленно заключается в особую вакуолю и в таком виде в дальнейшем распределяется между дочерними особями инфузории.
В ряде случаев между симбионтами складываются настолько тесные взаимозависимые отношения, что вне симбиоза они жить уже не могут. Очевидно, они необратимо утрачивают способность самостоятельно вырабатывать целый ряд веществ, которые в готовом виде поступают от симбиотирующих с ними водорослей. Реальность подобного предположения полностью подтвердилась в опытах с гидрой, которая, оказывается, в нужном количестве получает мальтозу из клетки симбиотирующей там зеленой водоросли, систематическую принадлежность которой точно установить так и не удалось.
Иногда нераспадающиеся эндосимбиозы приводят к образованию такого комплекса, симбио-тическая природа которого выявляется с большим трудом. Так случилось с двумя водорослями — цианофорой и глаукоцистисом.
В 1924 г. была описана новая для науки водоросль, названная цианофорой парадоксальной (Cyanophora paradoxa, табл. 5, 7). Позднее детальное изучение этого организма показало, что цианофора представляет собой симбиоз бесцветной одноклеточной водоросли криптомонады (отдел Pyrrophyta) и поселяющейся в ней внутриклеточно сине-зеленой водоросли (цпанеллы) из рода хроококкус (Chroococcus, отдел Cyanophyta). Клетки последней под влиянием симбиотического образа жизни настолько сильно видоизменяются, что теряют свой типичный облик. Это выражается главным образом в сильной редукции клеточной оболочки.
Она уменьшается не только по толщине, но и по числу входящих в ее состав слоев: вместо четырехслойной, обычно характерной для свободноживущих сине-зеленых водорослей, она становится двухслойной.
Еще большим преобразованиям подвергаются цианеллы, входящие в состав глаукоцистиса (Glaucocystis nostochinearum) — очень своеобразной одноклеточной водоросли, описанной в конце прошлого века. Ее систематическое положение долгое время не удавалось точно определить. На основании сине-зеленой окраски ее сначала отнесли к отделу Cyanophyta. В дальнейшем выявление целого ряда признаков, абсолютно несвойственных сине-зеленым водорослям (наличие морфологически оформленного ядра, окрашенных телец, размножение посредством автоспор), позволили отнести этот организм к зеленым водорослям (отдел Chlorophyta). Только в 30-е годы текущего столетия было наконец установлено, что глаукоцистис представляет собой крайне своеобразную форму эндосимбиоза обесцветившейся одноклеточной водоросли, близкой к роду ооцистис (Oocystis), и палочковидной сине-зеленой водоросли, которая претерпела здесь такие сильные преобразования, что установить точно ее систематическую принадлежность не представляется возможным. В равной степени это может быть любой модифицированный представитель из ряда родов одноклеточных палочковидных сине-зеленых водорослей. В симбиозах подобного рода глаукоцистис является пока единственным примером установления столь тесных взаимоотношений между партнерами. Сине-зеленые водоросли (цианеллы) располагаются в клетках глаукоцистиса либо упорядоченно в виде двух групп, либо беспорядочно, случайно.
Цианеллы и свободноживущие сине-зеленые водоросли по своей тонкой организации ничем не отличаются друг от друга. Примечательно, что в цианеллах отсутствуют включения запасных питательных веществ, представленных различными метаболическими гранулами. По всей видимости, надобность в этом отпадает, поскольку цианеллы получают необходимые им вещества прямо из клетки хозяина. В то же время цианеллы поставляют в клетки хозяина некоторые продукты, которые вырабатываются ими в процессе фотосинтеза. Об этом свидетельствует присутствие в цитоплазме бесцветных клеток организма-хозяина крахмальных зерен. Явление это весьма необычное, поскольку у всех хлорофиллоносных зеленых растений единственным местом локализации крахмальных зерен является пластида (хлоропласт). В условиях симбиоза его участники достигают, вероятно, максимальной специализации, в силу которой симбиотирующие сине-зеленые водоросли принимают на себя функции хлоропластов, но ими не становятся. В пользу последнего свидетельствует существенная разница в организации цианелл и пластид. У клеток бесцветного симбионта глаукоцистиса утрачивается способность к самостоятельному образованию крахмала, который образуется там, очевидно, при непосредственном участии цианелл.
Изучение с помощью электронного микроскопа цианелл, входящих в состав глаукоцистиса, выявило у них сильную степень редукции клеточной оболочки. Она сохраняется здесь в виде едва заметного контура, который можно обнаружить лишь при условии высокого качества фиксации и обработки материала. Более тщательное исследование цианелл показало, что их окружает лишь тонкая (100 ^ 10.4) мембрана, называемая плазмалеммой. Такая степень редукции клеточного покрова — уникальное явление среди вступающих в симбиоз сине-зеленых водорослей.
Из приведенной характеристики цианелл видно, что они представляют собой не что иное, как клетки сине-зеленых водорослей, лишенных запасных веществ и клеточных оболочек.
Деление цианелл, как и клеток свободноживущих сине-зеленых водорослей, осуществляется путем перетяжки пополам. Оно автономно и не приурочено к периоду размножения клетки-хозяина. В каждую его дочернюю клетку обычно попадает по несколько цианелл. Таким образом обеспечивается непрерывность симбиоза. В отличие от органелл распределение цнанелл между дочерними клетками хозяина носит случайный характер, поэтому их число там сильно варьирует. Не вызывает никакого сомнения, что само деление и характер расхождения цианелл по дочерним клеткам регулируется не хозяином, что было бы вполне естественно, если бы они превратились в органеллы, а самими цианеллами, сохранившими все свойства клеток. Однако даже в условиях такого высокоразвитого симбиоза, примером которого является глаукоцистис, оба партнера все же сохраняют своп индивидуальные черты и автономность. Об этом свидетельствует их способность к раздельному существованию вне клеток хозяина. В специально подобранной питательной среде изолированные симбионты ведут себя как самостоятельные организмы. Они там не только успешно растут и развиваются, но II размножаются.
Среди симбиозов, образованных с участием водорослей, наибольший интерес представляет симбиоз водорослей с грибами, известный под названием лишайникового симбиоза.
ВЗАИМООТНОШЕНИЯ ГРИБА И ВОДОРОСЛИ В ТЕЛЕ ЛИШАЙНИКА
Вопрос взаимоотношения гриба и водоросли в слоевище лишайника занимал умы ученых еще в конце прошлого столетия, да и в паше время продолжает волновать лихенологов. Со дня открытия С. Швендснера прошло более 100 лет. За этот период появилось не менее десятка теорий, пытающихся объяснить отношения между грибом и водорослью, однако среди них нет ни одной общепризнанной и окончательно доказанной. С. Ш в е н д е не р, обнаружив, что лишайник состоит из гриба и водоросли, предположил, что гриб в слоевище паразитирует на водоросли. Однако он ошибочно отвел грибу роль хозяина, а водоросли — раба.
Но уже в те времена некоторые ученые выдвинули мысль о двустороннем паразитизме компонентов лишайника — гриба на водоросли и водоросли на грибе. При этом было высказано предположение, что гриб и водоросль в слоевище лишайника находятся в полном морфофизиологическом единстве и связаны между собой так же, как корни и листья цветковых растений. Такое сравнение, безусловно, было совсем необоснованным.
Наибольшее распространение среди ученых того времени получила теория мутуалистического симбиоза. Сторонники этой теории считали, что в слоевище лишайника гриб и водоросль находятся во взаимовыгодном симбиозе: водоросль “снабжает” гриб органическими веществами, а гриб “защищает” водоросль от чрезмерного нагревания и освещения и “обеспечивает” ее водой и неорганическими солями. Однако в 1873 г. этой идеалистической теории был нанесен удар. Известный французский исследователь Е.Борн е, изучая анатомическое строение слоевища лишайников, обнаружил внутри водорослевых клеток грибные отростки — гаустории, всасывающие органы гриба. Это позволяло думать, что гриб использует содержимое клеток водорослей, т. е. ведет себя как паразит.
За прошедшие со времен Борне 100 лет в слоевище лишайников было открыто и описано много различных форм абсорбционных, или всасывающих, гиф гриба. Эти гифы плотно прижимаются к клетке водоросли или проникают в нее и служат, как предполагают, для передачи веществ, которые образуют водоросли в результате своей жизнедеятельности, грибному компоненту.
О том, что в слоевище лишайника происходит обмен веществами между грибом и водорослью, ученые стали говорить сразу после открытия двойственной природы лишайников. Однако некоторые экспериментальные подтверждения этим предположениям были получены лишь за последние три десятилетия. Применение новейших методов физиологических исследований с использованием меченых атомов углерода и азота, особых красящих веществ и некоторых других позволило установить, что гриб получает вещества, ассимилируемые водорослью, и ведет себя в слоевище лишайника как паразитический организм. Однако для существования как самого гриба, так и лишайника в целом необходимо, чтобы водоросль, окруженная со всех сторон грибными гифами, все-таки могла жить и более или менее нормально развиваться. Если гриб начнет проявлять себя слишком активно, поражать все без исключения водоросли и, использовав их содержимое, уничтожать их, это в конце концов может привести к гибели всех водорослей слоевища. Но тогда, уничтожив весь свой запас питания, по-
гибнет и сам гриб, а значит, перестанет существовать и лишайник.
Гриб должен использовать лишь часть водорослей, оставляя резерв — здоровые и нормальные водоросли, содержимым которых он мог бы питаться.
Учеными были замечены любопытные защитные реакции со стороны лишайниковых водорослей. Например, одновременно с проникновением гаустория в клетку водоросли эта клетка делилась. При этом плоскость деления, как правило, проходила как раз через участок, занятый гаусторием, а образовавшиеся в результате этого процесса дочерние клетки были свободны от гаусториев. Было замечено также, что обычно гриб поражает водоросли, уже достигшие определенной стадии зрелости. В молодых растущих водорослях происходит энергичное отложение веществ в оболочке клетки и быстрое ее утолщение. Эта толстая оболочка клетки фикобионта препятствует проникновению абсорбционных органов гриба. Однако большей частью защитная реакция водорослей против активности грибного компонента очень слаба.
Способность водорослей нормально развиваться и даже размножаться в слоевище лишайника сохраняется скорее благодаря умеренности паразитизма самого гриба.
Ученые отмечают, что степень паразитизма гриба на водоросли различна не только у разных видов лишайников, но даже в одном и том же слоевище. Резкий паразитизм обнаружен лишь у примитивных лишайников. Гаустории, проникающие глубоко внутрь протопласта водоросли, пока что были найдены лишь у наиболее просто организованных форм, в слоевище которых еще нельзя различить оформленных дифференцированных слоев. В слоевищах более высокоорганизованных лишайников часть клеток водорослей поражена грибными гифами, а остальные продолжают нормально жить и развиваться. Обычно у высокоорганизованных форм лишайников паразитизм гриба на водоросли носит весьма умеренный характер: прежде чем гриб убьет пораженные им клетки, успевает вырасти одно или несколько поколений водорослей.
Отношения между мико- и фикобионтом в слоевище лишайника не сводятся только к паразитизму гриба на водоросли. Ученые предполагают, что эти отношения гораздо сложнее. Еще в начале нашего века крупнейший русский лихенолог А. А. Е ленки н, изучая анатомическое строение лишайников, обнаружил в их слоевище некральные зоны водорослей — скопления отмерших, потерявших зеленую окраску клеток, расположенные несколько ниже зоны живых водорослей. К этим бесцветным мертвымклеткам водорослей тоже тянулись грибные гифы. Это привело А. А. Еленкина к мысли, что гриб в слоевище лишайника вначале проявляет себя как паразитический организм, поражая живые клетки водоросли и используя их содержимое. Затем, убив водоросль, гриб переходит к сапрофитному способу питания, поглощая и ее мертвые остатки. Таким образом, гриб в слоевище лишайника ведет себя и как паразит, и как сапрофит. И отношения между грибом и водорослью в слоевище лишайника А. А. Еленкин назвал эндопаразитосапрофитизмом.
Интересную мысль о взаимоотношении компонентов в слоевище лишайника высказал в 60-х годах нашего столетия крупнейший советский лихенолог А. Н. О к сне р. Он пришел к выводу, что водоросль в слоевище лишайника, полностью изолированная от внешней среды грибной тканью, обязательно должна забирать у грибного компонента все необходимые для своего существования вещества, за исключением тех органических соединений, которые она сама вырабатывает на свету в процессе ассимиляции углекислоты. К этим жизненно необходимым для водоросли веществам относится прежде всего вода, а также минеральные соли, азотистые и некоторые другие неорганические соединения. Следовательно, и водоросль в слоевище лишайника проявляет себя как паразит. Причем это вовсе не противоречит общему характеру ее питания. Как показало изучение лишайниковых водорослей в чистых культурах, многие из них, будучи большей частью автотрофными организмами, способны и к миксотрофному питанию.
Таким образом, ученые считают, что водорослевый и грибной компоненты лишайника находятся в очень сложных взаимоотношениях. Микобионт ведет себя как паразит и сапрофит на теле водоросли, а фикобионт, в свою очередь, паразитирует на лишайниковом грибе. При этом паразитизм фикобионта всегда носит более умеренный характер, чем паразитизм гриба.
О
днако
все высказанные по этому поводу точки
зрения до сих пор 'остаются лишь догадками
и большей частью не подтверждены
экспериментально: лишайники оказались
очень трудным объектом для
физиологических исследований. Ученые
пока не научились выращивать и поддерживать
в живом состоянии слоевище лишайников
в искусственных условиях. Тот контакт
между грибом и водорослью, который с
такой легкостью достигается в природе
(достаточно вспомнить многообразие
лишайников!), никак не удается воспроизвести
в лабораторных условиях. Наоборот,
при переносе лишайников в лабораторию
этот контакт легко нарушается и растение
просто погибает. Время от времени
появляются сообщения об удачных опытах
выращивания лишайника в условиях
лаборатории, но пока эти сообщения
единичны и не всегда достоверны.
Одной из причин неудач подобных попыток можно считать чрезвычайно медленный рост лишайников. Лишайники — многолетние растения. Обычно возраст взрослых слоевищ, которые можно увидеть где-нибудь в лесу на стволе деревьев или на почве, составляет не менее 20—50 лет. В северных тундрах возраст некоторых кустистых лишайников рода кладония достигает 300 лет. Слоевище лишайников, имеющих вид корочки, в год дает прирост всего 0,2—0,3 мм.
Кустистые и листоватые лишайники растут несколько быстрее — в год их слоевище увеличивается на 2—3 мм. Поэтому, чтобы вырастить взрослый лишайник в лаборатории, требуется не менее 20 лет, а может быть, и вся жизнь исследователя. Трудно проводить столь долговременный эксперимент!
Вот почему физиологические особенности лишайников, в том числе взаимоотношения компонентов, как правило, изучают на культурах изолированных мико- и фикобионтов. Этот метод очень перспективен, так как позволяет ставить длительные и воспроизводимые опыты. Но, к сожалению, данные, полученные этим методом, не могут полностью отразить те процессы, которые происходят в целом слоевище лишайника.
И тем более мы не вправе считать, что в природе, в естественных условиях, в слоевищах лишайника эти процессы протекают точно так же, как в культурах изолированных симбионтов. Вот почему все теории, пытающиеся объяснить взаимоотношения компонентов лишайников, остаются пока лишь догадками.
Более успешным оказалось изучение форм контакта между гифами гриба и клетками водорослей в слоевищах лишайников. Как показали исследования с применением электронной микроскопии, в слоевище лишайников можно встретить по крайней мере пять типов контакта между грибными гифами и водорослевыми клетками (рис. 289).
Чаще всего отдельная клетка водоросли и клетка грибной гифы находятся в непосредственном контакте друг с другом. В таком случае гриб образует специальные абсорбционные, всасывающие органы, которые проникают внутрь водорослевой клетки или плотно прижимаются к ее оболочке.
В настоящее время среди абсорбционных органов гриба в слоевище лишайников различают несколько типов: гаустории, импрессории и аппрессории.
Формы контакта между гифами гриба и клетками водорослей в слоевище лищайников.
Гаустории— это боковые выросты гиф гриба, которые прорывают оболочку клетки водоросли и проникают в ее протопласт (рис. 289, 2). Обычно в клетке водоросли развивается одни гаусторий, но иногда их может быть и два. В слоевище лишайника гаустории встречаются в большом количестве и существуют продолжительное время. Было замечено, что в оболочках молодых гаусториев нет отложений целлюлозы, которая могла бы затруднять обмен между клеткой водоросли и гифой гриба. Старые гаустории почти всегда одеты довольно толстым слоем целлюлозы. Различают интрацеллюлярные (внутриклеточные) и интрамембранные (внутриоболочковые) гаустории.
Интрацеллюлярные гаустории полностью прорывают оболочку клетки водоросли и проникают глубоко внутрь ее протопласта (рис.289, 3). Интрацеллюлярные гаустории образуются в случае резкого паразитизма гриба на водоросли. Это особенно характерно для лишайников с примитивным строением слоевища.
У более высокоорганизованных лишайников образуются только интрамембранные гаустории. Они прорывают оболочку клетки водоросли и достигают ее протопласта, но не углубляются в него, а остаются в оболочке водорослевой клетки (рис. 289, 5). Наибольшее количество интрамембранных гаусториев образуется в слоевище лишайников весной, в начале вегетационного периода. С наступлением осени они далеко отступают от протопласта водоросли.
Другой тип всасывающих органов гриба — импрессории— тоже боковые выросты грибных гиф, но, в отличие от гаусториев, они не прорывают оболочку клетки водоросли, а вдавливают ее внутрь (рис. 289, 6, 7). Импресcopии отмечены у очень многих лишайников, например у широко распространенной пельтигеры (Peltigera).
Интересно, что в слоевищах, произрастающих во влажных местообитаниях, импрессории почти не развиваются, у тех же видов в сухих местообитаниях они образуются в большом количестве. При длительной засухе число импрессориев также увеличивается. Предполагают, что в засушливые периоды и в сухих местообитаниях гриб, чтобы удовлетворить потребности в питании, увеличивает свою всасывающую поверхность за счет увеличения количества и размеров импрессориев.
В отличие от гаусториев и импрессориев, образованных боковыми отростками гифы, аппрессории образуются вершиной грибной гифы. Такая вершина гифы плотно прижимается снаружи к оболочке клетки водоросли, никогда не проникая ни в ее протопласт, ни в ее внутренний слой (рис. 289, 8).
Наличие в слоевищах многих лишайников абсорбционных органов гриба хорошо доказывает паразитическую сущность отношений микобионта к фикобионту. Но во многих случаях у лишайникового гриба все же не удается обнаружить особых абсорбционных органов, чаще всего у лишайников, фикобионт которых имеет тонкие оболочки клеток. В таких случаях уже внешний контакт гифы гриба и клетки водоросли может обеспечить обмен веществами между ними. Так, например, обстоит дело у многих видов рода кладония. Фикобионтом кладонии является одноклеточная зеленая водоросль требуксия. У этих лишайников отдельные клетки водорослей окружены со всех сторон тонкими тонкостенными гифами, иногда поделенными на мелкие клеточки. Эти гифы, которые носят название обволакивающих или контактных, не проникают в протопласт клеток водоросли и не внедряются в их оболочку, а просто окружают клетки со всех сторон, так что каждая из них становится похожей на маленький шар, охваченный пальцами рук (рис. 289,7). Иногда гифы полностью оплетают водоросли в виде сплошного покрова и при этом, сливаясь своими стенками, даже образуют клеточную псевдопаренхимную ткань. На первый взгляд кажется, что водоросли не особенно страдают от плотного окружения гифами гриба: они долго сохраняют свою зеленую окраску и продолжают интенсивно делиться.
Но в более старых участках слоевища можно найти немало отмерших обесцвеченных клеток — гриб рано или поздно все-таки убивает водоросли. Такой же тип контакта между гифами гриба и клетками водорослей был найден у некоторых слизистых и базидиальных лишайников.
У ряда лишайников, в слоевище которых встречаются нитчатые улотриксовые водоросли, можно наблюдать еще один тип контакта. Как правило, в таком случае нити водорослей бывают целиком покрыты грибными гифами. Причем лишь иногда гифы образуют на поверхности водорослевой нити рыхлую сетку. Чаще же они располагаются очень густо и, срастаясь своими стенками, образуют сплошной чехол. Отдельная лопасть такого лишайника имеет вид тончайшего волоса. Под микроскопом она напоминает полую трубку, стенки которой образованы сросшимися грибными гифами; внутри трубки тянется нить водоросли.
У слизистых лишайников семейства коллемовых (Collemataceae) обычно не наблюдается никакого контакта между грибными гифами и клетками водорослей. Слоевище коллемовых не имеет дифференцированной структуры: нити водоросли носток разбросаны в беспорядке среди грибных гиф по всей толще слоевища (рис. 297, 2). Никаких абсорбционных отростков в клетках водорослей обычно не заметно; гифы гриба и нити сине-зеленой водоросли расположены друг около друга, не вступая в видимый контакт. Предполагают, что в данном случае гриб поглощает органические вещества, ассимилируемые водорослями, прямо из слизи, которая обычно окружает нити ностока. Однако более тщательное изучение этих лишайников показало, что у многих видов коллемы (Collema) в слоевище время от времени образуются специальные абсорбционные гифы, которые тесно прижимаются к одной из клеток водорослевой нити, а через некоторое время можно наблюдать отмирание этой клетки.
Описанные выше формы контакта между гифами микобионта и клетками водорослей, по всей видимости, не исчерпывают всего многообразия способов, с помощью которых гриб и водоросль в слоевище лишайников устанавливают между собой тесную связь. Исследования в этом направлении только начинаются. Можно думать, что дальнейшее изучение тончайших структур лишайникового слоевища с помощью электронного микроскопа не только даст много нового в описании физических контактов между грибным и водорослевым компонентами лишайников, но и откроет новые горизонты в понимании их взаимоотношений
_________________________________________
Пока лишайники являют собой единственный строго доказанный случай возникновения одного совершенно нового организма из двух. Этот факт послужил толчком для поиска синтетических форм в самых различных систематических группах растений и животных. Однако все усилия, приложенные в данном направлении, оказались пока безрезультатными. Тем не менее, предположение о возможности существования синтетических форм организмов оказалось настолько заманчивым, что в биологии появляется новое течение. В отличие от обычных, прочно утвердившихся взглядов биологов на происхождение организмов как на переход от простого к сложному путем дифференциации, зарождается новое представление о возникновении сложного организма из более простых путем синтеза. Некоторые биологи стали рассматривать растительную клетку не как продукт постепенной дифференциации протопласта, а как симбиотический комплекс, синтезированный из нескольких простых организмов. Эти идеи зародились и получили наиболее полное развитие в трудах наших отечественных ученых.
Впервые предположение о важной роли формативного симбиоза (т.е. симбиоза, приводящего к образованию новых форм) в эволюции организмов было высказано академиком А. С. Фаминциным в 1907 г. Развивая дальше эти мысли, К. С. Мережковский в 1909 г. сформулировал гипотезу симбиогенного происхождения организмов и назвал ее “теорией симбиогенезиса”. В дальнейшем она получила широкую известность среди биологов. В 20-е годы ее поддержал и развил дальше известный советский ботаник Б. М. Коз о-П о ля нс кий. В наши дни эти идеи, уже на новом уровне развития биологии, были возрождены американской исследовательницей Сага н-М а р гу л и с в ее гипотезе происхождения эукариотических клеток. В соответствии с этой гипотезой такие клеточные органеллы, как митохондрии, базальные тела жгутиков и пластиды эукариотических клеток, возникли из симбиотирующих в них прокариотических клеток сине-зеленых водорослей и бактерий. В качестве основного довода приводятся некоторые черты сходства в составе, строении и поведении перечисленных органелл и прокариотов. Бесспорно, эти факты заслуживают самого пристального внимания. Однако они недостаточны для обоснования гипотезы симбиогенеза, поскольку черты сходства, как известно, могут появляться у разных по происхождению структур или организмов II вследствие параллелизма в эволюции. Так, система стигма — жгутик у золотистых, желто-зеленых и бурых водорослей по внешнему виду и функциям очень напоминает палочки сетчатки глаза животных, хотя весь процесс их заложения и ход онтогенетического развития говорят о том, что об общности происхождения этих образований не может быть и речи.
Исследование симбиотических организмов в электронном микроскопе показывает, что даже у такого высокоразвитого симбиоза, как глаукоцистис, партнеры сохраняют свои индивидуальные черты и автономность. Анализ симбиозов водорослей с различными организмами выявляет определенную направленность в развитии отношений между партнерами, главным образом по линии максимальной специализации функций и вызванных этим обстоятельством структурных перестроек при сохранении их как самостоятельных организмов; это идет вразрез с положениями гипотезы симбиогенеза. Все это свидетельствует о том, что в настоящее время гипотеза симбиогенеза находится на той стадии разработки, когда логические построения явно преобладают над фактами.
Безусловно, симбиоз может привести к созданию новых организмов, что подтверждается появлением такой своеобразной растительной группы, как лишайники. Отрицать роль симбиоза в эволюции нельзя. И все же очевидно, что это не единственный и не основной путь образования новых форм жизни. С одной стороны, к такому выводу подводит факт существования именно лишайников, поскольку они образуют крайне специализированную и обособленную группу организмов, представляющих слепую ветвь филогенетического развития. С другой стороны, сейчас накапливается большой фактический материал по тонкой организации клетки. Он дает возможность воссоздать картину вероятного обособления и усложнения организации некоторых клеточных органелл у водорослей. Кстати, именно отсутствие подобного рода фактов в свое время как раз и стимулировало зарождение гипотезы симбиогенеза.
Список использованной литературы: “Жизнь растений” (Том 3; Стр72 “Сожительство водорослей с другими организмами”.-Т. В. Седова; Стр385 “Взаимоотношения гриба и водоросли в теле лишайника” Н. С. Голубкова.)
Под ред. профессора М. М. Голлербаха.
Москва “Просвещение” 1977 год
487 стр.
Лекции Н. К Христофоровой “Биотические отношения” (Симбиоз)