Частная физиология центральной нервной системы
МИНИСТЕРСТВО ЗДРАВООХРАНЕНИЯ УКРАИНЫ
ЛУБЕНСКОЕ МЕДИЦИНСКОЕ УЧИЛИЩЕ
РЕФЕРАТ
С ФИЗИОЛОГИИ
НА ТЕМУ:

 Частная
физиология центральной нервной системы
Частная
физиология центральной нервной системы
Выполнила:
студентка группы Ф-31
Чиженко Яна
Лубны 2009
Особенности строения спинного мозга. Спинной мозг является наиболее древним и примитивным отделом, центральной нервной системы, В центральной части спинного мозга находится серое вещество. Оно состоит преимущественно из нервных клеток и образует выступы — задние, передние и боковые рога. В прилежащих спинальных ганглиях располагаются афферентные нервные клетки. Длинный отросток афферентной клетки находится на периферии и образует воспринимающее окончание (рецептор), а короткий заканчивается у клеток задних рогов. В передних рогах расположены эффекторные клетки (мотонейроны), аксоны которых иннервируют скелетные мышцы; в боковых рогах — нейроны вегетативной нервной системы. В сером веществе находятся многочисленные вставочные нейроны. Среди них обнаружены особые тормозные нейроны — клетки Реншоу, названные так по имени автора, который впервые их описал. Вокруг серого вещества располагается белое веодество спинного мозга. Оно образовано нервными волокнами восходящих и нисходящих путей, соединяющих различные участки спинного мозга друг с другом, а также спинной мозг с головным (рис. 71).
Функции спинномозговых корешков. Связь спинного мозга с периферией осуществляется посредством нервных волокон, проходящих в спинномозговых корешках; по ним к спинному мозгу поступают афферентные импульсы и проходят от него на периферию эфферентные импульсы. По обеим сторонам спинного мозга располагается 31 пара передних и задних корешков.
Функции спинномозговых корешков были выяснены при помощи методов перерезки и раздражения.
Выдающийся шотландский анатом и физиолог Белл и французский исследователь Мажанди установили, что при односторонней перерезке передних корешков спинного мозга отмечается паралич конечностей этой же стороны, чувствительность же сохраняется полностью. Перерезка задних корешков приводит к утрате чувствительности, двигательная функция при этом сохраняется. Таким образом, было показано, что афферентные импульсы поступают в спинной мозг через задние корешки (чувствительные), эфферентные импульсы выходят через передние корешки (двигательные).

Спинной мозг выполняет две функции: рефлекторную и проводниковую.
Рефлекторная функция спинного мозга. В спинной мозг поступают афферентные импульсы от рецепторов кожи, проприорецепторов двигательного аппарата, интерорецепторов кровеносных сосудов, пищеварительного тракта, выделительных и половых органов. Эфферентные импульсы от спинного мозга идут к скелетным мышцам (за исключением мышц лица), в том числе к дыхательным — межреберным и диафрагме. Кроме того, от спинного мозга по вегетативным нервным волокнам импульсы поступают ко всем внутренним органам, кровеносным сосудам, потовым железам и т. д.
Мотонейроиы спинного мозга возбуждаются за счет афферентных импульсов, поступающих к ним от различных рецепторов организма. Однако уровень активности мотонейронов зависит не только от центростремительных импульсов, но и от сложных внутрицентральных взаимоотношений. Большая роль в регуляции деятельности мотонейронов принадлежит нисходящим влияниям головного мозга (коры больших полушарий, ретикулярной формации ствола мозга, мозжечка и др.), а также внутриспинальным воздействиям многочисленных вставочных нейронов. Среди вставочных нейронов особая роль принадлежит клеткам Реншоу. Эти клетки образуют на мотонейронах тормозные синапсы. При возбуждении клеток Реншоу активность мотонейронов притормаживается, что предупреждает перевозбуждение и контролирует их работу. Деятельность мотонейронов спинного мозга контролируется также потоком, импульсов, идущих от пропри орецепторов мышц (обратная афферентация).

Социальные рефлексы, то есть рефлексы, присущие самому спинному мозгу, молено изучить в чистом виде только после отделения спинного мозга от головного (спинальное животное). Первым следствием поперечной перерезки между продолговатым и спинным мозгом является спинальный шок, который длится от нескольких минут до нескольких недель в зависимости от уровня развития центральной нервной системы. Спинальный шок проявляется резким падением возбудимости и угнетением рефлекторных функций всех нервных центров, расположенных ниже места перерезки. В возникновении спинального шока большое значение имеет устранение нервных импульсов, поступающих к спинному мозгу из вышележащих отделов центральной нервной системы, в том числе от нейронов ретикулярной формации ствола мозга.
По прекращении спинального шока постепенно восстанавливаются рефлекторная деятельность, скелетных мышц, величина артериального давления, рефлексы мочеиспускания, дефекации и ряд половых рефлексов. У спинального животного не восстанавливаются произвольные движения, чувствительность и температура тела, а также дыхание. Спинальные животные могут жить только при условии искусственного дыхания. Следовательно, центры, регулирующие эти функции, находятся в вышележащих отделах центральной нервной системы.
Рефлекторные центры спинного мозга. В шейном отделе спинного мозга находится центр диафрагмального нерва, центр сужения зрачка; в шейном и грудном отделах — центры мыши, верхних конечностей, мышц груди, спины и живота; в поясничном отделе — центры мышц нижних конечностей, в крестцовом отделе — центры мочеиспускания, дефекации и половой деятельности, в боковых рогах грудного и поясничного отделов спинного мозга — центры потоотделения и спинальные сосудодвигательные центры.
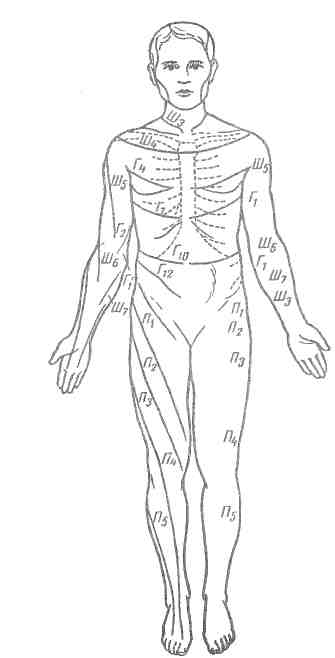
Рефлекторные дуги отдельных рефлексов проходят через определенные сегменты спинного мозга. Возбуждение, возникшее в рецепторе по центростремительному нерву, поступает в соответствующий отдел спинного мозга. Центробежные волокна, выходящие из спинного мозга в составе передних корешков, иннервируют строго определенные участки тела (рис. 72).
Проводниковая функция спинного мозга. Через спинной мозг проходят восходящие и нисходящие пути.
Восходящие нервные пути передают информацию от тактильных, болевых, температурных рецепторов кожи и от проприорецепторов мышц через нейроны спинного мозга и другие отделы центральной нервной системы к мозжечку и коре большого мозга.
Нисходящие нервные пути (пирамидный и экстрапирамидный) связывают кору большого мозга, подкорковые ядра и образования ствола мозга с мотонейронами спинного мозга. Они обеспечивают влияние высших отделов центральной нервной системы на деятельность скелетных мышц.
Непосредственным продолжением спинного мозга у всех позвоночных животных и человека является продолговатый мозг.
Продолговатый мозг и мост мозга (варолиев мост) вместе со средним и промежуточным мозгом образуют ствол мозга. Нисходящих в состав ствола мозга входит большое количество ядер, восходящих и путей. Важное функциональное значение имеет находящаяся в стволе мозга ретикулярная формация (см. рис. 73).
В продолговатом мозге по сравнению со спинным мозгом нет четкого сегментарного распределения серого и белого вещества.
Скопление нервных клеток приводит к образованию ядер, являющихся центрами более или менее сложных рефлексов. Из 12 пар черепных нервов, связывающих головной мозг с периферией организма — его рецепторами и эффекторами, восемь пар (V—XII) берут свое начало в продолговатом мозге.
Продолговатый мозг выполняет две функции — рефлекторную и проводниковую.
Рефлекторная функция продолговатого мозга. В продолговатом мозге находятся центры как относительно простых, так и более сложных рефлексов. За счет деятельности продолговатого мозга осуществляются: 1) защитные рефлексы (мигание, слезоотделение, чиханье, кашлевой и рвотный рефлексы); 2) установочные рефлексы, обеспечивающие тонус мускулатуры, необходимый для поддержания позы и осуществления рабочих актов; 3) лабиринтные рефлексы, способствующие правильному распределению мышечного тонуса между отдельными группами мышц и установке той или иной позы тела; 4) рефлексы, связанные с функциями систем дыхания, кровообращения, пищеварения.
Проводниковая функция продолговатого мозга. Через продолговатый мозг проходят восходящие пути от спинного мозга к головному и нисходящие пути, связывающие кору больших полушарий со спинным мозгом. Продолговатый мозг и мост мозга имеют собственные проводящие пути, соединяющие ядро и оливу вестибулярного нерва с мотонейронами спинного мозга.
Через восходящие пути и черепные нервы продолговатый мозг получает импульсы от рецепторов мышц лица, шеи, конечностей и туловища, от кожи лица, слизистых оболочек глаз, носовой и ротовой полости, от рецепторов слуха, вестибулярного аппарата, рецепторов гортани, трахеи, легких, интерорецепторов пищеварительного аппарата и сердечнососудистой системы.
Функции продолговатого мозга изучаются на бульбарных животных, у которых поперечным разрезом продолговатый мозг отделяется от среднего. Следовательно, жизнь бульбарных животных поддерживается за счет деятельности: спинного и продолговатого мозга. У таких животных отсутствуют произвольные, движения, отмечается потеря всех видов чувствительности, нарушается регуляция температуры тела (теплокровное животное превращается в холоднокровное). У бульбарных животных сохраняются рефлекторные- реакции и осуществляется регуляция функций внутренних органов.
Рефлекторные центры продолговатого мозга, В продолговатом мозге располагается ряд жизненно важных центров: дыхательный, сердечнососудистый и пищевой центры. За счет деятельности этих центров осуществляется регуляция дыхания, кровообращения и пищеварения. Таким образом, основная биологическая роль продолговатого мозга заключается в обеспечении постоянства состава внутренней среды организма.
За счет связей с проприорецепторами продолговатый мозг выполняет роль регулятора тонуса скелетной мускулатуры, прежде всего обеспечивая тоническое напряжение мышц разгибателей, предназначенных для преодоления силы тяжести организма.
Продолговатый мозг регулирует работу спинного мозга. Эта регуляция направлена на функциональное объединение всех сегментов спинного мозга, на обеспечение условий для целостной его деятельности. Продолговатый мозг осуществляет более тонкие формы приспособительных реакций организма к внешней среде по сравнению со спинным мозгом.
К образованиям среднего мозга относят ножки мозга, ядра III (глазодвигательный) и IV (блоковый) пар черепных нервов, пластинку крыши (четверохолмие), красные ядра и черное вещество. В ножках мозга проходят восходящие и нисходящие нервные пути.
В строении среднего мозга полностью утрачиваются сегментарные признаки. В среднем мозге клеточные элементы образуют сложные скопления в виде ядер. Ядерные образования относятся непосредственно к среднему мозгу, а также к входящей в его состав ретикулярной формации.
Передние бугры пластинки крыши получают импульсы от сетчатой оболочки глаз. В ответ на эти сигналы осуществляется регуляция просвета зрачка и аккомодация глаза. Аккомодация — приспособление глаза к ясному видению разноудаленных предметов за счет изменения кривизны хрусталика.
Задние бугры пластинки крыши получают импульсы от ядер слуховых нервов, расположенных в продолговатом мозге. Благодаря этому происходит рефлекторная регуляция тонуса мышц среднего уха, а у животных — поворот ушной раковины к источнику звука. Таким образом, при участии передних и задних бугров четверохолмия осуществляются установочные, ориентировочные рефлекторные реакции на световые и звуковые раздражения (движения глаз, поворот головы и даже туловища в сторону светового или звукового раздражителя). При разрушении ядер четверохолмия зрение и слух сохраняются, но исчезают ориентировочные реакции на свет и звук.
С деятельностью бугров четверохолмия тесно связаны функции ядер III к IV пар черепных нервов, возбуждение которых определяет движение глаз вверх, вниз, в стороны, а также сведение (конвергенция) и разведение глазных осей при переносе взора с удаленных предметов на близкие и обратно.
Красные ядра участвуют в регуляции мышечного тонуса и в проявлении установочных рефлексов, обеспечивающих сохранение правильного положения тела в пространстве. При отделении заднего мозга от среднего тонус мышц-разгибателей повышается, конечности животного напрягаются и вытягиваются, голова запрокидывается. Следовательно, у здорового животного и человека красные ядра несколько притормаживают тонус мышц-разгибателей.
Черное вещество также регулирует мышечный тонус и поддержание позы, участвует в регуляции актов жевания, глотания, артериального давления и дыхания, то есть деятельность черного вещества, как и красных ядер, тесно связана с работой продолговатого мозга.
Таким образом, средний мозг регулирует тонус мышц, что является необходимым условием координированных движений. Средний мозг регулирует ряд вегетативных функций организма (жевание, глотание, артериальное давление, дыхание). За счет среднего мозга расширяется (становится многообразнее) рефлекторная деятельность организма (появляются ориентировочные рефлексы на звуковые и зрительные раздражения).
Образования ствола мозга принимают участие в правильном распределении тонуса между отдельными группами мышц. Рефлексы, обеспечивающие мышечный тонус, получили название тонических. В осуществлении этих рефлексов участвуют мотонейроны спинного мозга, вестибулярные ядра продолговатого мозга, мозжечок, образования среднего мозга (красные ядра). В целостном организме проявление тонических рефлексов контролируется клетками моторной зоны коры больших полушарий.
Тонические рефлексы возникают при изменении положения тела и головы в пространстве за счет возбуждения проприорецепторов мышц, рецепторов вестибулярного аппарата внутреннего уха и тактильных рецепторов кожи.
Тонические рефлексы делят на две группы: статические и статокинетические. Статические рефлексы возникают при изменении положения тела, особенно головы, в пространстве. Статокинетические рефлексы проявляются при перемещении тела в пространстве, при изменении скорости движения {вращательного или прямолинейного).
Таким образом, тонические рефлексы предотвращают возможность нарушения равновесия, потерю активной позы и способствуют восстановлению нарушенной позы.
Промежуточный мозг — часть переднего отдела ствола мозга. Основными образованиями промежуточного мозга являются зрительные бугры (таламус) и подбугровая область (гипоталамус).
Зрительные бугры — массивное парное образование, они занимают основную массу промежуточного мозга. Наибольших размеров и наивысшей сложности строения зрительные бугры достигают у человека.
Зрительные бугры являются центром всех афферентных импульсов. Через зрительные бугры к коре головного мозга поступает информация от всех рецепторов нашего организма, за исключением обонятельных. Кроме того, от зрительных бугров нервные импульсы передаются к различным образованиям ствола мозга. В зрительных буграх обнаружено большое количество ядерных образований. Функционально их можно разделить на 2 группы: специфические и неспецифические ядра.
Специфические ядра получают информацию от рецепторов, перерабатывают ее и передают в определенные области коры большого мозга, где возникают соответствующие ощущения (зрительные, слуховые и т. д.).
Неспецифические ядра не имеют прямой связи с рецепторами организма. Они получают импульсы от рецепторов через большое количество переключений (синапсов). Импульсы от этих образований через подкорковые ядра поступают к множеству нейронов, расположенных, в различных областях коры большого мозга, вызывая повышение их возбудимости.
При повреждении зрительных бугров у человека наблюдается полная потеря чувствительности или ее снижение на противоположной стороне, выпадает сокращение мимической мускулатуры, которое сопровождает эмоции, могут возникать расстройства сна, понижение слуха, зрения и т. д.
Гипоталамическая (подбугровая) область участвует в регуляции различных видов обмена веществ (белков, жиров, углеводов, солей, воды), регулирует теплообразование и теплоотдачу, состояние сна и бодрствования. В ядрах гипоталамуса происходит образование ряда гормонов, которые затем депонируются в задней доле гипофиза. Передние отделы гипоталамуса являются высшими центрами парасимпатической нервной системы, задние — симпатической нервной системы. Гипоталамус участвует в регуляции многих вегетативных функций организма.
К подкорковым, или базальным, ядрам относят три парных образования: хвостатое ядро, скорлупу чечевицеобразного ядра и бледный шар. Базальные ядра расположены внутри больших полушарий, в нижней их части, между лобными долями и промежуточным мозгом. Развитие и клеточное строение у хвостатого ядра и скорлупы одинаковы. Поэтому их рассматривают как единое образование — полосатое тело.
Полосатое тело регулирует сложные двигательные функции, безусловнорефлекторные реакции цепного характера: бег, плавание, прыжки. Эту функцию полосатое тело осуществляет через бледный шар, притормаживая его деятельность. Кроме того, полосатое тело через гипоталамус регулирует вегетативные функции организма, а также вместе с ядрами промежуточного мозга обеспечивает осуществление сложных безусловных рефлексов цепного характера — инстинктов.
Бледный шар является центром сложных двигательных рефлекторных реакций (ходьба, бег), формирует сложные мимические реакции, участвует в обеспечении правильного распределения мышечного тонуса. Свои функции бледный шар осуществляет через образования среднего мозга (красные ядра и черное вещество). При раздражении бледного шара наблюдается общее сокращение скелетных мышц противоположной стороны тела. При поражении бледного шара движения теряют свою плавность, становятся неуклюжими, скованными.
Использованная литература
1. Физиология / Под ред. С.А. Георгиевой. – 2-е изд. – Ф48 М.: Медицина, 1986. – 400 с.