Физиология дыхания
Министерство образования Российской Федерации
Пензенский Государственный Университет
Медицинский Институт
Кафедра Терапии
Реферат
на тему:
«Физиология дыхания»
Пенза, 2008
План
Вентиляция легких и легочные объемы
Легочное кровообращение
Вентиляционно-перфузионные отношения
Литература
1. Вентиляция легких и легочные объемы
Вентиляция легких - это смена воздуха в легких, совершаемая циклически при вдохе и выдохе. Легочную вентиляцию характеризуют прежде всего четыре основных легочных объема: дыхательный, резервный объем вдоха, резервный объем выдоха и остаточный объем. Вместе они составляют общую емкость легких. Сумма всех перечисленных объемов равняется максимальному объему, до которого могут быть расправлены легкие. Легочные емкости представляют собой сумму двух и более объемов.
Объем воздуха в легких и дыхательных путях зависит от конституционально-антропологических и возрастных характеристик человека, свойств легочной ткани, поверхностного натяжения альвеол, а также силы, развиваемой дыхательными мышцами. Для оценки вентиляционной функции легких, состояния дыхательных путей, изучения паттерна дыхания применяют различные методы исследования: пневморафию, спирометрию, спирографию и др. С помощью спирографа можно определить и записать величины легочных объемов воздуха, проходящих через воздухоносные пути человека.
При спокойном вдохе и выдохе через легкие проходит сравнительно небольшой объем воздуха (около 500 мл), который принято называть дыхательным объемом (ДО).
При форсированном (глубоком) вдохе человек может дополнительно вдохнуть еще определенный объем воздуха. Этот резервный объем вдоха (РО>вд>) - максимальный объем воздуха, который способен вдохнуть человек после спокойного вдоха. Величина резервного объема вдоха составляет у взрослого человека примерно 1,8-2,0 л.
После спокойного выдоха человек может дополнительно выдохнуть еще определенный объем воздуха. Это резервный объем выдоха (РО>выд>), величина которого составляет в среднем 1,2 - 1,4 л.
Объем воздуха, который остается в легких после максимального выдоха и в легких мертвого человека - остаточный объем легких (ОО). Величина остаточного объема составляет 1,2 -1,5 л.
Различают следующие емкости легких.
1. Общая емкость легких (ОЕЛ) - объем воздуха, находящегося в легких после максимального вдоха.
2. Жизненная емкость легких (ЖЕЛ) включает в себя дыхательный объем, резервный объем вдоха, резервный объем выдоха. ЖЕЛ - это объем воздуха, выдохнутого из легких после максимального вдоха при максимальном выдохе (ЖЕЛ = ОЕЛ – ОО); ЖЕЛ составляет у мужчин 3,5 - 5,0 л, у женщин - 3,0-4,0 л.
3. Емкость вдоха (Евд) равна сумме дыхательного объема и резервного объема вдоха, составляет в среднем 2,0 - 2,5 л.
4. Функциональная остаточная емкость (ФОЕ) - объем воздуха в легких после спокойного выдоха. В легких при спокойном вдохе и выдохе постоянно содержится примерно 2500 мл воздуха, заполняющего альвеолы и нижние дыхательные пути. Благодаря этому газовый состав альвеолярного воздуха сохраняется на постоянном уровне.
Объем легочной вентиляции удобно определять как объем газа, поступающего в дыхательные пути и покидающего их за определенный отрезок времени. Минутный объем дыхания (МОД) - объем воздуха, проходящий через легкие за одну минуту. Величина легочной вентиляции определяется глубиной дыхания и частотой дыхательных движений. В покое частота дыхательных движений человека составляет примерно 16 в мин ("эйпноэ" или "хорошее дыхание"), а объем выдыхаемого воздуха - около 500 мл. Умножив частоту дыхания в минуту на величину дыхательного объема, получим МОД, который у человека в покое составляет в среднем 8 л/мин. При этом акт вдоха проходит несколько быстрее, чем акт выдоха.
Максимальная вентиляция легких (МВЛ) - объем воздуха, который проходит через легкие за одну минуту во время максимальных по частоте и глубине дыхательных движений. Максимальная вентиляция возникает во время интенсивной работы, при недостатке содержания О>2> (гипоксия) и избытке СО>2> (гиперкапния) во вдыхаемом воздухе. В этих условиях МОД может достигать 150 - 200 л/мин.
Прежде чем поток вдыхаемого воздуха достигнет альвеолярно-капиллярной мембраны, он проходит по огромному количеству ВП, расположенных между насосом или ротовой полостью и альвеолами. Распределение дыхательного объема по проводящим ВП и газообменивающим зонам оказывает глубокое влияние на дыхательную функцию. В газообмене может участвовать только та часть дыхательного объема воздуха, которая попадает в нормально перфузируемые альвеолы. Поэтому наиболее важным показателем легочной вентиляции является объем альвеолярной вентиляции (V>А>). Остальная часть общего объема является вентиляцией так называемого мертвого пространства (V>D>). Объем альвеолярного пространства можно вычислить по формуле:
V>A> = ЧД (V>T>-V>D>).
Мертвое пространство включает объем дыхательных путей, в которых не происходит газообмен (анатомическое мертвое пространство), и объем неперфузируемых альвеол (альвеолярное мертвое пространство). Сумма анатомического и альвеолярного мертвого пространства называется физиологическим мертвым пространством. В норме у взрослого человека при вертикальном положении тела мертвое пространство равно 150 мл (примерно 2 мл/кг) и практически состоит только из анатомического мертвого пространства. Надо иметь в виду, что фактический объем анатомического мертвого пространства может варьировать в зависимости от способа осуществления ИВЛ (через маску, мундштук, трахеальную трубку, трахеостомическую канюлю) и от объема используемых указанных деталей.
У здорового человека в состоянии покоя альвеолярное мертвое пространство весьма невелико, но при некоторых патологических состояниях - при гиповолемии, легочной эмболии или эмфиземе могут возникать очаги - зоны альвеолярного мертвого пространства. Основной причиной увеличения отношения мертвого пространства к дыхательному объему в таких ситуациях является значительное снижение или полное отсутствие кровотока в вентилируемых альвеолах:
V>D>/V>T> = Paco>2> - Peco>2>/ Paco>2>.
Уравнение, известное как уравнение Бора, показывает, что отношение мертвого пространства к дыхательному объему может быть рассчитано как частное от деления разности PCO>2> альвеолярного и выдыхаемого воздуха (PeCO>2>) на альвеолярное PCO>2>. Поскольку альвеолярное PCO>2> практически совпадает с артериальным PCO>2>, V>D>/V>T> может быть рассчитано с помощью одновременного измерения PCO>2> в пробах артериальной крови (PaCO>2>) и выдыхаемого газа.
Скорость образования СО>2> (VCO>2>) у здорового человека массой 70 кг в состоянии покоя составляет около 200 мл в 1мин. Альвеолярное напряжение углекислого газа (РАСО>2>) отражает баланс между общей выработкой (продукцией) углекислого газа (VCO>2>) и альвеолярной вентиляцией (элиминацией СО>2>):
P>A>CO>2> = КVCO>2>/V>A>,
где: V>A> - альвеолярная вентиляция,
К - константа, равная 0,863.
Система регуляции дыхания «установлена» на поддержание PCO>2> в артериальной крови на уровне 40 мм рт. ст. Уравнение показывает, что при постоянной скорости образования двуокиси углерода РCO>2> изменяется обратно пропорционально альвеолярной вентиляции.
Подобно тому, как РаCO>2> определяется балансом между продукцией СО>2> и альвеолярной вентиляцией, альвеолярное РO>2> является функцией скорости поглощения кислорода через альвеолярно-капиллярную мембрану и альвеолярной вентиляции. Поскольку парциальные давления азота и водяного пара в альвеолах постоянны, Р>А>О>2> и Р>А>СО>2> изменяются реципроктно по отношению друг другу в зависимости от изменений альвеолярной вентиляции.
Сумма парциальных давлений О>2>, СО>2>, N>2> и водяного пара постоянна, парциальное давление О>2> либо СО>2> может быть рассчитано, если одно из них известно. Расчет основывается на уравнении альвеолярного газа:
Р>АО2 >= Р>IО2> - Р>АСО2>(FiO>2> + 1-FiO>2>/R) ,
где: РiО>2> – давление кислорода во вдыхаемом газе,
FiO>2> - фракционная концентрация О2 во вдыхаемом газе,
R - дыхательное газообменное соотношение.
R- дыхательное газообменное соотношение, выражает скорость выделения СО>2>, относительно скорости поглощения О>2>, т.е. R =VCO>2>/VO>2>. В устойчивом состоянии организма дыхательное газообменное соотношение равно дыхательному коэффициенту (RQ), который описывает отношение продукции двуокиси углерода к потреблению кислорода на клеточном уровне. Это соотношение зависит от того, что преимущественно используется в организме в качестве источников энергии - углеводы или жиры. В процессе метаболизма 1г углеводов выделяется больше СО>2>.
Легкие довольно разнородны с точки зрения региональных механических свойств ВП и паренхимы, что является причиной неравномерного распределения в них воздуха. У здорового человека в положении стоя имеется градиент плеврального давления между верхушкой и основанием легкого. Плевральное давление наибольшее (т.е. наиболее отрицательное) у верхушки легких и наименьшее (т.е. наименее отрицательное) у основания. Градиент составляет около 0,25 см вод. ст. на каждый сантиметр высоты. Поскольку транспульмональное давление равно Рalv-Ppl, то его величина на верхушке больше, чем у основания легких. В результате альвеолы из различных зон оказываются в разных точках кривой «давление-объем». Альвеолы в верхних отделах легких из-за более высокого транспульмонального давления расправлены почти максимально, относительно нерастяжимы и значительно меньше увеличиваются в объеме во время вдоха. И наоборот, альвеолы в нижних отделах легких благодаря более низкому транспульмональному давлению более растяжимы и больше увеличиваются во время вдоха. Иными словами, при низком исходном объеме легкие легче раздуть, чем при высоком, так как по мере их расширения падает растяжимость. Статическая растяжимость выше в нижних областях легких, они занимают больший объем и, соответственно, там распределяется большая часть дыхательного объема. Фактически данное обстоятельство повышает эффективность газообмена, поскольку легочный кровоток также преобладает у основания легких.
Сопротивление дыхательных путей также способствует возникновению регионарных различий в легочной вентиляции. Конечный альвеолярный объем при вдохе определяется исключительно растяжимостью только в том гипотетическом случае, если время вдоха не ограничено. В действительности же время вдоха лимитировано частотой дыхания и временем, необходимым для выдоха. Следовательно, слишком короткое время вдоха не позволит альвеолам достичь ожидаемого объема. В связи с наличием различия в податливости различных альвеол, а также резистентности дыхательных путей в различных зонах легкого, регионарное время заполнения альвеол различное.
Расправление легких во время вдоха можно описать математически с использованием постоянной времени – «τ»:
τ = C> >> >R = 0,1 л/см 2 см Н>2>О/ (л с) = 0,2 с.
Время, соответствующее 1 τ - это время, необходимое для расправления альвеолы приблизительно на 60 % от максимального объема. Расправление на 99 % требует времени, равного 4τ.
Регионарное снижение растяжимости легких или увеличение резистентности дыхательных путей приводит к возрастанию неравномерности вентиляции, а также могут стать причиной асинхронного заполнения альвеол во время вдоха. Некоторые альвеолы продолжают заполняться и тогда, когда из других альвеол газ уже начал выходить.
2. Легочное кровообращение
Малый круг кровообращения представляет собой контур большого объема с низким сопротивлением, «функционально» расположенный между правой и левой половинами сердца. В отличие от кровообращения любого другого органа, малый круг должен вмешать весь объем сердечного выброса как в состоянии покоя, так и при напряжении. Из 5 л крови, протекающих через легкие за одну минуту, в легочных капиллярах одномоментно находится и участвует в газообмене только 70-100 мл. Этот небольшой объем крови образует на альвеолокапиллярной мембране пленку площадью 50-100 м2 и толщиной в один эритроцит. Емкость капилляров легких относительно постоянна, но общий внутрилегочный объем крови может изменяться от 500 до 1000 мл. Внутрилегочный объем крови незначительно увеличивается при каждом вдохе (при самостоятельном дыхании) и во время сердечной систолы.
Большинство легочных вен обладают тонкими сжимаемыми и растяжимыми стенками, легко поддающимися воздействию как внутрисосудистого, так и внесосудистого давлений. При напряжении происходит расширение функционирующих и включение в циркуляцию ранее нефункционирующих сосудов. Это позволяет сосудистому руслу легких пропускать возросший сердечный выброс при весьма незначительном повышении давления в легочной артерии. У здорового взрослого человека в состоянии покоя на уровне моря среднее давление в легочной артерии обычно составляет 9-15 мм рт. ст. Систолическое и диастолическое давление - от 15 до 25 мм рт. ст. и от 5 до 10 мм рт. ст. соответственно.
Увеличение сердечного выброса или давления в легочной артерии сопровождается снижением легочного сосудистого сопротивления. Это снижение нелинейно: его степень зависит от вазомоторного тонуса, существовавшего до повышения сердечного выброса, а также от уровня давления в легочной артерии и величины кровотока.
Легочный кровоток так же неравномерен, как и вентиляция. В частности, на его распределение влияют изменение позы и физическая нагрузка. В нижерасположенные отделы легких поступает крови больше, чем в вышерасположенные. Например, при вертикальной позиции кровоток минимален в верхушках, в положении лежа на спине – в вентральных отделах легких. Неравномерное распределение легочного кровотока можно объяснить различиями гидростатического давления в кровеносных сосудах. Если рассматривать легочную артериальную систему как единый столб крови высотой около 30 см, то разница в давлении между его вершиной и основанием составит примерно 30 см вод. ст., или 23 мм рт. ст. Поскольку в малом круге давление крови невелико, такая разница довольно существенна. В результате действия силы тяжести создается градиент внутрисосудстого давления, составляющий 1 см вод. ст. на каждый сантиметр высоты легкого.
При умеренной физической нагрузке кровоток в верхних и нижних отделах увеличивается, и регионарные различия его распределения сглаживаются.
В каждом легком условно принято выделять три зоны - в зависимости от соотношения альвеолярного (Рalv), артериального (Ра) и венозного (Pv) давлений.
Первая зона (верхняя) определяется как область, где альвеолярное давление (Раlv) больше давления в легочной артерии (Рра), которое в свою очередь, превосходит легочное венозное давление (РalvPaPv). Эта зона находится в верхних отделах легких и представляет собой альвеолярное мертвое пространство, потому что здесь давление в альвеолах сжимает легочные капилляры, а кровоток минимальный. Верхушка вертикально расположенного легкого перфузируется только за счет пульсирующего характера кровотока в легочной артерии.
В средней зоне (зона 2) давление в легочной артерии превышает альвеолярное, которое в свою очередь больше, чем давление в венозном русле малого круга кровообращения (PpaPalvPpv). Вследствие этого кровоток не определяется обычным градиентом между средним давлением в легочной артерии и альвеолярным давлением. Изменяющиеся отношения между альвеолярным и сосудистым давлениями попеременно смещают давление оттока в диапозоне между альвеолярным и венозным, создавая так называемый резистор Старлинга. Следовательно, кровоток через капилляры зоны 2 выглядит прерывистым благодаря каналам, которые открыты, когда венозное давление превышает альвеолярное, и закрыты, когда альвеолярное давление превосходит легочное венозное.
В зоне 3 легочный капиллярный кровоток непрерывен и определяется артериально-венозным градиентом давления. В этом случае обычные расчеты легочного сосудистого сопротивления становятся правомерными.
При малом объеме легких большое значение имеет сопротивление внеальвеолярных сосудов. В этих условиях снижается регионарный кровоток, причем преимущественно в области оснований легких, где легочная паренхима расправлена слабее всего. Это дает основание говорить и о четвертой зоне, в которой сопротивление кровотоку создают, как полагают, экстраальвеолярные, а не альвеолярные сосуды. Она исчезает с глубоким вдохом предположительно из-за выпрямления этих сосудов при расправлении легких.
Описанные зоны являются функциональными, а не анатомическими структурами. Поверхности, разделяющие зоны не закреплены топографически и перемещаются по вертикали легких в соответствии с изменениями отношений между легочным артериальным, легочным венозным и альвеолярными давлениями.
В регуляции сосудистого тонуса принимают участие, как вегетативная нервная система, так и местные факторы, причем последние более значимы. Наиболее мощный стимул легочной вазоконстрикции – это гипоксия (в противоположность сосудорасширяющему действию гипоксии в большом круге кровообращения). Гипоксическая легочная вазоконстрикция, как реакция на альвеолярную гипоксию, приводит к заметному сужению прилегающих к альвеолам прекапиллярных мышечных легочных артерий и артериол. Вазоконстрикция происходит как при гипоксии в легочной артерии (в смешанной венозной крови), так и при альвеолярной гипоксии. Однако стимулирующий эффект последней более выражен. Медиаторы, вовлеченные в гипоксическую легочную вазоконстрикцию, пока полностью не идентифицированы. Этот феномен возникает либо благодаря прямому действию гипоксии на легочные сосуды, либо за счет преобладания выработки сосудосуживающих лейкотриенов над продукцией сосудорасширяющих простагландинов (простагландин I>2>). Возможно, гипоксия подавляет образование оксида азота (NO). В случае локальной альвеолярной гипоксии (как при пневмонии или региональном ателектазе) легочная вазоконстрикция тоже локализуется, отводя кровоток от гипоксических областей и уменьшая степень вентиляционно-перфузионного несоответствия. Однако при диффузном характере альвеолярной гипоксии возникающая легочная вазоконстрикция воздействует на легкое в целом. Например, при хронической обструктивной болезни легких гипоксическая легочная вазоконстрикция является компонентом повышения сосудистого сопротивления легких.
Гипероксия не оказывает существенного влияния на легочное кровообращение у здоровых людей. Значительная ацидемия (рН7,2) вызывает легочную вазоконстрикцию. У человека ацидемия действует синергически с гипоксией. Значительная алкалемия (рН7,5) уменьшает сужение сосудов в ответ на гипоксию. В отличие от гипоксемиии, гиперкапния, по-видимому, способствует легочной гипертензии, вызывая ацидемию, а не прямую вазоконстрикцию.
3. Вентиляционно-перфузионные отношения
Количественная связь между вентиляцией и перфузией выражается вентиляционно-перфузионным отношением. В норме альвеолярная вентиляция (V) составляет 4 л/мин, легочный капиллярный кровоток (Q) - 5 л/мин, а их соотношение V/Q - соответственно 0,8. Апикально-базальные градиенты кровотока и вентиляции совпадают по направлению, но величина изменений вентиляции и перфузии от верхушки к основанию различна. Кровоток к основанию возрастает в большей степени, поэтому в апикальных отделах легких V/Q выше, чем в базальных. Альвеолы с V/Q менее единицы обеспечивают нормальную разницу между идеальным средним Р>А>О>2 >и Р>а>О>2>, т.е. альвеолярно-артериальную разницу по кислороду. В норме эта разница составляет от 5 до 10 мм рт. ст. у здоровых молодых людей, и может возрасти до 20 мм рт. ст. у здоровых пожилых людей.
Грубое нарушение соответствия вентиляции и перфузии сопровождается развитием дыхательной недостаточности. Глубина патофизиологических воздействий на артериальную оксигенацию нарушенных вентиляционно-перфузионных отношений намного превосходит воздействия других механизмов развития гипоксемии, включая гиповентиляцию, диффузионный блок и шунтирование.
Для отдельной легочной единицы (комплекс "альвеола-капилляр") V/Q может варьировать от 0 (отсутствие вентиляции) до бесконечности V/Q= (отсутствие кровотока). Первое состояние представляет собой внутрилегочный шунт, второе - альвеолярное мертвое пространство.
В физиологии дыхания под шунтированием понимают возврат десатурированной смешанной венозной крови из правых отделов сердца в левые без насыщения кислородом в легких. Этот тип шунта обозначают как шунт «справа-налево», он приводит к снижению ("разбавлению") содержания кислорода в артериальной крови. Физиологический шунт - понятие, используемое для количественного описания воздействия как абсолютного (анатомического) шунта, так и зон с низким V/Q. В норме физиологический шунт составляет менее 5 % сердечного выброса. Под абсолютными шунтами понимают анатомические шунты и те легочные единицы, где V/Q равно нулю. Относительный шунт - участок легкого с низким, но не нулевым значением V/Q. С практической точки зрения, гипоксемию, обусловленную относительным шунтом, можно частично корригировать, увеличив концентрацию кислорода во вдыхаемой смеси. При абсолютном шунте гипоксемию таким способом уменьшить нельзя.
Физиологический шунт рассчитывают, использую уравнение шунта:
Qs/Qt = (Сс'О>2>- СаО>2>)/(Сс'О>2>- CvO>2>),
где: Qs/Qt - фракция шунтируемой крови,
Сс'О>2> - содержание кислорода в легочной капиллярной крови,
СаО>2>; - содержание кислорода в артериальной крови,
CvO>2> — содержание кислорода в смешанной венозной крови.
Фракцию шунтируемой крови (Qs/Qt) можно рассчитать в клинических условиях, если измерить парциальное давление кислорода и насыщение гемоглобина кислородом в артериальной и смешанной венозной крови. Для получения образца смешанной венозной крови необходима катетеризация легочной артерии. Для вычисления парциального давления кислорода в крови конечных легочных капилляров используют уравнение альвеолярного газа. Принято, что при FiO>2> > 0,21 кровь конечных легочных капилляров насыщена кислородом на 100 %.
Хотя альвеолярные единицы с низким V/Q обычно вызывают уменьшение PaO>2>, в редких случаях они способствуют повышению PaCO>2>. Это явление не связано с большей растворимостью СО>2> по сравнению с О>2>, поскольку в норме диффузионных препятствий поглощению О>2> не существует. Люди с умеренным увеличением фракции альвеол с высоким V/Q без труда повышают общую вентиляцию и выводят больше СО>2> из других легочных единиц, компенсируя этим дополнительное мертвое пространство. Этот эффект возможен благодаря линейности кривой диссоциации СО>2>-гемоглобина (т.е. содержание СО>2> в крови линейно связано с PaCO>2)>.
Подобного компенсаторного ответа на гипоксемию, возникающую из-за наличия альвеол с низким V/Q, не существует. Хотя увеличение общей вентиляции повышает P>А>O>2> в альвеолах с V/Q1, увеличение содержания О>2> в конечно капиллярной крови минимально. Оно не компенсирует вклад десатурированной крови, оттекающей от альвеол с низким V/Q.
Теоретически физиологический эффект альвеол с V/Q больше единицы, но не с бесконечной величиной, может быть описан как функциональный эквивалент дополнительного мертвого пространства, т.е. «альвеолярного мертвого пространства». Воздействие единиц с высоким V/Q рассчитывается с помощью уравнения Бора.
Обмен газов в легких. Кинетическая энергия всех молекул атмосферного газа создает атмосферное или барометрическое давление. Давление плевральное или альвеолярное обычно выражают относительно атмосферного и называют манометрическим давлением.
Атмосферный воздух представляет собой смесь газов: азота, кислорода, углекислого газа, водяных паров. Количество аргона и двуокиси углерода в нем очень мало, а давление водяного пара при нормальных условиях окружающей среды невелико. Поэтому в практических целях атмосферный воздух можно рассматривать как смесь 21% кислорода и 79% азота (FiO>2> – 0,21 и FiN>2> - 0,79 - фракционные концентрации кислорода и азота соответственно).
В газовой смеси кинетическая энергия каждого газа создает давление, известное как парциальное давление газа. Смесь газов, находящаяся в резервуаре, оказывает на его стенки общее давление, равное сумме парциальных давлений всех газов газовой смеси (закон Дальтона).
Давление водяного пара во вдыхаемом воздухе, который нагревается до температуры тела и полностью увлажняется, составляет 47 мм рт.ст. Обычно фракционную концентрацию газа рассчитывают после вычитания давления водяного пара (т.е. как «сухой газ»). Парциальное давление газа представляет собой произведение его фракционной концентрации и общего давления «сухой» смеси.
Движение газа через альвеолярно-капиллярную мембрану происходит путем диффузии, согласно закону Фика. В соответствии с этим законом скорость переноса газа через тканевую поверхность или «мембрану» прямо пропорциональна разнице парциального давления газа по обе стороны мембраны и константе мембраны, известной как диффузионная способность (Dм):
V>G >= Dм (P>1>-P>2>),
где: V>G >- скорость переноса газа через тканевую поверхность,
P>1 >- парциальное давление газа по одну сторону тканевой поверхности,
P>2>- парциальное давление газа по другую сторону тканевой поверхности.
Диффузионная способность, в свою очередь, состоит из нескольких компонентов, включая растворимость газа в ткани (), площадь тканевой поверхности (А), ее толщину (d) и молекулярную массу газа (МВ):
Dм = k
A/d
/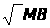 ,
,
где: k - константа.
Соединяя обе формулы, получаем:
V>G>
= k
A/d
/
(P>1>-P>2>).
Из последнего уравнения следует, что для данного газа скорость его диффузии через альвеолярно-капиллярную мембрану возрастает: а) с увеличением площади поверхности мембраны, растворимости и градиента давления газа по обе стороны мембраны; б) с уменьшением толщины мембраны и молекулярного веса газа.
Не следует представлять диффузионную способность легких (Dм) как некую константную максимальную величину, характеризующую исключительно «пропускную способность» альвеолокапиллярной мембраны. Прежде всего, это конкретная величина, относящаяся к данным условиям функционирования легких, а, кроме того, она характеризует диффузионную способность не только альвеоло-капиллярной мембраны, но всей легочной системы в целом.
Большая общая площадь и малая толщина (0,4-0,5 мкм) альвеолярно-капиллярной мембраны создают чрезвычайно благоприятные условия для диффузии кислорода. Усиленное связывание кислорода с гемоглобином при насыщении свыше 80 % также способствует его диффузии. В нормальных условиях покоя время транзита эритроцита через легочной капилляр составляет около 0,75 с. Кровь, протекающая по легочному капилляру, полностью оксигенируется за время, в течение которого эритроцит преодолевает одну треть длины капилляра. Напряжение кислорода в смешанной венозной крови поднимается с 40 мм рт.ст. до величины альвеолярного 100 мм рт.ст.
Связывание кислорода с гемоглобином — главный фактор, ограничивающий скорость перехода О>2 >из альвеолярного газа в кровь. Поэтому диффузионная способность легких зависит не только от диффузионных свойств альвеолярно-капиллярной мембраны, но также и от величины легочного кровотока. В норме поглощение кислорода кровью лимитируется главным образом скоростью легочного кровотока, а не его диффузией через альвелярно-капиллярную мембрану.
Нарушения диффузии возникают редко. Их причиной могут стать: расстройства вентиляционно-перфузионных отношений; обширная деструкция альвеолярно-капиллярной мембраны с увеличением так называемого расстояния диффузии (утолщение мембран при фиброзе легких, склерозе легочных сосудов, отеке легких, интерстициальной пневмонии и др.); уменьшение поверхности, через которую осуществляется диффузия (резекция легочной ткани). Нарушение транспорта кислорода вследствие недостаточного времени капиллярного транзита усиливаются при повышенном потреблении кислорода и увеличении сердечного выброса (например, при физической нагрузке).
Поскольку растворимость СО>2> в тканях примерно в 20 раз больше, чем О>2>, то и скорость диффузии СО>2 >через альвеолярно-капиллярную мембрану в 20 раз выше. Поэтому система в целом располагает значительными резервами относительно диффузии СО>2>, и незначительные изменения в состоянии паренхимы легких не сопровождаются появлением артериально-альвеолярного градиента СО>2>.
Литература
«Неотложная медицинская помощь», под ред. Дж. Э. Тинтиналли, Рл. Кроума, Э. Руиза, Перевод с английского д-ра мед. наук В.И.Кандрора, д. м. н. М.В.Неверовой, д-ра мед. наук А.В.Сучкова, к. м. н. А.В.Низового, Ю.Л.Амченкова; под ред. Д.м.н. В.Т. Ивашкина, Д.М.Н. П.Г. Брюсова; Москва «Медицина» 2001
2. Интенсивная терапия. Реанимация. Первая помощь: Учебное пособие / Под ред. В.Д. Малышева. — М.: Медицина.— 2000.— 464 с.: ил.— Учеб. лит. Для слушателей системы последипломного образования.— ISBN 5-225-04560-Х