Культуры изолированных клеток и тканей как новый источник для получения лекарственного сырья
Федеральное агентство по образованию
Государственное образовательное учреждение высшего профессионального образования
Петрозаводский государственный университет
Медицинский факультет
Курсовая работа
по фармакогнозии
«Культуры изолированных клеток и тканей как новый источник для получения лекарственного сырья».
Панкрашкина Полина
3 курс 320 группа
Руководитель:
К.б.н., ст. преподаватель
Морозова К.В.
Петрозаводск 2007г.
Содержание
Введение
1. История создания культуры клеток и тканей
2. Культивирование растительных клеток и тканей
2.1 Понятие культуры клеток и тканей
2.2 Синтез вторичных метаболитов
2.3 Влияние генетических, физических и химических факторов на рост и развитие культур и синтез вторичных метаболитов
3. Культура ткани растений и синтез вторичных метаболитов
3.1 Образование полифенолов в культуре ткани чайного растения
3.2 Образование -карболиновых алкалоидов в культуре ткани гармалы обыкновенной
3.3 Накопление алкалоидов в культуре ткани раувольфии змеиной
3.4 Алкалоиды каллусных клеток Мака прицветного
3.5 Образование вторичных метаболитов в культуре ткани рода Rutaceae
Заключение
Литература
Введение
Растения являются незаменимым источником получения очень многих практически важных веществ. При этом следует подчеркнуть, что промышленное получение некоторых соединений, например, сердечных гликозидов, флавоноидов, кумаринов, эфирных масел достигается только путем выделения их из растительного сырья. Между тем возможности получения так называемых " метаболитов интереса" в достаточном количестве зачастую ограничены. Это связано с сокращением ресурсов некоторых ценных дикорастущих растений, принадлежностью многих лекарственных растений к группам эндемов, редким и исчезающим видам. В связи с этим большой интерес в качестве источника биологически активных веществ представляют культуры растительных клеток.
Цель моего исследования – привлечь внимание к актуальности этой темы, как примеру слияния высоких технологий, науки и экономически выгодного способа получения лекарственного сырья. Сбор дикорастущего сырья наносит вред экологической обстановке региона и всей планете в целом, а при неправильном сборе может и вовсе привести к вырождения зарослей. Культивировать целое растение также не практично если в качестве сырья используется, к примеру, только его корневища или корни. Это значит, что вся надземная часть в лучшем случае идёт на корм животным или просто выбрасывается. Культивирование изолированных клеток и тканей даёт возможность получать именно то сырьё которое необходимо, а кроме того позволяет увеличить количество и качество самих биологически активных веществ.
1. История создания культур клеток и тканей
История развития метода культуры тканей начинается на рубеже XIX-XX вв. с опытов немецких ученых Фехтинга, Рехингера и Хаберландта, которые пытались выращивать на растворах сахарозы изолированные из растений кусочки тканей, группы клеток, волоски. Не достигнув экспериментальных успехов, эти исследователи, однако, высказали ряд важных идей и гипотез, которые были подтверждены значительно позже. В 1947 г. Телл и Готре впервые показали способность синтеза вторичных соединений, а именно алкалоидов, в клеточной культуре белены черной. В нашей стране систематические исследования в этой области были начаты Р.Г. Бутенко в 1957 г. в Институте физиологии растений им. К.А. Тимирязева АН СССР, которая получила клеточные культуры женьшеня и ряда других лекарственных растений. До начала 70-х годов спектр соединений, образуемых клеточными культурами в количествах, характерных для целого растения, был очень ограничен. Это Nicotiana tabacum, в которой некоторые исследователи наблюдали синтез относительно больших количеств никотина (0.7 %), Dioscorea deltoidea, накапливающая до 1.6 % диосгенина, Ammi visnaga, содержащая в 20 раз больше виснагина в культуре ткани, чем в растении, и некоторые другие. Экспериментальные данные, накопившиеся к этому периоду, указывали, что биосинтез многих соединений в недифференцированных тканях сильно подавлен, а появление продуктов во многих случаях было связано с регенерацией корней, побегов и других морфологических структур, т.е. с процессом дифференциации ткани. С начала 70-х годов список фармакологически ценных вторичных продуктов биосинтеза, обнаруженных в культурах тканей, значительно расширился. В 80-е годы на основе метода культуры тканей возникли новые направления биотехнологии, важнейшим из которых была клеточная инженерия — генетическое конструирование новых форм.
2. Культивирование растительных клеток и тканей
Культуры растительных клеток могут синтезировать самые разнообразные по химической природе вещества. Среди них эфирные масла, фенольные соединения, алкалоиды, стероиды, терпеноиды и др. Но несмотря на то, что биомасса культивируемых клеток с начала 80-х годов используется в качестве источника экономически важных продуктов, ряд трудностей и нерешенных вопросов сдерживает широкомасштабное применение культивируемых клеток, обусловливает нерентабельность биотехнологических производств многих ценных видов растений. Содержание практически важных вторичных метаболитов в высших растениях определяется активностью их синтеза, эффективностью транспорта и депонирования в органах запаса растения. Все эти признаки определяются генетически, находятся под контролем развития организма и максимально реализуются в оптимальных внешних условиях.
2.1 Понятие культуры клеток и тканей
В самом общем смысле культура клеток и тканей – это искусственное in vitro индуцирование делений клеток или выращивание в пересадочной культуре тканей, возникших путём пролиферации клеток изолированных сегментов разных частей растения.
Все объекты, культивируемые in vitro, выращиваются стерильными. Стерилизуются исходные кусочки ткани растений (экспланты), питательная среда; антисептически в специальных боксах стерильным инструментом проводятся манипуляции по выращиванию объектов. Сосуды в которых культивируются ткани и клетки, закрываются так, чтобы предотвратить инфицирование в течение продолжительного времени. В культуре тканей лекарственных растений можно выделить три главных направления: получение недифференцированной каллусной массы, создание источников генетического разнообразия форм растений, а так же клеточную селекцию и клональное микроразмножение растений. В природе каллусообразование – естественная реакция на повреждение растений. В культуре изолированных тканей при помещении экспланта (т. е. фрагмента ткани или органа) на питательную среду его клетки дедифференцируются, переходят к делению, образуя однородную недифференцированную массу – каллус. В асептических условиях каллус отделяют и помещают на поверхность агаризованной питательной среды для дальнейшего роста. В результате получают культуру каллусной ткани, которую можно поддерживать неограниченно долго, периодически разделяя её на трансплантаты и пересаживая её на свежую среду. Каллусы легко образуются на эксплантах из различных органов и частей растений: отрезков стебля, листа, корня, проростков семян, фрагментов паренхимы, тканей клубня, органов цветка, плодов, зародышей и т. д. Культивирование каллусных клеток проводят главным образом двумя способами: на агаризованных питательных средах или различных гелеобразующих подложках (силикагель, биогели, полиакриламидные гели, пенополиуретан и др.) и в жидкой питательной среде. В жидкой питательной среде каллус легко распадается на отдельные агрегаты клеток и даёт начало так называемой суспензионной культуре.
Каллусные клетки в культуре in vitro подвержены значительной генетической изменчивости. Изменчивость геномов может приводить к генетическим изменениям у растений-регенерантов, полученных из культуры каллусных клеток, клеточных суспензий или изолированных протопластов. Такие растения получили названия сомаклональных вариантов. Сомаклональные варианты, сохраняя основные свойства прототипа, часто выгодно отличаются от него устойчивостью к болезням, экологическим стрессам, а иногда несколько изменённой биосинтетической способностью и более высокой продуктивностью.
Неотселектированные недифференцированные клетки накапливают, как правило, незначительное, по сравнению с интактным растением, количество веществ специализированного обмена. Только благодаря правильно разработанной стратегии получения высокопроизводительных штаммов к настоящему времени получены культуры тканей, в которых содержание вторичных продуктов достаточно велико, чтобы служить лекарственным сырьем. Однако для многих культур неоднократные попытки различных исследователей определить условия накопления продуктов, характерных для родительских растений, были неудачными. Это касается, в частности, индукции морфинановых алкалоидов в культуре ткани Papaver somniferum, винбластина — в Catharanthus roseus, хинолиновых алкалоидов — в Cinchona ledgeriana, дигоксина — Digitalis lanata и др. Чаще всего в клеточных культурах при длительном культивировании снижается или совсем теряется способность клеток накапливать соединения вторичного метаболизма из-за возникновения малоактивных, но более жизнеспособных вариантов. Снижение биосинтетического потенциала в культуре in vitro происходит из-за подавления дифференциации клеток и их специализации, т.е. в результате потери способности к реализации генетической информации, относящейся ко вторичному обмену.
Важной характеристикой клеточной популяции является ее стабильность в отношении синтеза, транспорта и депонирования метаболитов « интереса». Стабильность может сохраняться в течение всего времени существования популяции. При этом сохраняются и активно работают гены синтеза, системы транспорта и депонирования. Возможен случай постепенного (в течение нескольких лет) увеличения числа клеток со сниженным синтезом метаболитов. И, наконец, в случае полной нестабильности клетки популяции очень быстро теряют свой биосинтетический потенциал. Вопрос о стабильности и нестабильности тесно связан с изучением биологии клеток разных популяций. В организме растения синтез метаболитов, их транспорт и отложение в запас находятся под строгим контролем развития. Часто эти события не только разведены во времени, но и происходят в разных органах растения. Клетка вне организма обычно не транспортирует метаболиты в соседние клетки или в питательную среду, хотя в ряде случаев это явление наблюдается (биосинтез алкалоидов в клеточных культурах мака). На выход вторичных продуктов в культурах растительных клеток влияют многие факторы, однако все способы регуляции вторичного метаболизма в культуре in vitro можно разделить на две группы: физиологическая и генетическая регуляции синтеза вторичных метаболитов.
Подбор физических и химических условий культивирования является наиболее простым и часто применяемым подходом для повышения продуктивности. В основе физиологического регулирования процессов вторичного синтеза лежит изучение влияния факторов культивирования на рост и метаболизм клеток. Большое внимание уделяется таким факторам культивирования, как регуляторы роста, минеральные вещества, витамины, сахара, свет, аэрация, температура, а также иммобилизация клеток и обработка элиситорами. Во многих случаях эти работы привели к успеху, однако они выполняются эмпирически и поэтому длительны и трудоемки. К тому же следует оговориться, что несмотря на эффективность повышения уровня биосинтеза физиологическими методами, добиться количественно значимых изменений в дедифференцированных клеточных культурах, сопоставимых с уровнем в интактном растении, лишь за некоторым исключением, не удается. Стимулирование же синтеза элиситорами носит, к сожалению, временный характер.
Более эффективной в этом плане является генетическая регуляция синтеза вторичного метаболизма в системе in vitro. С использованием экспериментального мутагенеза стало возможным получение довольно продуктивных штаммов. С помощью этого метода в ИФР РАН был получен мутантный штамм Dioscorea deltoidea DM-0.5 (мутаген — N- нитрозометилмочевина, доза — 0.5 ммоль/ч) — сверхпродуцент фуростаноловых гликозидов, высокая способность к синтезу — 6-8 % в сухой массе клеток — сохранялась в течение длительного времени (около 30 лет) [7]. Следует отметить, что метод индуцированного мутагенеза носит также эмпирический характер и не менее трудоемок, чем физиологические способы регуляции вторичного метаболизма. Ряд перспективных культур был получен в результате генетической трансформации и других генно-инженерных манипуляций. Особенно следует отметить трансформанты, полученные с помощью плазмид агробактерий (Agrobacterium rhizogenes A. Tumefaciens), в частности « бородчатых корней», продуктивность которых оказалась достаточно высокой. Поскольку одной из основных причин снижения уровня биосинтеза в культурах in vitro является дедифференциация ткани, то один из путей повышения синтеза вторичных соединений в клеточных культурах связан с дифференцировкой ткани и органогенезом. Повышение содержания вторичных соединений было отмечено в органогенных культурах видов Senecio, Lichroa ledgeriana.
Известно, что физиологическое действие условий in vitro приводит к генетической гетерогенности системы. Речь идет о так называемой сомаклональной изменчивости, которая возникает при длительном культивировании. На генетической изменчивости клеток в культуре in vitro основана селекция штаммов, обеспечивающая большой выход ценных продуктов вторичного метаболизма растительных клеток. При клонировании суспензионной культуры клеток паслена были выделены линии, накапливающие больше 3 % соланидина, получен штамм клеток руты душистой, содержащей в 20 раз больше алкалоида рутакридона по сравнению с растением. Биотехнологическое использование клеточных культур в качестве сырья в промышленных масштабах становится реальностью. В виде примеров можно привести производство шиконина из Lithospermum erythrorhison в Японии (фирма Toshiba) — ценного для косметики, пищевой промышленности и медицины растительного нафтохинонового пигмента. В России производство культуры ткани женьшеня («Биоженьшень») осуществляется на биохимических заводах. Экстракт, получаемый из биомассы женьшеня, используется в качестве биологически активной добавки к кремам, лосьонам, а в пищевой промышленности — для приготовления тонизирующих напитков. Для получения ценного противоаритмического препарата аймалина на ХПХФО «Здоровье»(Харьков, Украина) организовано опытное производство биомассы культуры тканей Rauwolfia serpentina. Таким образом, возможности, открытые методом культуры тканей, позволили в настоящее время создать биотехнологическое производство принципиально новых видов сырья для получения необходимых соединений.
В лаборатории биохимии и биотехнологии растений также получены значительные результаты по получению культур растительных клеток — продуцентов экдистероидов. В начале 90-х годов были получены каллусные культуры Serratula coronata и Ajuga reptans — продуценты экдистероидов. Полученные штаммы различались по степени соответствия интактным растениям по количественному составу экдистероидов и соотношению индивидуальных компонентов. Если в клеточных культурах S. Coronata наблюдали заметное снижение уровня биосинтеза по сравнению с интактными растениями (20-100 раз), то ряд каллусных культур A. Reptans по суммарному содержанию экдистероидов не уступал дикорастущим растениям. Для обеих клеточных культур была отмечена тенденция к снижению уровня синтеза экдистероидов с увеличением продолжительности культивирования, однако были выявлены штаммы и со стабильным уровнем синтеза. Среди длительно культивируемых каллусных культур S. Coronata и A. Reptans были выявлены штаммы с относительно высоким содержанием 20-гидроксиэкдизона (экдистероида, обладающего высоким тонизирующим и ранозаживляющим действием), из которых в 1999 г. нами были получены суспензионные культуры. Методы глубинного культивирования клеток высших растений в последние годы привлекают все больший интерес, поскольку этот метод обладает рядом преимуществ перед поверхностным культивированием (каллусными культурами): обеспечение одинаковых условий для всех клеток популяции; увеличение скорости их роста и биосинтетического потенциала; возможность автоматизации процессов.
Перечень клеточных линий согласно видовой принадлежности
|
ВИД |
ОРГАН или ТКАНЬ |
НАЗВАНИЕ ЛИНИИ |
|
Aristolochia manshuriensis |
Стеблевые сегменты |
A - 2 |
|
Arnebia euchroma |
Пазушная почка |
AE - 1 |
|
Camellia sinensis |
Стебель |
ChS-2 (ЧС-2) |
|
Dioscorea deltoidea |
ИФР Д1,каллус |
IPHR DM 0.5 (ИФР ДМ 0.5) |
|
ИФР Д1,каллус |
IPHR DM1 (ИФР ДМ1) |
|
|
ИФР Д1,каллус |
IPHR DM8 (ИФР ДМ8) |
|
|
Корень |
IPHR D1 (ИФР Д1) |
|
|
Epimedium macrosepalum |
Черешок листа |
EM-1 |
|
Eritrichium incanum |
Корень |
ERSR |
|
Medicago sativa |
Лист |
L-1 (Л-1) |
|
Panax ginseng |
Корень |
DAN-25 (ДАН-25) |
|
Корень |
IPHR G1 (ИФР Ж1) |
|
|
Корень |
PANAX-13 (ПАНАКС-13) |
|
|
Стеблевая опухоль |
R-1 |
|
|
Panax quinquefolius |
Корень |
IPHR G10 (ИФР Ж10) |
|
Poliscias filicifolia |
Лист |
BFT-01-95 (БФТ-01-95) |
|
Rhodiola rosea |
Стебель |
ZK-1 (ЗК - 1) |
|
Rubia cordifolia |
Стеблевой апекс |
RС - 1 |
|
Scorzonera hispanica |
Опухоль корня |
SFR-SH-1 (СФР-SH-1) |
|
Stephania glabra |
линия VILAR Sg-6 |
VILAR Sg-48 (ВИЛАР Sg - 48) |
|
Stevia rebaudiana Bertoni |
Лист |
SR - 1 |
|
Ungernia victoris |
Луковица |
U - 1 |
2.2 Синтез вторичных метаболитов
Вторичный метаболизм культивируемых клеток привлекает всё больше внимания исследователей, это обусловлено, прежде всего перспективностью промышленного использования культивируемых клеток растений для получения соединений специализированного обмена растений. Особую актуальность этот вопрос приобретает в связи с возрастающей остротой экологических проблем. В медицине 25% всех применяемых лекарств содержат соединения растительного происхождения. Если приплюсовать к этому потребности пищевой промышленности, парфюмерии, сельского хозяйства, то становится очевидной необходимость замены плантационного, а тем более дикорастущего сырья на гарантированно получаемую промышленным способом биомассу культивируемых клеток, содержащую необходимые соединения в достаточном количестве.
Как показал почти полувековой опыт исследования вторичных соединений в клеточных культурах растений (с 1940 года), для этого необходимо решение многих фундаментальных проблем биологии культивируемых клеток. Наиболее серьёзной из них является разработка стратегии контроля синтеза вторичных соединений в культивируемых клетках растений. До сих пор неясно, возможна ли разработка единой стратегии или она должна быть специфической для разных классов вторичных соединений, или же индивидуальной для каждого конкретного случая.
2.3 Влияние генетических, физических и химических факторов на рост и развитие культуры клеток и тканей, на синтез вторичных метаболитов
На культивирование клеток оказывают влияние многие факторы, такие как:
1.генетика экспланта (выбор вида растения, выбор конкретного растения донора)
2.эпигенетика экспланта (выбор органа растения)
3.генетика популяции клеток (культивирование (селекция) культуры, получение мутантов)
4.физиология популяции клеток (оптимизация условий (химических и физических факторов) роста и синтеза вторичных соединений)
А) Химические факторы.
Углеводное питание. Как показано в многочисленных работах, культуры могут расти на различных углеводах (испытано более 30 различных соединений). Как правило, лучший рост отмечается на двух сахарах – глюкозе или сахарозе. В то же время высокий уровень синтеза свойственен культурам на сахарозе. На средах с глюкозой синтез часто сильно ослаблен. Причины этого явления не ясны.
Минеральное питание. Минеральный состав сред оказывает большое влияние на синтез вторичных соединений, при этом наиболее важно содержание фосфора, калия и различных форм азота. Высокие концентрации фосфора в большинстве случаев приводят к улучшению роста культуры и ухудшению синтеза вторичных метаболитов. Их синтез начинается обычно после исчерпания фосфора из среды. Высокие концентрации фосфора в среде снижают синтез никотина в культуре клеток табака, антоцианов в культивируемых клетках моркови, фенолов в клетках чая. В то же время имеются сообщения о повышении содержания вторичных соединений при повышенных концентрациях фосфора – алкалоидов в клетках барвинка розового, антрахинонов в культуре клеток Galium.
Показано, что как для роста, так и для синтеза вторичных соединений необходимы минеральные формы азота. Органические формы – пептон, дрожжевой экстракт и другие – тормозят и рост, и синтез. Очень важно соотношение аммонийного и нитратного азота. Можно проследить определенную тенденцию – повышение доли нитратного азота способствует увеличению синтеза вторичных веществ, в частности, диосгенина в культуре клеток диоскореи.
Фитогормоны. Эти компоненты среды привлекают наибольшее внимание исследователей. Однако влияние гормонов на синтез вторичных соединений в культуре клеток неоднозначно и может изменяться в зависимости от класса вторичных соединений, физиологического состояния культуры, условий культивирования и др. Наиболее интенсивно изучались ауксины и цитокинины, так как они, как правило, являются необходимыми компонентами сред. Имеется большое число примеров, когда ауксины стимулировали синтез вторичных соединений, и примерно столько же, когда подавляли. Ауксины стимулировали синтез антоцианов в культурах моркови и тополя, антрахинонов в Cassia fistula, Cassia torra, сапогенинов в тригонелле; уменьшали и исключали синтез антрахинонов и шиконина в воробейнике, скополамина - в белене, хлорогеновой кислоты – в табаке.
Результатов по влиянию цитокининов на синтез вторичных соединений существенно меньше, что не позволяет уловить какие-либо закономерности. Результаты по другим классам других фитогормонов весьма фрагментарны и противоречивы.
РН среды. По влиянию рН среды на синтез вторичных соединений данных очень немного. Однако, по имеющимся сведениям, рН среды существенно влияет на этот процесс. Например, при изучении синтеза индола в культуре клеток ипомеи в условиях рН-стата оказалось, что при рН 6,3 синтез индола максимален и в два раза превышает уровень синтеза без рН-статирования. В рН-стате при уровне рН 4,8 синтез полностью прекращается.
Б) Физические факторы.
Аэрация. Очень важный фактор, но практических данных немного. Важно как значение расхода кислорода, так и соотношение СО2/О2. Высокое отношение О2/СО2 может ингибировать и рост и синтез вторичных продуктов.
Температура. Влияние температуры на синтез вторичных метаболитов изучалось лишь в нескольких работах. Показано, например, что оптимум синтеза никотина в клетках табака находится при 270, уклонение на 50 в любую сторону приводит к снижению синтеза более чем в 3 раза.
Свет. Этому фактору посвящено наибольшее число работ. При этом разбирается как полная индукция синтезов, так и влияние на количество соединений света разной интенсивности и качества.
Свет увеличивает содержание эпикатехинов и протоантоцианидов в культуре клеток чая, серпентина в барвинке, витамина В6 и суммы алкалоидов в дурмане и руте. Красный цвет увеличивает синтез подофиллотоксина в культивируемых клетках подофилла.
Достаточно часто освещение изменяет соотношение разных классов вторичных соединений. Например, в культуре клеток Tabernaemontana divaricata под действием света существенно изменялись соотношения аспидосперматановых, коринантиновых, плюмерановых и ибогановых алкалоидов.
В то же время часто свет выключает или снижает синтез вторичных соединений. Синтез гиосциамина и скополамина в культуре клеток дурмана происходит только в темноте, освещение снижает содержание алкалоидов в клетках культуры хинного дерева, шиконина – в культуре клеток воробейника. В последнем случае показано, что свет разрушает ФМН, необходимый для синтеза шиконина.
В) Генетические факторы.
Культуры клеток растений характеризуются большой степенью генетической гетерогенности клеток в популяции. При этом гетерогенность наблюдается на разных уровнях организации генома – на уровне точковых мутаций, хромосомных перестроек, наборов хромосом и, наконец, цитоплазматических генов. В качестве основных причин гетерогенности выдвигаются:
А) гетерогенность исходных эксплантов
Б) мутационный эффект компонентов среды культивирования
В) эффект отсутствия организменного контроля («сита мейоза» и др.)
Эта гетерогенность клеточной популяции позволяет с помощью клонирования отбирать линии клеток с сильно изменёнными свойствами, в частности с повышенным синтезом вторичных метаболитов. Таким образом были отобраны штаммы – продуценты шиконина, аймалицина, серпентина, берберина. Однако судьба отобранных клонов при длительном культивировании различна: в одних случаях синтез вторичных метаболитов в клонах нестабилен и возвращается на уровень исходной культуры, в других случаях клон достаточно стабилен. Причины такого различия в поведении не ясны. Наконец, максимальные изменения генома культивируемых клеток могут быть получены в результате индуцированного мутагенеза. В полученных мутантных штаммах возможны резкие качественные и количественные изменения синтезов вторичных соединений.
3. Культура ткани растений и синтез вторичных метаболитов
3.1 Образование полифенолов в культуре ткани чайного растения
Фенольные соединения (ФС), или полифенолы, занимают одно из центральных мест среди вторичных соединений благодаря их всеобщему распространению в растениях и разнообразным функциям (защита при патогенезах, механических повреждениях и облучении, использование в качестве запасного энергетического материала и др.). Полифенолы имеют также важное биотехнологическое значение (использование в медицине, пищевой промышленности и некоторых других областях народного хозяйства).
В качестве объекта, позволяющего исследовать процессы регуляции образования ФС, была использована культура тканей чайного растения. Известно, что это растение обладает специализированным обменом веществ, направленным на синтез соединений дифенилпропановой структуры (катехина, галлокатехина, катехингаллата.ю галлокатехингаллата).
Каллусные культуры стебля чайного растения (Camellia sinensis L., грузинская разновидность) выращивали в темноте или при непрерывном освещении (3000 лк) при 260 и относительной влажности воздуха 70% на оптимальной питательной среде, содержащей 2,4-Д (2*10-5 М) и глюкозу (2,5%). Содержание суммы растворимых ФС, флаванов и лигнина определяли спектрофотометрическими методами с использованием реактива Фолина –Дениса, ванилинового реактива и 2,6-дихлорхинонхлоримида соответственно.
Гетеротрофная (выращиваемая в темноте ) каллусная культура чайного растения, как и большинство пролиферирующих клеток, обладает более низким биосинтетическим потенциалом, чем исходная ткань интактного растения.
|
Содержание и состав фенольных соединений стебля чайного растения и полученной из него гетеротрофной каллусной культуры |
|||||||
|
Содержание, мг/г сухой массы |
Качественный состав |
||||||
|
ФС |
ФЛ |
Л |
ПК |
ГК |
ПА |
ФВ |
Л |
|
Молодой стебель |
|||||||
|
80,0 |
66,3 |
4,0 |
++ |
++ |
+ |
+ |
+ |
|
Каллусная культура |
|||||||
|
16,2 |
10,0 |
5,0 |
+ |
- |
++ |
- |
+ |
ФС – фенольные соединения; ФЛ – флаваны; ПА – проантоцианидины; ГК – галлокатехины и галловые эфиры катехинов; ФВ – флавонолы; Л – лигнин.
Как видно из представленных данных, в каллусной культуре, происходит как уменьшение общего содержания ФС, так и обеднение их качественного спектра. Однако, способность к синтезу характерных для чайного растения флаванов (представленных простейшими катехинами и их биогенетическими аналогами проанотоцианидинами), а также фенольного полимера лигнина сохраняется. При этом содержание проантоцианидинов и лигнина оказывается даже выше, чем в исходной ткани. Как свидетельствуют данные электронно-микроскопических исследований, это обусловлено, по-видимому, активацией эндомембранной системы клеток (эндоплазматического ретикулума и аппарата Гольджи), являющиеся местом синтеза фенилпропаноидных предшественников. Из всего этого следует, что по составу фенольного комплекса и активности фенольного метаболизма гетеротрофная каллусная культура приближается к тканям корня интактного растения.
К числу факторов, способных оказывать влияние на биосинтетический потенциал клетки, относятся гормоны и гормоноподобные соединения, а также свет.
В гетеротрофной каллусной культуре чайного растения НУК (2*10-5 М), введённая в питательную среду взамен 2,4-Д, значительно стимулирует образование растворимых ФС (примерно в 10 раз) и в меньшей степени лигнина (в 3 раза). Это согласуется с литературными данными о том, что НУК может быть использована в качестве ауксина для «продукционных» сред, т. е. сред, способствующих накоплению вторичных соединений.
В отличии от НУК, 1 мг/л кинетина (5*10-6 М) способствует главным образом лигнификации тканей ( в 3 раза по сравнению с контролем), лишь незначительно влияя на образование флаванов.
Таким образом, введением в питательную среду гормоноподобных соединений можно добиться направленной регуляции синтеза определённых типов ФС в культивируемых in vitro клетках и тканях. При этом ауксины (НУК в большей степени, чем 2,4-Д) способствуют синтезу растворимых ФС, в том числе и характерных для чайного растения флаванов, тогда как цитокинины преимущественно воздействуют на образование лигнина. При этом во всех случаях происходит лишь количественные изменения в синтезе ФС, что обусловлено, по-видимому, активацией ферментов лишь тех звеньев фенольного метаболизма, которые являются общими для изученных полифенолов.
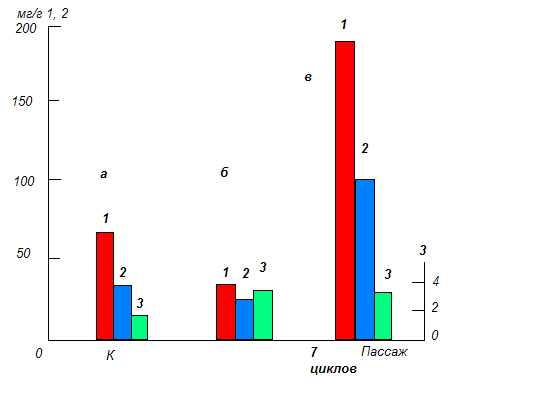
Влияние света. Так, перенесение гетеротрофных каллусных культур чайного растения в условия непрерывного освещения во время первых двух субкультивирований приводит главным образом к увеличению образования лигнина ( в 1,5 раза), тогда как образование растворимых ФС несколько снижается.
Влияние длительного освещения на образование суммы растворимых ФС(1), флаванов(2) и лигнина(3) в каллусной культуре чайного растения (в мг/г сухой массы). К – контроль; а – темнота; б, в – свет.
При длительном же пассировании культур в условиях непрерывного освещения (7 культуральных циклов) содержание суммы растворимых ФС, а также флаванов значительно увеличивается. Как показали электронно-микроскопические исследования, это связано с формированием в ткани хлоропластов. Следует также отметить, что такие фотомиксотрофные каллусные культуры помимо флаванов синтезируют ещё один класс характерных для чайного растения ФС, а именно флавонолы. Последние представлены кемпферолом и кверцетином, а также несколькими их гликозидами. Таким образом, формирование в каллусной культуре хлоропластов, являющихся одним из центров синтеза ФС в клетках растений, оказывает значительное влияние на их биосинтетический потенциал и, главное, способствует расширению спектра синтезируемых ФС.
Таким образом, полученные данные свидетельствуют о том, что в случае гетеротрофных (не содержащих хлоропласты) каллусных культур усиление образования ФС при действии фитогормонов (ауксинов и цитокининов) или света ( на протяжении первых субкультивирований) происходит лишь за счёт активации ряда ферментов фенольного метаболизма (фенилаланинаммиаклиазы, 4-гидроксилазы коричной кислоты, оксициннамоил-КоА-лигазы и др.). В случае же частично фототрофных (содержащих хлоропласты) культур увеличение образования ФС происходит за счёт двух слагаемых: активации внехлоропластовых ферментных систем (как в первом случае) и благодаря функционированию в хлоропластах специализированного центра биосинтеза ФС.
3.2 Образование -карболиновых алкалоидов в культуре ткани гармалы обыкновенной
Гармала обыкновенная (Peganum harmala) – лекарственное растение, относящееся к семейству парнолистниковых (Zygophyllaceae) и широко применяемое в народной и официальной медицине. Терапевтический эффект экстрактов гармалы, обладающих значительным влиянием на сердечно-сосудистую деятельность и на центральную нервную систему, обусловлен содержанием в ней алкалоидов, которые оказывают ингибирующее действие на такие ферменты, как моноаминооксидаза и ацетилхолинэстераза. Помимо этого, алкалоиды гармалы проявляют антибактериальную активность, которая увеличивается при использовании УФ-света, что свидетельствует об их фототоксичности.
В целом растении обнаружены два типа алкалоидов – хиназолиновые, производные антраниловой кислоты, и индольные -карболинового типа, производные триптофана. При этом надземная часть растения содержит оба типа алкалоидов, в то время как -карболиновые алкалоиды характерны в основном для корней растения. Наиболее богаты алкалоидами семена гармалы, в которых содержание этих соединений может достигать 5 – 6 %. Корни растения в ранний период вегетации содержат около 1% алкалоидов, в то время как надземная часть – лишь 0,03%. Характер распределения алкалоидов и изменение их содержания в целом растении в течение его вегетации послужили основанием для высказывания предположения о том, что место синтеза -карболиновых алкалоидов являются корни гармалы.
Для экспериментальной проверки правильности этой гипотезы были введены в культуру каллусные ткани от гипокотиля и корня проростка гармалы, полученного из семян египетского происхождения. Ткани выращивали на агаризованной среде Мурасиге и Скуга с добавлением 2,4-Д (1 мг/л) и кинетина (1 мг/л). несмотря на то, что каллусные ткани были получены от частей ювенильного растения, они имели некоторые морфологические различия, которые сохранялись в течение четырёхлетнего культивирования: гипокотильная ткань была более плотной и отличалась от корневой ткани своей кремовой окраской, причём в ряде случаев на отдельных участках ткани была заметна яркая красная пигментация. Общим свойством тканей была их относительная гомогенность и отсутствие визуальных признаков дифференциации.
В каллусных тканях гармалы первых пассажей было обнаружено присутствие веществ, имеющих голубую и ярко-жёлтую флуоресценции. При экстрагировании лиофилизированных тканей метанолом и последующем хроматографировании концентрированных экстрактов в тонком слое силикагеля в системе хлороформ : метанол : аммиак (10 : 4 : 1) было показано присутствие в тканях четырёх -карболиновых алкалоидов – гармина, гармалина, гармола и гармалола, типичных для корней целого растения. Идентификация алкалоидов в каллусных тканях проводилась путём хроматографирования экстрактов с аутентичными образцами алкалоидов в различных системах растворителей и сопоставления спектров поглощения и спектров флуоресценции обнаруженных веществ с соответствующими спектрами чистых алкалоидов.
По своему качественному составу гипокотильная и корневая ткани не различались. Доминирующими алкалоидами в обеих тканях были гармин и гармалол, количественное содержание которых определяли по поглощению в УФ-свете (гармин – при 245 нм, гармалол – при 390 нм).
В условиях оптимального, но недифференцированного роста содержание гармина в каллусных тканях было почти в 500 раз, а гармалола – 20 раз ниже, чем в исходном растении. Концентрация гармина и гармалола была выше в гипокотильной ткани, однако с пассированием эта разница практически нивелировалась, а общее содержание алкалоидов резко снижалось.
|
Изменение содержания индольных алкалоидов (мкг/г сухой ткани) в каллусных тканях гармалы при культивировании |
||||
|
Номер пассажа |
Корневая ткань |
Гипокотильная ткань |
||
|
Гармин |
Гармалол |
Гармин |
Гармалол |
|
|
5 |
62,2 |
89,2 |
174,1 |
129,7 |
|
7 |
45,2 |
11,4 |
50,2 |
40,5 |
С целью регуляции степени дифференциации каллусных тканей гармалы и соответственно образования в них алкалоидов в питательную среду вместо 2,4-Д вводили индолилмасляную кислоту (ИМК) (2 мг/л), а также в ряде случаев произвели замену кинетина на бензиламинопурин (1 мг/л). изменение гормонального состава питательной среды вызывает снижение ростовой активности тканей, увеличение их плотности и проявление в них признаков корневой дифференциации. Длительное субкультивирование тканей на среде, содержащей ИМК, приводило к появлению некрозов у корневой ткани и к прекращению её роста. Гипокотильная ткань сохраняла ростовую активность, однако она была значительно ниже, чем у ткани, растущей на 2,4-Д.
Химический состав каллусных тканей гармалы при перенесении их на среду, содержащую ИМК, показал, что концентрация -карболиновых алкалоидов в них резко увеличилась. Однако, снижение ростовой активности тканей при проявлении корневой дифференциации не компенсировалась увеличением концентрации алкалоидов, и общая продуктивность тканей при этом практически не увеличилась.
|
Влияние сочетания различных гормонов на содержание индольных алкалоидов в каллусных тканях гармалы (мкг/г сухой ткани) |
|||||
|
Сочетание экзогенных гормонов |
Номер пассажа |
Корневая ткань |
Гипокотильная ткань |
||
|
Гармин |
Гармалол |
Гармин |
Гармалол |
||
|
2,4-Д+кинетин |
5 |
62,2 |
89,2 |
174,1 |
129,7 |
|
ИМК+кинетин |
11 |
308,1 |
621,6 |
212,1 |
240,0 |
|
ИМК+БАП |
11 |
161,1 |
200,0 |
262,2 |
439,2 |
Ненадёжность изменения состава экзогенных гормонов и нестабильность роста каллусных тканей гармалы при таком способе регуляции их дифференциации и биосинтеза индольных алкалоидов заставили обратиться к генетической трансформации клеток гармалы с помощью Ri-плазмиды Agrobacterium rhizogenes. Для этого использовали дикий штамм А-4, суспензией клеток которого было проведено инфицирование пораненных участков гипокотиля стерильного проростка гармалы.
Через 3 – 4 недели после заражения на месте поранения и инфицирования можно было увидеть образование хорошо растущих адвентивных корней. При вычленении этого участка гипокотиля и его переносе на питательную среду, содержащую антибиотики (клафоран, 500 мг/л), после двух субкультивирований удавалось добиться элиминирования бактерий. Последующее отделение корней и перенесение их на безгормональную питательную среду привело к получению интенсивно растущей ризогенной культуры, которую принято называть культурой «бородатых корней» (“hairy root culture”). О том, что это генетически изменённая культура, свидетельствовал характер образования и роста корней, независимость их роста от экзогенных гормонов и содержание в культуре маннопина.
Ризогенная культура гармалы хорошо росла в жидкой питательной среде при условии её небольшого объёма и постоянного перемешивания. Полное погружение корней в питательную среду и статика не обеспечивали достаточной для роста аэрации культуры. Наилучший рост ризогенной культуры наблюдался на подложках (вискоза и бумажный фильтр), смоченных избытком питательной среды. При таком способе выращивания трёхнедельная культура гармалы достигала 20-кратного увеличения массы.
Генетически трансформированная ризогенная культура гармалы наряду с проявлением корневой дифференциации восстанавливала и способность к биогенезу индольных алкалоидов. Просмотр культур в УФ-свете показывал интенсивную флуоресценцию корней, типичную для гармина и гармалола. Хроматоспектрометрический анализ метанольных экстрактов из ризогенной культуры показал, что она по количественному и качественному составу алкалоидов мало отличается от корней ювенильных растений гармалы. Если при максимальном содержании -карболиновых алкалоидов в неорганизованно растущих тканях концентрация гармина в них была более чем в 100 раз, а гармалола – в 5 раз ниже концентрации этих алкалоидов в корнях проростка, то в трёхнедельной ризогенной культуре 12-го пассажа концентрация гармина составляла 1/3 от концентрации алкалоида в корнях проростка, а концентрация гармалола превышала концентрацию алкалоида в корнях. При этом восстанавливалось и типичное для корней целого растения преобладание гармина над гармалолом.
|
Содержание алкалоидов в ризогенной культуре гармалы (мг/г сухой массы корней) |
||
|
Пассаж культуры |
Гармин |
Гармалол |
|
12-й пассаж (3-недельная культура) |
12,85 |
2,46 |
|
12-й пассаж (5-недельная культура) |
19,90 |
2,01 |
|
Корни проростков |
50,66 |
2,16 |
|
Стебель проростков |
10,85 |
1,84 |
Если принять во внимание интенсивность роста ризогенной культуры (20-кратное увеличение массы) и непрерывность её выращивания, то можно увидеть, что продуктивность такой культуры значительно превышает продуктивность ювенильных растений гармалы.
Таким образом, генетически трансформированная культура гармалы, наделённая способностью к синтезу гормонов, обеспечивающих появление корневой дифференциации, характеризуется высоким уровнем содержания -карболиновых алкалоидов, который вполне сравним с уровнем биосинтеза этих алкалоидов в органах целого растения. Чётко установленная зависимость между корневой дифференциацией и уровнем синтеза -карболиновых алкалоидов подтверждает правильность гипотезы о локализации образования этих индольных алкалоидов в корнях гармалы. В этом отношении ризогенная культура гармалы наряду с аналогичными культурами белладонны, дурмана, белены и других паслёновых служит примером корнеспецифичности биосинтеза определённых групп алкалоидов.
Результаты, полученные с трансформированной культурой гармалы, показывают возможность и эффективность использования приёмов генетической инженерии не только на растениях семейства паслёновых, но и на растениях других семейств, в которых образование алкалоидов или других вторичных веществ локализовано в корневой системе. Индукция биосинтеза вторичных веществ в таких генетически изменённых культурах может служить реальной основой для получения искусственно выращиваемых растительных систем, продуцирующих те же ценные биологические вещества и на том же количественном уровне, что и целое растение.
3.3 Накопление алкалоидов в культуре ткани раувольфии змеиной
Культура ткани раувольфии змеиной (Rauwolfia serpentina) является перспективным источником аймалина – алкалоида группы индолина, обладающего противоаритмическим действием. Аймалин содержится в смеси алкалоидов биомассы в количестве 30-40%. 40-50% составляет вомиленин – индолениновый алкалоид, который рассматривается как биогенетический предшественник аймалина. Превращение вомиленина в аймалин под действием ферментов, выделенных из клеток раувольфии, в настоящее время осуществлено in vitro и включает в себя гидрирование вомиленина и его метилирование. Источником пиридиннуклеотидов является витамин РР (никотиновая кислота и никотинамид). Перенос метильных групп осуществляется с помощью серосодержащих аминокислот – метионина и ацетилметионина.
Целью настоящей работы было выяснение влияния витамина РР, серосодержащих аминокислот, вомиленина (как предшественника аймалина) и сульфат-иона на накопление алкалоидов, в частности, аймалина в стеблевой культуре ткани раувольфии (клеточная линия А). В качестве контроля использовали регламентную питательную среду (Р). В опытных вариантах в неё вносили 2 или 5 мг/л никотиновой кислоты (среды Н2 и Н5), 10 мг/л метионина или ацетилметионина (среды М и АМ), 1 мг/л вомиленина (среда В). Вомиленин добавляли после автоклавирования среды в асептических условиях, остальные добавки вносили до автоклавирования. Среды S1 и S2 были обеднены неорганической серой: в S1 не вносили сульфат аммония; в S2, кроме того, в 5 раз уменьшили содержание сульфата магния. Опыты проводили на трёх пассажах в десятикратной повторности. После культивирования в течение 70-75 суток при 2620 биомассу высушивали. В ней титриметрически определяли сумму алкалоидов, фотоколориметрическим методом – аймалин, хроматоспектрофотометрическом – вомиленин.
Результаты опытов приведены в таблице. Они показывают, что добавление к среде никотиновой кислоты не влияло на содержание алкалоидов в биомассе. Снижение содержания в среде сульфат-иона привело к уменьшению количества суммы алкалоидов, аймалина – в 1,5 раза, и к возрастанию (приблизительно в 2 раза) содержания в биомассе вомиленина. При введении в среду серосодержащих аминокислот в 1,2 раза увеличилось общее содержание алкалоидов в биомассе, а содержание аймалина возросло в 2 раза. Добавление вомиленина вызвало накопление в биомассе аймалина при сохранении контрольного уровня содержания суммы алкалоидов. Это вполне согласуется с литературными данными о превращении вомиленина в аймалин.
|
Накопление алкалоидов в биомассе в зависимости от состава среды |
||||||||
|
Количество биомассы или алкалоидов |
Питательная среда |
|||||||
|
Р |
Н2 |
Н5 |
S1 |
S2 |
М |
АМ |
В |
|
|
Биомасса, г/1л среды |
29,02,5 |
27,53,3 |
27,84,6 |
24,63,0 |
27,33,8 |
24,02,0 |
25,62,7 |
25,32,7 |
|
Сумма алкалоидов |
||||||||
|
Мг/1л среды |
47231 |
49049 |
50636 |
31032 |
31424 |
46345 |
44038 |
41613 |
|
% к воздушно-сухой массе |
1,630,11 |
1,780,18 |
1,820,13 |
1,260,13 |
1,150,09 |
1,930,19 |
1,720,15 |
1,640,05 |
|
Аймалин |
||||||||
|
Мг/1л среды |
1307 |
11522 |
11119 |
597 |
8210 |
27623 |
30732 |
20745 |
|
% к воздушно-сухой массе |
0,450,05 |
0,420,08 |
0,400,07 |
0,240,05 |
0,300,03 |
1,150,12 |
1,200,07 |
0,820,18 |
|
% к сумме алкалоидов |
28,8 |
23,6 |
22,1 |
19,0 |
26,1 |
59,2 |
69,8 |
50,0 |
|
Вомиленин |
||||||||
|
Мг/1л среды |
11615 |
821 |
864 |
15017 |
14219 |
11512 |
1283 |
9610 |
|
% к воздушно-сухой массе |
0,400,05 |
0,300,02 |
0,310,03 |
0,610,07 |
0,520,07 |
0,480,05 |
0,500,01 |
0,380,04 |
|
% к сумме алкалоидов |
24,5 |
17,0 |
17,0 |
48,4 |
45,2 |
24,8 |
29,0 |
23,1 |
Результаты исследования свидетельствуют о том, что накопление алкалоидов в биомассе связано с содержанием в среде как неорганической серы, так и серосодержащих аминокислот. Для получения обогащенной аймалином биомассы целесообразно добавлять к питательной среде метионин и ацетилметионин.
3.4 Алкалоиды каллусных тканей мака прицветникового
Мак прицветниковый (Papaver bracteatum) может быть использован c целью получения тебаина – морфинового алкалоида, являющегося предшественником кодеина и морфина. Тебаин составляет до 98% от суммы алкалоидов P.bracteatum и в лабораторных условиях легко может быть переведён в кодеин наиболее широко распространённое в мире противокашлевое средство.
Литературные данные свидетельствуют о том, что в культуре клеток растений рода Papaver доминируют алкалоиды протопионового, бензафенантридинового и тетрагидропротоберберинового типов, берущих начало от (+)-ретикулина и отсутствуют, либо присутствуют в следовых количествах алкалоиды морфинановой группы, характерные для растений это рода. При повышении уровня дифференцировки может проявляться тенденция к восстановлению спектра алкалоидов, характерного для целого растения.
Целью настоящей работы являлось получение каллусных тканей P.bracteatum, регенерантов из них и сравнение спектров алкалоидов в культуре тканей и в онтогенезе целых растений.
Штамм Р11 был способен к регенерации при культивировании по следующей схеме: каллусная ткань, выращиваемая в темноте, пересаживалась на среду, отличающуюся от исходной отсутствием гормонов, и выдерживалась в течение 30 дней в темноте. При этом в примыкающем к среде слое каллуса образовывались очаги дифференциации, которые затем пересаживали на свежую питательную среду того же состава и выставляли на свет. Через 2-3 недели внутри и на поверхности каллуса наблюдалось появление зачатков зелёных листочков, иногда образовывались корни. По мере формирования листочков регенеранты высаживали на среду Стрита для активации образования корней. При достижении корнями длины 5-10 мм регенеранты высаживали в вазоны со стерильной почвой и переносили в теплицу.
Присутствие в растительных образцах сангвинарина или тебаина определялось с помощью хроматографии хлороформного экстракта лиофилизированной ткани в тонком слое силикагеля, содержащего и не содержащего люминесцентную добавку, для раздельного определение флуоресцирующих и поглощающих зон.
Количественное определение сангвинарина в культуре тканей рассчитывали по экстинкции при 340 нм вытяжки 10%-ной H3SO4 из хлороформного экстракта лиофилизированной ткани. О количестве алкалоида, переходящего в сангвинарин, судили по разности в содержании сангвинарина в подкисленном муравьиной кислотой хлороформном экстракте до и после облучения его светом лампы ДРШ-250 в течение 1 минуты.
От различных частей двулетних растений мака прицветникового кавказской популяции и популяции Arya 11 были получены каллусные ткани. При качественном изучении спектра алкалоидов в каллусных культурах тебаин обнаружен не был, в то время как в тканях всех штаммов присутствовал сангвинарин, не характерный для целого растения. Содержание сангвинарина в тканях колебалось от 0,5 до 1,7%, а в штамме, полученном из цветоноса растения популяции Arya 11, оно составляло 2,70,4%. Наряду с сангвинарином в тканях был обнаружен алкалоид, который на свету переходил в сангвинарин. Содержание его в каллусных тканях по отношению к общему содержанию сангвинарина составляло от 11 до 40%.
Изучение спектров целых растений показало, что они способны к синтезу не только тебаина, но и сангвинарина. Тебаин присутствовал во всех частях растений на протяжении всего времени наблюдения за ними, в то время как сангвинарин был обнаружен в листьях растений только первого года жизни, при чём содержание его с возрастом снижалось. В растениях, выращенных в естественных условиях, в пределах своего ареала, сангвинарин обнаружен не был, и биосинтез его является, по-видимому, явлением, не характерным для сформировавшихся растений.
Из этого факта, что растения мака прицветникового способны в определённых условиях продуцировать сангвинарин, следует, что проявление способности к его биосинтезу в культуре тканей данного растения не является случайным. Исследование спектра алкалоидов регенерантов, полученных из каллусной культуры штамма Р11, показало, что они были способны к биосинтезу сангвинарина, а тебаин был обнаружен в листьях растений-регенерантов лишь спустя 5 месяцев после выращивания их теплице.
Таким образом, способность к биосинтезу алкалоидов тебаина и сангвинарина позволяет выделить две противоположные ситуации: культура тканей способна к биосинтезу только сангвинарина, а выращиваемые в природных условиях растения – к биосинтезу одного тебаина. Кроме того, имеются и промежуточные переходные положения, когда биосинтез обоих алкалоидов одновременно идёт в растениях-регенерантах и в ювенильных растениях.
По-видимому, биосинтез сангвинарина связан с ювенильной стадией развития растения и его появление в системе с низким уровнем дифференцировки является, следовательно, закономерным для растений рода Papaver.
3.5 Образование вторичных метаболитов в культуре тканей растений семейства Rutaceae
Многие растения семейства рутовых известны как продуценты физиологически активных соединений. Интерес к этому семейству особенно возрос после обнаружения у некоторых метаболитов Rutaceae противоопухолевой активности. Лечебные свойства руты обыкновенной и близкого, но менее известного вида Boenninghausenia albiflora обусловлены наличием в них кумаринов, фурохиналиновых и акридоновых алкалоидов. Спектры кумаринов и алкалоидов обоих видов в значительной мере подобны, однако в B.albiflora обнаружен неизвестный до сих пор для Ruta акридоновый алкалоид норакроницин, который легко преобразуется in vitro в акроницин, обладающий широким спектром противоопухолевой активности.
Из стерильных проростков семян B.albiflora были получены каллусные культуры, которые проявили различные морфологические признаки при выращивании на среде Мурасиге и Скуга. При длительных пересевах были отобраны штаммы: В1 –гомогенная ткань, В2 –ткань с ризогенезом и В3 –ткань с элементами стеблевого органогенеза.
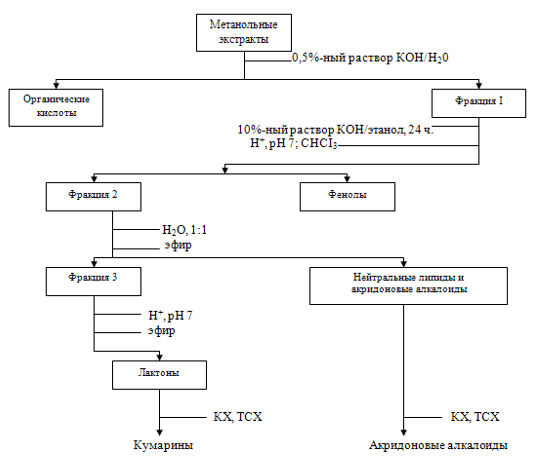
Для изучения состава вторичных метаболитов, синтезируемых в клетках B.albiflora in vitro, лиофилизированные ткани В1, В2 и В3 экстрагировали метанолом.
Предварительная тонкослойная хроматография (ТСХ) метанольных экстрактов показало наличие множества зон с флуоресценцией, типичной для кумаринов. Из суммы метанол-растворимых веществ были выделены фракции органических кислот, фенольных соединений и лактонов, мешающих обнаружению пренилированных акридонов (рутокредон, норакроницин).
Фракционирование метанольных экстрактов тканей В1, В2, В3
С помощью препаративной ТСХ из фракций выделяли индивидуальные соединения и затем идентифицировали их сравнением с чистыми веществами по хроматографическому поведению и УФ-спектром.
Анализ фракций лактонов показал наличие во всех трех штаммах (в качестве преобладающих компонентов) кумаринов: рутомарина, рутакультина, бергаптена, ксантотоксина, хелиеттина, дафноретина и умбеллиферона. Следует отметить, что кумариновый состав штаммов В1, В2, В3 в значительно большей мере соответствует составу В.japonica (наличие бергаптена, ксантотоксина, рутомарина, умбеллиферона и дафноретина), чем состава В.albiflora, в котором ранее были обнаружены лишь бергаптен, дафноретин, и следы рутомарина. Рутакультин и хелиеттин (дезацитилрутамарин) обнаружены в роде Boenninghusenia впервые.
Из остатков метанольных сумм, обогащенных липидными компонентами и имеющими характерный желтый цвет, были выделены и идентифицированы алкалоиды рутакридон и его водорастворимое производное – глюкозит гравакридондиола. Норакроницин ни в одном из штаммов не был обнаружен.
Штаммы В1 и В3 (гомогенные не дифференцированная ткань и ткань с элементами стеблевого органогенеза) по составу кумаринов и пренилированных акридонов практически не различались, тогда как ткань с выраженным ризогенезом – штамм B2 – характеризовалась значительно большим рутакультина и рутакридона. Содержание рутакридона и глюкозида гравакридондиола, определено методами ТСХ и УФ-спектрометрии, составляло для штаммов В1 и В2 – следы и 0,01; 0,01 и 0,03% соответственно.
Известно, что в культурах клеток Rutaceae в большинстве случаев не теряется способность к синтезу вторичных метаболитов, но их концентрация бывает значительно ниже, чем в органах интактных растений. В некоторых случаях в культуре обнаруживается также изменение в спектре метаболитов, вплоть до появления веществ, не типичных для целого растения. Изменения в спектре вторичных метаболитов в культуре ткани и снижение их концентрации объясняется физиологическими особенностями растительных клеток, не достигающих в культуре in vitro уровня дифференциации, соответствующего уровню специализированных клеток целого растения.
Отсутствие в исследованных штаммах ткани В.albiflora норакроницина, возможно, объясняется именно низким уровнем организации культивируемых клеток. Взаимосвязь физиологической и биохимической дифференциации в культуре клеток В.albiflora доказывается так же тем, что при исследовании ее с помощью люминесцентного микроскопа ярко-оранжевая флуоресценция, характерная для акридоновых алкалоидов, была обнаружена только в резогенном штамме В2. Причем флуоресценция была локализована в корневых трихомах и в некоторых поверхностных клетках корней. Количественное определение акридоновых алкалоидов В1 и В2 также подтверждает это положение.
Таким образом, в штаммах тканях В.albiflora, не содержащих типичного для целого растения алкалоида норакроницина, обнаружены наряду с рутакридоном и глюкозидом гравакридондиола, кумарины и фурокумарины, характерные для растений рода Boenninghusenia, а также новые для рода кумарины – рутакультин и хелиеттин. Количество акридоновых алкалоидов в штаммах зависит от уровня дифференциации ткани.
Заключение
В заключении хочется отметить, что эта тема является очень перспективной для фармацевтической промышленности, так как продуктивность культуры тканей можно регулировать, а значит количество необходимого соединения будет гарантировано. Кроме того культуры защищены от внешних неблагоприятных факторов, способных снизить качество сырья. Процесс сбора и обработки такого сырья значительно упрощается за счёт культивирования только нужной ткани растения в стерильных условиях. Проблемой является трудоёмкий процесс отбора необходимой ткани и разработки среды для получения оптимального результата. Работы которые представлены в данной курсовой – это лишь малая часть, которая не отображает всего масштаба этой темы.
Литература
Бутенко Р.Г. Культура изолированных тканей и физиология морфогенеза растений. М.: Наука, 1964. 272 с.
Бутенко Р.Г. Культура клеток растений и биотехнология. М.: Наука, 1986. 286 с.
Бутенко Р.Г. Биология культивируемых клеток и биотехнология растений. М.: Наука, 1991. 280 с.
Бутенко Р.Г. Клеточные технологии для получения экономически важных веществ растительного происхождения. М.: 1986. с.3-20.
Воллосович А.Г.//Культура изолированных тканей и клеток растений. М.: Наука, 1970. с.234-235.
Действие кинетина на дифференциацию и образование фенольных соединений в каллусной культуре чайного растения / М.Н. Запрометов и др. // Физиол. растений, 1986. T. 33. № 2. С. 356-364.
Запрометов М.Н. Вторичный метаболизм и его регуляция в культурах клеток и тканей растений // Культура клеток растений. М.: Наука, 1981. С. 37-50.
Кунах В.А. Изменчивость растительного генома в процессе дедифференцировки и каллусообразования in vitro // Физиол. растений, 1999. Т. 46. № 6. С. 919-929.
Лекарственное сырьё растительного и животного происхождения. Фармакогнозия: учебное пособие/ под ред. Г.П.Яковлева. СПб.: СпецЛит, 2006. 845 с.
1