Старая гипотеза «перевернутости» хордовых подтверждается
Старая гипотеза «перевернутости» хордовых подтверждается
Д.А. Воронов
В 1822 г. французский зоолог Этьен Жоффруа Сент-Илер (1772 - 1844) опубликовал статью, в которой утверждал, что позвоночные и членистоногие животные, при всех различиях между ними, устроены по одному плану [1]. Однако у позвоночных центральная нервная система расположена на спинной стороне, сердце - на брюшной, а у членистоногих наоборот. Жоффруа Сент-Илер легко избавился от противоречия, нарисовав перевернутого кверху брюхом омара. Так возникла гипотеза, согласно которой позвоночные животные - это перевернутые в процессе эволюции беспозвоночные.
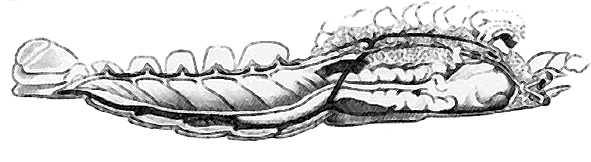
Перевернутый омар (Жоффруа Сент-Илер Э., 1822).
К столь странной на первый взгляд гипотезе Жоффруа Сент-Илер пришел потому, что уже с начала научной работы был уверен в единстве строения животных. Вероятно, на его мировоззрении сказалось то, что до занятий зоологией он изучал физику - науку, уже тогда успешно объяснявшую разнообразные природные явления с помощью небольшого числа единых принципов. В 1796 г. в одной из своих первых работ Жоффруа Сент-Илер писал: “...природа замкнулась в определенных рамках и создала все живые существа по единому плану, одинаковому в принципе, но который она варьировала на тысячу ладов во всех его деталях” [2]. Любопытно, что столь общее утверждение было высказано в статье, посвященной частному вопросу систематики полуобезьян.
В более поздних работах (1806 и 1807) Жоффруа Сент-Илер доказал общность строения рыб, рептилий, птиц и млекопитающих, позволившее объединить их в один тип позвоночных животных. Изучая главным образом скелет, он сумел сопоставить даже столь непохожих друг на друга животных, как рыбы и млекопитающие. Именно он обнаружил, что три слуховые косточки в черепе млекопитающих - это видоизмененные жаберные дуги рыб.
Такое соответствие называется гомологией. Какие-либо структуры гомологичны, если они произошли от одной и той же структуры общего предка сравниваемых животных, даже если в настоящее время они выполняют разные функции. Например, рука человека гомологична крылу птицы, так как у них общее происхождение, и между их частями можно установить соответствие. Структуры же разного происхождения, выполняющие одинаковые функции, называют аналогичными. Пример аналогичных структур - крыло птицы и крыло мухи.
Если при исследовании позвоночных животных идея единства строения оказалась чрезвычайно плодотворной, то найти и доказать общность в организации позвоночных и беспозвоночных было довольно трудно. Выдвинув гипотезу “перевернутости” позвоночных, уже саму по себе спорную, Жоффруа Сент-Илер стал искать гомологичные органы у беспозвоночных. Однако на этом пути его ждала неудача, так как он нередко отождествлял явно несопоставимые структуры. Например, Жоффруа Сент-Илер полагал, что наружный скелет членистоногих гомологичен позвоночнику, с той лишь разницей, что членистоногие живут внутри этого “позвоночника”, как улитка - внутри раковины. Позднее было показано, что скелеты членистоногих и позвоночных не гомологичны, так как они развиваются из разных зародышевых листков: у членистоногих - от наружного (эктодермы), а у позвоночных - от среднего (мезодермы).
В 1830 г. знаменитый французский зоолог и палеонтолог Жорж Кювье (1769 - 1832) в известной публичной дискуссии подверг взгляды Жоффруа Сент-Илера сокрушительной критике [3]. Сам Кювье полагал, что, существует четыре типа животных - позвоночные, моллюски, членистые и лучистые, каждый из которых характеризуется особым планом строения, не позволяющим их сравнивать. Несмотря на это, гипотеза “перевернутости” позвоночных не была забыта, и у нее время от времени появлялись сторонники, среди которых особенно известен немецкий зоолог Антон Дорн (1840 - 1909), основатель Неаполитанской морской зоологической станции. Дорн связывал происхождение позвоночных с перевернутыми кольчатыми червями [4]. Однако в целом гипотеза была непопулярной и, как правило, даже не упоминалась в учебниках.
Примеры “перевернутых” животных
Распространенная причина неприятия гипотезы “перевернутости” позвоночных - ее кажущаяся бессмысленность “с точки зрения животного”. Так, авторы известного учебника по анатомии позвоночных А.Ромер и Т.Парсонс пишут, что с гипотезой “перевернутости” трудно согласиться, “так как даже червяк различает верх и низ” [5]. Однако авторы, похоже, “забыли” что перевернутость встречается часто, причем среди самых разных животных.
Из беспозвоночных всю жизнь проводят кверху брюшной стороной обитающие в толще воды жаброногие раки (Anostraca). Для плавания поворачиваются на спину клопы семейства гладышей (Notonectidae), щитни (Notostraca, подотряд листоногих раков) и мечехвосты (Xiphosura, класс хелицеровых). Много перевернутых животных в сообществе морских и пресноводных организмов, обитающих снизу поверхностной пленки воды (гипонейстон). К ним относятся разнообразные ресничные черви (Turbellaria), брюхоногие моллюски (Gastropoda) и др. Ветвистоусые ракообразные из рода Scapholeberis прикрепляются к поверхностной пленке за счет несмачиваемых щетинок, расположенных на брюшной стороне раковины, а брюхоногие моллюски из рода Janthina всю жизнь проводят у поверхности открытого океана, прикрепившись снизу к поплавку, который они сами строят из воздушных пузырьков.
Кверху брюхом зарываются в грунт ланцетники (Branchiostoma, или Amphioxus). Эти животные принадлежат к подтипу бесчерепных современного типа хордовых, к которому еще относятся подтипы оболочников и позвоночных. Поэтому далее мы будем говорить не о типе позвоночных, как было принято во времена Жоффруа Сент-Илер, а о типе хордовых, как принято теперь.
Любители плавать на спине есть и среди рыб, например представители семейства перистоусых сомов (Mochocidae). Полагают, что им пришлось перевернуться, чтобы питаться у поверхности (остальные сомы кормятся на дне водоемов), так как рот у сомов расположен с брюшной стороны. Например, обитающий в бассейне реки Конго чернобрюхий перистоусый сомик (Synodontis nigriventris) хватает падающих в воду насекомых или объедает обрастания с плавающих на поверхности листьев. У этого сомика, чтобы быть незаметным с воздуха и из глубины, даже окраска “перевернута”: в отличие от других рыб, его спина светлее, чем брюхо.
“Перевернутые” животные есть даже среди млекопитающих. Ленивцы (Bradypodidae) почти всю жизнь висят на ветвях деревьев спиной вниз. Сообразно положению тела, шерсть у ленивцев растет не от спины к брюху, как у других млекопитающих, а наоборот, так, чтобы стекала дождевая вода.
Если гипотеза “перевернутости” позвоночных верна, то ленивцев и перистоусых сомиков следует считать вернувшимися в нормальное для далеких предков положение! Добавим к этому, что бывают и другие нестандартные положения животных - достаточно вспомнить лежащую на боку камбалу или вертикальную позу человека.
Данные сравнительной анатомии и эмбриологии
Непопулярность гипотезы “перевернутости” хордовых может объясняться еще и недостатком сравнительно-анатомических доводов в ее пользу, что проявилось еще во время дискуссии Жоффруа Сент-Илера и Кювье.
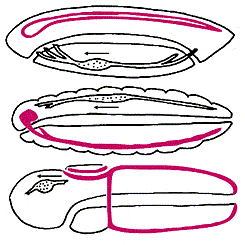
Сопоставление схем организации животных разных типов (сверху вниз):
хордовых (ланцетника), членистоногих и полухордовых.
Передний конец тела всюду обращен влево, спинная сторона - кверху.
Центральная нервная система выделена цветом,
направление кровотока показано стрелкой рядом с сердцем.
Жоффруа Сент-Илер отмечал обратное, по сравнению с беспозвоночными, расположение нервной системы и сердца у хордовых. Но расположение нервной системы - критерий недостаточно убедительный: у беспозвоночных может быть несколько продольных нервных стволов, причем не только на брюшной стороне, но также по бокам и вдоль спины. Например, у представителей класса кишечнодышащих из типа полухордовых (ближайших родственников хордовых) два продольных нервных ствола: брюшной и спинной, причем спинной ствол у них частично свернут в трубку, точно так же, как у хордовых. Следовательно, можно предположить, что у предка хордовых было два нервных ствола, как у современных полухордовых, но затем брюшной ствол редуцировался и остался только спинной. И в этом случае гипотеза “перевернутости” не нужна.
Расположение сердца - признак более надежный. Заметим, что направление кровотока у кольчатых червей, членистоногих, полухордовых и хордовых (если последних перевернуть) совпадает. Однако многие беспозвоночные лишены кровеносной системы, поэтому считается, что разное расположение сердца (снизу у хордовых и сверху у беспозвоночных) связано с независимым возникновением у них кровеносной системы.
Среди других анатомических признаков, подтверждающих “перевернутость” хордовых, отметим лишь расположение печени: над средней кишкой у беспозвоночных и под - у хордовых [6].

Расположение желтка (показано цветом) у зародышей лягушки (вверху) и дрозофилы.
Передний конец тела обращен влево, спинная сторона - вверх.
Кроме того, гипотеза “перевернутости” хордовых подтверждается в результате сравнения богатых желтком зародышей наземных членистоногих и таких позвоночных, как рыбы, земноводные, рептилии и птицы. У членистоногих желток всегда располагается на спинной стороне зародыша, а у позвоночных - всегда на брюшной [7]. Строение таких зародышей становится сопоставимым, только если признать гомологию спинной стороны позвоночных брюшной стороне беспозвоночных.
Молекулярная биология развития
В последние годы убедительные доказательства гипотезы “перевернутости” хордовых получены молекулярными биологами [8]. У зародышей позвоночных (лягушки и рыбы) на стадии гаструлы на спинной стороне синтезируется белок CHD (chordin), а на брюшной - белок BMP-4 (bone morphogenetic protein-4). У насекомых (дрозофилы) на стадии гаструлы на спинной стороне синтезируется белок DPP (decapentaplegic), а на брюшной - белок SOG (short gastrulation).
Формирование структур и органов спинной и брюшной сторон тела у позвоночных и насекомых определяется соотношением концентраций упомянутых белков. Так, у зародыша лягушки в норме белок CHD синтезируется на спинной стороне, но если в опыте повысить его концентрацию на брюшной с помощью инъекции, необходимой для его синтеза матричной РНК, то на этом месте в дальнейшем разовьются структуры спинной стороны. Наоборот, искусственное повышение концентрации белка BMP-4 на спинной стороне зародыша вызывает развитие там структур брюшной стороны. Аналогичные результаты были получены и на дрозофиле: формирование структур спинной стороны связано с повышенной концентрацией белка DPP, а брюшной - SOG.
Выяснилось также, что в белках CHD и SOG оказались одинаковыми 28% аминокислотных последовательностей, а из 100 аминокислот C-концевых частей молекул белков BMP-4 и DPP, отвечающих за их функционирование, совпадают 76%. Даже 28%-ая гомология сравниваемых белков свидетельствует о несомненном общем происхождении, от одного и того же белкового предка. Таким образом, “спинной” CHD позвоночных гомологичен “брюшному” SOG дрозофилы, а “брюшной” BMP-4 позвоночных гомологичен “спинному” DPP дрозофилы. Но ведь эти гомологии - не что иное, как подтверждение гипотезы “перевернутости” хордовых!
Удивительна эволюционная консервативность функции рассматриваемых белков, которая проявляется в том, что белки лягушки CHD и BMP-4 функционально активны в зародыше дрозофилы, а белки дрозофилы SOG и DPP - в зародыше лягушки. Экспериментально показано, что “брюшной” белок лягушки BMP-4 вызывает у зародыша дрозофилы формирование структур спинной стороны, а “спинной” белок лягушки CHD приводит к формированию зародышем дрозофилы структур брюшной стороны. Наоборот, белки дрозофилы - “брюшной” SOG и “спинной” DPP - соответственно вызывают формирование зародышем лягушки структур спинной и брюшной сторон. А ведь, согласно современным оценкам, с момента расхождения эволюционных линий, ведущих к насекомым и хордовым, прошло более полумиллиарда лет! Такая консервативность белков делает гипотезу “перевернутости” хордовых еще более правдоподобной.
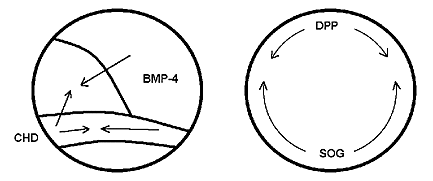
Диффузия (направление показано стрелками) белков, определяющих в начале гаструляции формирование спинной и брюшной сторон у зародышей позвоночных и насекомых: лягушки (слева) и дрозофилы. У зародыша лягушки спинная сторона определяется повышенной концентрацией белка CHD в области так называемого организатора (спинной губы бластопора), а брюшная - BMP-4 на противоположной организатору стороне зародыша. У зародыша дрозофилы спинная сторона определяется повышенной концентрацией белка DPP и брюшная - SOG. Так как в парах CHD/SOG и BMP-4/DPP белки гомологичны, можно сделать вывод, что спинная сторона лягушки соответствует брюшной стороне дрозофилы и, наоборот, брюшная сторона лягушки соответствует спинной стороне дрозофилы. Эти гомологии подтверждают гипотезу “перевернутости” хордовых.
Существуют и другие, дополнительные свидетельства в пользу гипотезы, полученные в результате сопоставления некоторых генов, из которых мы упомянем лишь о двух [9]. Развитие продольного нервного ствола связано с активностью вдоль его средней линии гомологичного для разных животных (насекомых, позвоночных, круглых червей) гена netrin: соответственно положению нервной системы этот ген активен на брюшной стороне у насекомых и на спинной - у позвоночных. Таким образом, брюшной нервный ствол беспозвоночных гомологичен спинному стволу позвоночных. Развитие сердца у дрозофилы определяется синтезом белков TINMAN и DMEF2, а у позвоночных - гомологичных им белков. Следовательно, расположенное на спинной стороне сердце насекомых и на брюшной стороне сердце позвоночных также оказываются гомологичными.
Итак, благодаря современным данным молекулярной биологии, сравнительной анатомии и эмбриологии подтвердилась гипотеза “перевернутости” хордовых, выдвинутая Жоффруа Сент-Илером еще в начале прошлого века.
Список литературы
1 Жоффруа Сент-Илер Э. О позвонке у насекомых // Избр. тр. / Ред. И.Е.Амлинский. М., 1970. С.375 - 390.
2 Жоффруа Сент-Илер Э. О естественных отношениях Маки (Makis lemur L.) и описание нового вида млекопитающих, сделанное гражданином Жоффруа, профессором зоологии при Музее естественной истории // Там же. С.11.
3 См.: Амлинский И.Е. Начальный этап сравнительно-морфологического обоснования единства животного мира // Там же. С.539 - 642.
4 Дорн А. Происхождение позвоночных животных и принцип смены функций. М.;Л., 1937.
5 Ромер А., Парсонс Т. Анатомия позвоночных. М., 1992. Т.1. С.43.
6 Подробнее см.: Малахов В.В. Новый взгляд на происхождение хордовых // Природа. 1982. №5. С.12 - 19; Малахов В.В., Попеляев И.С., Галкин С.В. // Биология моря. 1996. Т.22. №6. С.339 - 345; Иванова-Казас О.М. // Там же. 1997. Т.23. №4. С.247 - 254; Она же. // Онтогенез. 1998. Т.29. №5. С.373 - 385; Nubler-Jung K., Arendt D.J. // Zool. Syst. Evol. Research. 1999. V. 37. P. 93 - 100.
7 Иванова-Казас О.М. Эволюционная эмбриология животных. СПб., 1995.
8 Arendt D., Nubler-Jung K. // Nature. 1994. V.371. P.26; Robertis E.M.de, Sasai Y. // Ibid. 1996. V.380. P.37 - 40; Ferguson E.L. // Curr. Opin. Genet. Devel. 1996. V.6. P.424 - 431.
9 Robertis E.M.de, Sasai Y. // Ibidem.