Влияние глауконита на продуктивные качества гусят-бройлеров
СОДЕРЖАНИЕ
ВВЕДЕНИЕ
1. ОБЗОР ЛИТЕРАТУРЫ
Особенности физиологии пищеварения сельскохозяй ственной птицы
Основные факторы, влияющие на продуктивность гусят — бройлеров
Влияние природных алюмосиликатов на обмен веществ сельскохозяйственной птицы
Изменение продуктивности птицы под влиянием кормовой добавки цеолитов
Заключение по обзору литературы
2. СОБСТВЕННЫЕ ИССЛЕДОВАНИЯ
2.1. Методика и условия проведения исследований
Химический состав используемого глауконита
Методика, схема и техника проведения исследований
Методика и техника проведения балансового опыта
Методики лабораторных исследований
Содержание и кормление гусят - бройлеров
3. РЕЗУЛЬТАТЫ СОБСТВЕННЫХ ИССЛЕДОВАНИЙ
3.1 Динамика живой массы и сохранность гусят - бройлеров
за период откорма
3.2 Физиологические исследования
Влияние глауконита на переваримость питательных веществ рациона
Баланс и использование азота
Баланс кальция и фосфора
Гематологические исследования
Результаты контрольного убоя гусят - бройлеров
3.5 Конверсия протеина и энергии корма в питательные
вещества мясной продукции
3.6 Затраты корма и экономическая эффективность проведённых
исследований
РЕЗУЛЬТАТЫ ПРОИЗВОДСТВЕННОЙ ПРОВЕРКИ
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ СОБСТВЕННЫХ
ИССЛЕДОВАНИЙ
ВЫВОДЫ
ПРЕДЛОЖЕНИЯ ПРОИЗВОДСТВУ
ВВЕДЕНИЕ
Актуальность темы выпускной квалификационной работы. Птицеводство является одной из отраслей народного хозяйства, которое первое перешло на промышленную основу. Интенсивный путь развития отрасли позволил во многом решить проблему обеспечения населения страны яйцом и значительно увеличить производство мяса птицы.
В структуре себестоимости продукции птицеводства наибольший удельный вес занимают корма. Поэтому основным путём снижения себестоимости продукции птицеводства является кормление птицы полнорационными сбалансированными комбикормами, позволяющим обеспечить потребность птицы в нормируемых элементах питания.
Одними из широко используемых кормовых добавок природного происхождения являются цеолиты вулканического и осадочного происхождения. Эффективность их использования в птицеводстве доказана научными работами Г.А.Романова (1991), В.Фисинина (1990), В.Н.Николаева (1998), A.M. Шадрина (2000), S. Fugii (1974), F.Mumpton et P.Fishman (1974) и другими. Разработка в нашей стране новых месторождений гидрослюд, алюмосиликатов заставляет с научной точки зрения подходить к их практическому применению. В Челябинской области за последние годы интенсивно ведутся разработки Потаненского месторождения вермикулита и Карийского месторождения глауконита.
Первые научные работы по использованию глауконита в рационах сельскохозяйственных животных и птицы стали появляться десять лет назад. Научными работами Е.В. Иванова (2001) на свиньях, А.Н. Галатова (1999) -на овцах, Т.С. Кирсановой (2002) - на крупном рогатом скоте определены дозировки ввода глауконита в рацион, то с сельскохозяйственной птицей этот вопрос требует дальнейшего изучения. Единственные исследования
А.А. Замятина (2000) были выполнены на курах - несушках кросса Ломан LSL, и остаётся неизученным вопрос влияния глауконита на другие виды и кроссы птицы, определения оптимальной дозировки его включения в комбикорм в зависимости от концентрации в породе и величины помола.
Поэтому целью данной работы являлось изучение влияния глауконита на продуктивные качества гусят-бройлеров. В задачи исследований входило:
определить оптимальную дозировку глауконита в рационах гусят-ройлеров;
изучить влияние глауконита на изменение живой массы гусят и их сохранность;
установить изменения переваримости питательных веществ рациона под влиянием глауконита;
рассчитать баланс азота, кальция и фосфора;
проследить изменения физиологических и биохимических показателей крови;
влияние глауконита на показатели мясной продуктивности гусят и трансформацию кормового протеина и энергии в продукцию;
рассчитать затраты корма и экономическую эффективность проведённых исследований.
Научная новизна исследований заключается в том, что впервые было изучено влияние глауконита Карийского месторождения на рост и сохранность гусят-бройлеров, переваримость и использование питательных веществ рациона, изменения биохимического статуса в организме птицы, показатели мясной продуктивности, а так же установлена его оптимальная дозировка.
Теоретическое и практическое значение выпускной квалификационной работы. Дано научно-практическое обоснование широкого использования кормовой добавки глауконита в условиях промышленного и частного гусеводства как стимулятора роста, развития, сохранности птицы и повышения её убойных качеств.
1. ОБЗОР ЛИТЕРАТУРЫ
Рост и развитие живого организма, его способность к продлению рода возможно за счёт потребления питательных веществ растительного и животного происхождения, природных и синтетических минеральных веществ, витаминов, ферментов и гормонов. Использование их в организме сельскохозяйственных животных и птицы неодинаково ввиду анатомо-физиологических особенностей пищеварительного тракта. И поэтому освещать обзор литературы по теме нашей работы следует с физиологических особенностей пищеварения сельскохозяйственной птицы.
1.1 Особенности физиологии пищеварения сельскохозяйственной птицы
В результате длительного эволюционного процесса птица заняла особое место в животном мире. Она отличается от млекопитающих экстерьером, строением органов пищеварения, перевариванием корма, размножением, способностью к более высокой оплате корма продукцией (Агеев В.Н. и др., 1987).
В отличие от млекопитающих у птиц принятый корм в ротовой полости смачивается слюной, богатой муцином, проглатывается, а затем попадает в зоб, где он смешивается с водой, слюной, муциносодержащим секретом пищевода и зоба и подвергается частичному воздействию ферментов (амилаз и протеаз), находящихся в корме и выделяемых микрофлорой. Среда корма, как правило, кислая, а рН содержимого зоба значительно ниже 7 (4,5-5,8). Поэтому создаётся благоприятная среда для интенсивных бактериальных процессов расщепления корма.
Содержимое зоба из пищевода поступает в железистый желудок, в котором содержится пепсин, соляная кислота, сычужный фермент и муцин; рН железистого желудка 4,7-3,6 у кур и 3,4 у уток. Из железистого желудка корм перемещается в мускульный желудок, где кормовые массы интенсивно перетираются кутикулой и находящимся в желудке гравием. Кислая среда мускульного желудка (рН 3,9-2,6 у кур и 2,3 у уток) способствует расщеплению легкопереваримых белков и полипептидов, а ферменты микрофлоры продолжают гидролиз углеводов.
При прохождении через тонкий отдел кишечника химус перемешивается с соками кишечника, поджелудочной железы и желчью, что способствует дальнейшему расщеплению основных питательных веществ корма: пептонов, полипептидов и белков под действием протеаз - до аминокислот; углеводов под влиянием инвертаз и амилаз - до моносахоридов; жиров под влиянием липаз и желчи до глицерина и жирных кислот (Агеев В.Н. и др., 1987).
Г.П Мелехин и Н.Я. Гридин (1977) рассматривают переваривание корма в желудочно-кишечном тракте не столько механический, сколько биохимический процесс. К механическому процессу они относят проглатывание корма и его прохождение через пищеварительную систему в результате перистальтики. Биохимический процесс зависит от действия пищеварительных секретов, которые выделяются железами, расположенными в тканях пищеварительного тракта или органа, тесно связанными с пищеварением, такими как печень и поджелудочная железа.
В исследованиях Ц. Батоева (1992) установлено, что у птицы протоки поджелудочной железы и желчевыделения открываются в кишечник через одно отверстие. У кур, уток и гусей выделение сока поджелудочной железы в кишечник происходит непрерывно. Высокий уровень пищеварительной деятельности железы, особенно у гусей и уток, наблюдается при одновременном их кормлении и поении. При этом количество панкреатического сока увеличивается в 2,0-2,2 раза, а между кормлениями его выделяется меньше, чем у кур. Поэтому на 1 кг массы тела у кур и уток выделяется по 28 мл, а у гусей — 16 мл панкреатического сока в течение суток. В то же время у кур активность амилазы намного выше по сравнению с утками и гусями. Так, за первые 30 и 60 минут после приёма корма концентрация амилазы в кишечнике у кур увеличилась соответственно на 184 и 182 %, у уток она составила 146 и 142 %, а у гусей - 166 и 159 %. Концентрация ферментов панкреатического сока птицы намного превосходит секрет свиней и собак по содержанию амилазы в 10-20 раз, протеолитических ферментов - в 4-9 и 1-3 раза.
В силу амилолитической активности суточный объём панкреатического сока способен за 1 минуту гидролизовать такое количество крахмала, которое достаточно для обеспечения дневной потребности 8-10 и более голов птицы. Если принять, что минимальное время действия панкреатического сока в кишечнике всех видов птицы 60 мин., то может гидролизоваться огромное количество крахмала, превышающее суточную потребность кур в углеводах в 560 раз, уток - в 460, гусей - 900 раз. Автор отмечает, что в течение 2 часов после приёма корма на 1 кг массы тела у кур выделяется в 4,3 раза меньше панкреатического сока, но активность амилазы сока у кур в 2,2 раза больше, а протеаз в 1,9 раза, чем у свиней.
В.Ф. Каравашенко (1986) обращает особое внимание на нормирование в рационах сельскохозяйственной птицы сырой клетчатки, так как основное место её переваривания — слепые отростки кишечника, населённые микроорганизмами, выделяющими целлюлозолитические ферменты.
Оптимальным содержанием сырой клетчатки в рационах сельскохозяйственной птицы является: для ремонтного молодняка 7-10 %, бройлеров - 4-5, взрослых кур - 5,5, индеек - 6, уток - 7, гусей - 10% от сухого вещества. По данным И.Т. Маслиева (1968), Э.У. Кремптона и Л.Э. Хариса (1972), переваримость клетчатки в зависимости от вида птицы, возраста, уровня содержания её в рационе составляет от 5 до 30%.
Т.М. Околеловой (1990) установлено, что у гусей из тонкого отдела кишечника в слепые отростки поступают не все кормовые массы, а только жидкая часть с мелкоизмельчёнными частицами корма, в том числе и клетчатка. Минуя слепые отростки они попадают в прямую кишку и выводятся из организма. В пищеварительном тракте гусей, в отличие от других видов птицы корма, особенно объёмистые, в мышечном желудке подвергаются более основательной механической обработке. Сила давления в нём в два раза выше, чем у кур. Число сокращений мышечного желудка у гусей - 5, у кур и индеек - 2,9 раза в минуту. Длина тонкого отдела кишечника у гусей (от выхода желудка до клоаки) 285см, уток -196, у кур -180 см, длина слепой кишки соответственно 23, 10 и 14 см.
Продукты расщепления белков и углеводов, вода, минеральные вещества и витамины всасываются в тонком отделе кишечника. Вода и азотосодержащие вещества всасываются и в слепых отростках слепой кишки. Контроль за процессом всасывания находится в центральной нервной системе, в её продолговатом мозге. Всасывание представляет собой одну из основных функций пищеварительного тракта, которая обеспечивает проникновение через клеточные мембраны стенок пищеварительных органов продуктов ферментативного гидролиза белков, жиров, углеводов, витаминов, макро - и микроэлементов, воды и других поступивших с кормом соединений. Всосавшиеся в кровь и лимфу вещества разносятся по организму и включаются в обменные процессы (Недзвецкий В.К., Бикташев Р.У., 1975).
По мнению В.М. Селянского (1986), процесс всасывания регулируется рефлекторным и гуморальным путём.
Научными исследованиями Г.И. Азимова (1971) и И.Т. Маслиева (1968) установлено, что раздражение гипоталамуса вызывает изменение всасывания в тонком кишечнике, а гормон щитовидной железы тироксин повышает всасывание глюкозы. В результате чего протеин животных кормов переваривается в организме сельскохозяйственной птицы на 85-95 %, растительных - на 80-85 %, усвоение энергии находится на уровне от 65 % у кур до 70-80 % у гусей (Околелова Т.М., 1990; Агеев и др., 1987).
Следовательно, для сельскохозяйственной птицы характерен небольшой удельный вес пищеварительной системы (7-10,0 %) от общей массы тела, но с высокой активностью гидролитических ферментов в отличие от животных с однокамерным и многокамерным желудком.
1.2. Основные факторы, влияющие на продуктивность гусят-бройлеров
Статистические данные, которые приводит С. Смагулов (1989), свидетельствуют, что до революции 1917 года общее поголовье гусей в России превышало 100 млн. голов, из них около 10 млн. экспортировалось за
границу. В 1988 году в нашей стране осталось 2 млн. голов в общественном секторе и 2,5 млн. - в частном. К 1997 году поголовье гусей в России должно было увеличиться до 37,0 млн. голов.
Однако в силу сложившихся обстоятельств в последние годы было допущено значительное сокращение производства птицеводческой продукции. За последние 10 лет производство мяса птицы во всех категориях хозяйств уменьшилось в 2,2 раза. В этой связи отставание от достигнутого уровня 1990 года по производству мяса птицы по всем категориям хозяйств составило 1035 тыс. т., или 57 %. В «Концепция-прогноз развития животноводства России до 2010 года» (Данкверт С.А. и др., 2002) намечено в 2005 году выйти на производство мяса птицы примерно в 1,7 млн. т в убойной массе и поднять потребление мяса птицы в расчёте на душу населения общественного производства с 5,5 кг в 2000 году до 11,5 кг - в 2005. В решении этой задачи значительное место уделяется гусеводству путём создания в регионах страны родительских стад гусей в кооперативных и фермерских хозяйствах для производства инкубационного яйца и получения молодняка для продажи населению.
Гуси были одомашнены с незапамятных времён и получили самое широкое распространение благодаря неприхотливости к условиям содержания и кормления, а так же получения исключительно ценных продуктов питания (Шпекторов В., 1984).
Н.С. Ковацкий и др. (1990), Б.Ф. Бессарабов, (1994) обращают внимание и на то, что гуси способны потреблять и переваривать в больших количествах корма с повышенным содержанием клетчатки. Гуси обладают и прекрасными фуражировочными способностями, потребляя на пастбище до 2 кг зелёной массы в сутки. Такие особенности этой птицы позволяют расширить кормовую базу отрасли, экономить значительное количество зерна (Шпекторов В., 1984).
Поэтому немаловажным является повышенный спрос в нашей стране и за рубежом на такую продукцию гусеводства, как жирная гусиная печень, гусиный жир, перо-пуховое сырьё, гусиные пуховые шкурки.
По данным А. Зелятрова (1984), интенсивный откорм гусят на мясо наиболее развит в Венгрии, Чехословакии, Румынии, Болгарии, Польше. При этом гусеводство в зарубежных странах не является ведущей отраслью мясного птицеводства. В США, Канаде, странах ЕЭС на долю гусей приходится всего 0,5-1,0 % валового производства мяса птицы. Но, тем не менее, за 20 лет производство мяса гусей в мире возросло на 25 %, а в некоторых странах (Венгрия, Чехословакия, Польша, Франция) — в 2-4 раза.
При этом гусеводство в данных странах начинает приобретать интенсивные формы развития (Aitken J, Merrit E., 1967; Bielinska К., 1974; Jamani К., Marai J., Szabo J., 1973; Merten M., 1973; Rydlova F., 1973$ Boold F.,1974).
Однако гуси, по мнению В. Шпекторова (1984), по сравнению с другими видами птицы обладают гораздо более низкими
воспроизводительными способностями. Это и послужило одной из основных причин того, что после перевода птицеводства на промышленную основу гуси не смогли конкурировать с курами мясного направления и другими видами мясной птицы. Их поголовье стало быстро снижаться. Разработка учёными и практиками интенсивных методов ведения гусеводства (получения двух циклов яйцекладки в год, откорма гусят-бройлеров и т.д.) коренным образом изменило положение отрасли.
По данным П. Салеева (1982,1984), в нашей стране разводят более 20 пород и породных групп гусей, хорошо приспособленных к различным природно-климатическим условиям. По происхождению и хозяйственно-полезным качествам отечественные породы гусей можно разделить на три группы.
Первая группа. Гуси китайского происхождения (китайские, кубанские, переяславские, горьковские) характеризуются высокой яичной продуктивностью, но небольшой живой массой.
Вторая группа. Западноевропейские гуси (тулузские, крупные серые, виштинез, эмденские, рейнские, итальянские) имеют более рыхлую конституцию и сравнительно высокую яйценоскость.
Третья группа. Восточноевропейские гуси (роменские, арзамасские, уральские) имеют высокую жизнеспособность, но низкую продуктивность.
Для производства гусят-бройлеров, выращиваемых девять недель, необходимо использовать наиболее раннеспелые породы: итальянскую, рейнскую, горьковскую породные группы. Все они имеют белое оперение, что даёт возможность реализовать гусят на мясо в ранние сроки. Гуси горьковской породной группы высокопродуктивны, однако нуждаются в значительном улучшении мясных качеств. Наиболее приемлемым для производства гусят-бройлеров являются гуси рейнской и итальянской пород. Их средняя яйценоскость за один продуктивный цикл составляет 45-50 яиц, а при двух циклах - 75 штук. Гусят в 9-10 недельном возрасте сдают на мясо с живой массой 3,8-4,0 кг при затратах корма на 1 кг прироста 3,3-3,5 кг (СалеевП., 1984).
По наблюдениям П. Салеева (1982), гуси имеют высокую скорость роста. Живая масса одного гусёнка с суточного до 56-60 - дневного возраста увеличивается в 40-45 раз и достигает в среднем 4 кг при затрате на 1 кг прироста массы не более 3 кг корма. Причём самые низкие затраты корма у гусят отмечаются в первые три недели жизни (2,35 кг корма и 379 г протеина), в последующие 5-6 недель относительная скорость роста гусят уменьшается, доля поддерживающего корма и оплата корма приростом возрастают до 4,3 кг и 720 г протеина. К 8-9 — недельному возрасту у гусят отмечается высокий выход съедобных частей в тушке, а сама тушка приобретает хороший товарный вид. Более продолжительный срок
выращивания гусят на мясо (до 70-75 дней) в связи с наступлением в этот период ювенальной линьки не желателен, так как образующиеся в процессе ювенальной линьки зачатки новых перьев, так называемые пеньки, очень трудно удаляются при обработке. Товарный вид тушек ухудшается, и продукция переводится в категорию нестандартной (Мамаев В., 1990).
В. Лазер (1977, цит. По П.Салееву, 1982) вывел гибридных гусят, которых можно реализовать на мясо в 7-8 недельном возрасте с живой массой 4-4,5 кг при затратах 2,8 кг корма на 1 кг прироста живой массы. При закладке материнских линий применялись гуси итальянской и рейнских пород и так называемых GLUv =f и GLUv=L гусей. Для создания отцовских форм использовались в основном популяция гусей ландской, словацкой белой и массёб. Самые лучшие результаты В.Лазер получил от скрещивания самцов ландской с самками рейнской пород, живая масса гусят в 56 — дневном возрасте была 4,125 кг, убойный выход - 68,6 %, масса грудных мышц - 10,6 %, масса бедренных мышц - 19,4 %, затраты корма на 1 кг прироста живой массы - 3,01 кг и самые низкие показатели были получены от скрещивания самцов чешской с самками итальянской пород: 3,485 кг живая масса, 70,9 % убойный выход, 10,3 % масса грудных мышц, 17,8 % -масса бедренных мышц, 3,28 кг — затраты корма на единицу продукции.
На селекционно-племенной станции по гусеводству в Гробонево (цит. по П. Салееву, 1982) создан двухлинейный кросс, состоящий из материнской линии UV003 и отцовской UV002. Материнская линия создана с участием белой израильской и словацкой пород. Живая масса гибрида в 56- дневном возрасте составила3541 г.
По данным С. Шонина (1992), интенсивный откорм гусят, завезённых из Венгрии в Башкортостан в восьминедельном возрасте, позволил получить живую массу 4,4 кг и хорошо приспособленных к пожизненному ощипыванию пера.
При сравнительном откорме гусаков и гусынь кубанской и крупной серой пород Б.В. Смирновым (1978) было установлено, что наибольшую живую массу в 30-дневном возрасте имели гусаки крупной серой породы -2369 г, в 60 - дневном возрасте - 4500 г, в то время как кубанские гусаки имели живую массу соответственно 1881,3 и 3850,8 г. Гусыни крупной серой породы по росту и развитию также превосходили кубанских гусынь и имели живую массу в 30 дней 2075,0 г, в 60 дней - 3700 г, а кубанские соответственно 1710,4 и 3276,7 г.
Целесообразность снимать гусей с откорма, в 50-60 - дневном возрасте подтвердила в своих исследованиях Е.И. Ионова (1971,1973) на гусятах горьковской породной группе и на белой итальянской породе, когда наивысший среднесуточный прирост живой массы был получен в 40-дневном возрасте (69,1 г у самок и 81,4 г у самцов), в 60 - дневном он снизился до 54,9 и 59,9 г соответственно, а в 65- дневном возрасте он составил 41,4 и 57,5г.
По данным В.Ф. Каравашенко (1986), нормативными показателями живой массы 1 головы гусят в 9- недельном возрасте, выращенных на комбикормах с использованием травяной муки, зелёных и сочных кормов, являются: для крупной серой породы 4380 г, оброшинской серой - 4760, горьковской - 3650, итальянской -3850, помеси - 3950 г.
Следующий фактор, оказывающий, на наш взгляд, существенное влияние на продуктивность гусят-бройлеров, является полноценное и сбалансированное кормление.
О.Х. Геворкян и Р.П. Дорофеева (1981) при откорме гусят-бройлеров по периодам выращивания сравнивали норму сырого протеина, рекомендованную ВНИИТИП и 18,22 и 24 % в первые 20 дней, 16, 20 и 22% протеина в 21-65 день выращивания при концентрации обменной энергии 1173 кДж. При этом было установлено, что наибольший интенсивный рост и более высокую оплату корма продукцией имели гусята, выращенные при
содержании в комбикорме в период с 1 по 20 день 22% сырого протеина и 1173 кДж обменной энергии при ЭПО 533; во второй период (21-65 день) соответственно 20 %, 1173 кДж и ЭПО 587 кдж на один процент сырого протеина.
Учитывая, что энергия роста у гусят в первый месяц жизни составляет 200 %, во второй - 96, в третий - 17 %, поэтому Т.М. Околелова (1990) рекомендует выращивать гусят на мясо интенсивным способом до 65
- дневного возраста. При этом в комбикормах для первого возраста (1-3 недели) предусматривается более высокий уровень сырого протеина (20 %) и незаменимых аминокислот. В возрасте 4-8 недель содержание его снижается до 18 %, а содержание клетчатки увеличивается до 6 %. Среднесуточное потребление комбикорма гусятами по неделям выращивания составляет (на одну голову в сутки, г): в возрасте одной недели - 50; 2-90; 3-110; 4-220; 5- 270; 6-280; 7-328; 8 недели -328 г.
В.Ф. Каравашенко (1986) рекомендует на протяжении всего периода откорма гусят концентрацию энергии в 100 г комбикорма выдерживать на уровне 280 ккал, или 1,172 МДж., а сырого протеина - 20% в первые три недели и 18 % с 4 по 8 неделю откорма. Мясным гусятам лучше скармливать гранулированные комбикорма в начале выращивания в виде крошки, а с 3-недельного возраста гранулы диаметром 4-6 мм.
Г. Закревская (1972) , Н. Мээл (1970 , П. Ф. Салеев (1985), Э.И. Дергулян и др. (1982), G. Lnaniecka et. al. (1973), R. Qvilici (1968) считают, что в рационе гусят - бройлеров должно быть 275-290 ккал ОЭ в возрасте 1-20 дней , 290-300 ккал - 21 - 56 дней, 300 - 330 ккал - 57 -65 - дневном возрасте . Сырой клетчатки не более 5,5 - 6,5 % .Причем в комбикорма для 1 -20-дневных гусят необходимо включать: зерновые 65-75%, жмыхи и шроты
- 12-15, животные корма - 7-10, дрожжи - 2-3, травяную муку - 4- 5, минеральные корма - 1,0 -1,5 %. С 21 до 65 дня количество животных
кормов уменьшают до 3-5 %, а минеральные и зерновые увеличивают до 1,8 -2,2% и 78-83%.
По данным А.В. Зелятрова (1975), в Польше для выращивания гусят до 3-недельного возраста используют комбикорм: кукуруза 39,98%, ячмень-16, горох — 17, льняной шрот — 10, люцерновая мука — 10, рыбная мука — 4, дикальций фосфат -1,0, мел - 1,0, минерально-витаминный премикс - 1,0%, синтетический L-лизин, ДЬ-метионин.
В Венгрии комбикорм для гусят - бройлеров состоит из кукурузы -41%, ячменя - 13, пшеницы - 3, сои - 6, подсолнечнака-6, рыбной муки-3, люцерновой муки—12, гороха -5, дрожжей-3, минеральных веществ-8% (MolnarJ., PacsJ.,1973).
Не плохой результат был получен J. Bogre (1969) при замене в рационе гусят-бройлеров животного белка растительным. В опытной группе до 10 дня получали комбикорм, что и в контрольной с соотношением животного белка к растительному 1:4, а с 11 дня опытная группа получала только растительный белок. Средняя живая масса одной головы к концу откорма составила 4218 г в контрольной группе, 4274 г - в опытной, при затратах корма на 1кг прироста соответственно 2,85 и 2,92 кг.
Однако Л. Лепайые и др. (1982,1983) при снижении в последнюю девятую неделю откорма гусят уровня сырого протеина в комбикорме с 18,5 до 15,8 % получали практически одинаковую живую массу гусят (4,6- 4,4 кг), выход съедобных частей в тушке 53-54%, выход на 1 кг живой массы пищевого белка составил 86-87г, энергии мяса 7,5-8,8 МДж. При этом коэффициент конверсии протеина в возрасте гусят 0 — 9 недель составил 10-17%, коэффициент конверсии энергии - 10-25%.
Третьим основным фактором, влияющим на продуктивность гусят-бройлеров, является способ их выращивания и условия содержания.
До 60-х годов прошлого века в нашей стране применялась система выращивания гусят с пастбищным содержанием их до 180-дневного возраста. На смену ей был подложен способ выращивания гусят на мясо в летних лагерях, предусматривающий наряду с кормлением молодняка концентрированными кормами широкое использование зелёных кормов. В результате чего сроки выращивания гусят на мясо сократились с 150-180 дней до 75 дней. Переход на круглогодовое производство мяса гусей требовал интенсивных способов выращивания гусят на мясо, с учётом безвыгульного содержания (Салеев П., 1982).
На основании разработок, проведённых во ВНИТИПе, рекомендуются следующие варианты технологии круглогодового производства гусиного мяса:
Выращивание гусят в птичниках на полу с использованием подстилки.
Выращивание гусят в птичниках, оборудованных сетчатыми полами.
Выращивание гусят в клетках.
4. Выращивание гусят в летних лагерях под облегчёнными навесами и на специально оборудованных откормочных площадках.
В зависимости от способа выращивания гусят на мясо (на сетчатых полах, на полу с использованием подстилки или в клетках или в сочетании этих способов) различают напольную, на сетчатых полах, клеточную и комбинированную системы.
По данным В.А. Покровского (1962), разработанный ВНИТИП метод интенсивного выращивания гусят на мясо в базах (без пастбища) позволяет к 75-80 дню иметь живую массу гусят 4-4,5 кг с затратами на 1 кг прироста не более 2,5 кг концентратов, 5 кг зелени, т.е. всего 4
корм. ед. Наилучший прирост гусята дают в первые 2 месяца 1,8-1,9 кг, на третий - меньше 0,8 кг. По данной технологии с 1 по 20-30 день выращивания гусят содержат на глубокой подстилке.
Б.В. Смирновым (1978) были проведены исследования при сравнительном выращивании гусят в клетках и на полу. При одинаковой постановочной живой массе гусят на опыт наилучшие результаты (4172,5 г) имели группы, где гусята первые 20 дней выращивались в клетках, а с 21 по 75 день на полу. Самые низкие результаты по живой массе (3276,3 г) имели гусята, которые все 75 дней содержались в клетках, а те, которые были выращены только на полу, имели живую массу 3675,3 г.
Однако Е.И. Ионова и Т.А. Столляр (1872) в сравнительном опыте по выращиванию гусят на мясо в безвыгульных условиях на глубокой подстилке, планчатых полах и клеточных батареях лучшие результаты получили в клетках.
Ряд учёных (Иоцюс Г.П. и Старчиков Н.И.,1989, J. Hrour и др., 1984) предложили ограничить срок выращивания гусят в клеточных батареях до 60 дней, т. к. интенсивность роста птицы в старшем возрасте снижается . Так при плотности посадки 6 гол./кв.м живая масса самцов и самок в 75- дневном возрасте достигает соответственно лишь на 4,0 и 3,2 кг.
В последние годы при интенсивных способах выращивания молодняка водоплавающей птицы на мясо критерием её скороспелости принято считать быстроту оперённости как показатель, имеющий коррелятивную связь с живой массой организма (Альпейсов Ш.А.,1992)
Исследования ВНИТИПа (цит. по П. Салееву, 1982) показали, что оперяемость молодняка зависит также от способа выращивания. Лучшая оперяемость наблюдалась при выращивании гусят в клеточных батареях и на полу с использованием подстилки. Гусята, выращенные на
сетчатых полах, не уступая в живой массе по сравнению с другими способами (в клетках, на подстилке), имели несколько худший рост пера.
Важным моментом в технологии производства гусят-бройлеров является возможность использования водоёмов.
П. Салеев (1985) отмечает, что у гусят, приученных к воде в данном возрасте, ювенальное оперение сменяется быстрее, и в 56-дневном возрасте, их отправляют на убой с живой массой 4-4,5 кг.
К. Bartels et al.(1980), M. Day en и Н. Fiedler (1990) установили, что лучшее развитие перо-пухового покрова и более высокая живая масса гусят были получены в группах, где молодняк имел доступ к водным выгулам. Аналогичного мнения придерживаются чехословацкие учёные Z. Hudsky и E. Machalek (1989), Т. Majewska и A. Faruga (1982), E.Misikova и O.Palanska (1988).
Ш.А. Альпеисов (1988,1988) рекомендует применять систему выращивания молодняка, предусматривающую допуск гусят к сухостойным и водным выгулам. Она способствует улучшению зоогигиенических условий в помещениях и предупреждению расклёва. Пребывание гусят на воде способствует хорошему росту и развитию перьевого покрова.
Относительно условий содержания гусят Е.Ионова (1979) рекомендует в помещении размещать их по 8-10 гол./кв. м площади пола. Температура в помещении в первые пять дней должна быть 30-26°С, с 6 по 10 день 26-23°С, с 10 по 20-23°С и постепенно снижают до 20-18°С. Показателем оптимального обогрева гусят служит их поведение. Если они бодрые, активно двигаются, а после кормления для отдыха располагаются небольшими группами по 3-5 голов, значит температура в помещении нормальная. Воздух должен быть чистым, для чего периодически проветривают помещение. С 5-7 дня гусят можно выпускать на выгул сначала на 20-30 мин., с 3 недель большую часть времени они
должны проводить на выгуле с одновременным приучением их к водоёму.
Н.С. Ковацкий (1990) считает целесообразным выращивать гусят на мясо с разделением их в суточном возрасте по полу. Это позволяет к началу убоя птицы увеличить живую массу самцов на 7- 8 %, самок — на 5-6 % и повысить сохранность поголовья на 3-4%.
K. Bielinska (1973) и J. Bogre (1969) считают оптимальной температурой в помещении гусят 26-3 0°С в первую неделю жизни и 20-26°С - в остальной период выращивания. При этом влажность воздуха должна быть 66-75 %.
J. Molnar и J. Pacs (1973) рекомендуют не превышать плотность посадки гусят в помещении в возрасте 65 дней не более 3,5-6 голов на 1м2. А в период выращивания соблюдать температурный режим в возрасте: 1-3 дня - 30°С, 4-5 - 28°С, 6-7 - 26°С, 8-10 - 24С, 11-21 - 20°С, 22-65-18°С.
Е.И. Ионова (1975) в своём эксперименте получила живую массу гусят по группам в 65-дневном возрасте 3,438 и 3,673 г. При этом в период выращивания - температура в помещении снижалась с 32°С до 29°С к концу первых 5 дней, с 6 по 10 день поддерживалась на уровне 29-26° С, с И по 20 день снижалась с 26 до 20° С и с 21 дня до конца выращивания поддерживалась на уровне 20-16°С. Относительная влажность в первые 10 дней была пределах 68 - 73% , с 11 дня и до конца выращивания - на уровне 75-87%.
В соответствии с отраслевым стандартом на технологию откорма гусят (Ионова Е.,1984) их содержат в птичниках, в секциях по 250 голов со съёмными перегородками высотой 0,6 м. Подстилочный материал насыпают слоем 15 см. Откормленные гусята должны соответствовать по качеству требованиям ГОСТ 18292-72. Фронт кормления при сухом типе должен быть 2 см, влажном-6 см, фронт поения - 2 см. Доступ к воде
постоянный. Освещение в первые дни жизни круглосуточное, с 8 по 20 день уменьшают до 16 часов, затем до 14 часов. Температура в помещении в первые 3-4 недели жизни гусят поддерживается на уровне 22-26°С, под брудером 30-32°С, относительная влажность 65-75%. В последующие дни температура в помещении поддерживается на уровне 18-20°С, влажность та же, скорость движения воздуха-0,4 м/сек.
Таким образом, основными факторами, определяющими успех откорма гусят, являются порода, полноценное сбалансированное кормление, технологический приём выращивания и условия их содержания. Причём технология выращивания гусят-бройлеров выбирается в соответствии с природно-климатической зоной и соответствующей кормовой базой хозяйства.
1.3 Влияние природных алюмосиликатов на обмен веществ сельскохозяйственной птицы
Интенсификации птицеводства способствует широкое использование биологически активных веществ в рационах сельскохозяйственной птицы, повышающих ее продуктивность, способствующих эффективному использованию питательных веществ корма и снижающих затраты кормов на единицу продукции. К одной такой широко используемых кормовых добавок относятся природные алюмосиликаты-цеолиты. Учитывая, что механизм действия цеолитов на организм птицы в основной степени изучен, то основные положения его будут общими и для глауконита, так как они относятся к одной группе природных алюмосиликатов. По данным С.Г. Кузнецова (1994), разведанные ресурсы природных цеолитов на территории России составляет 8-10 млрд.т. Только для птицеводства годовая потребность в цеолитах исчисляется в 190 тыс.т ( Романов Г.А., 1991 ).
Основой для использования цеолитов в птицеводстве послужили публикации Японского исследователя Т. Онаги (1966) о высокой биологической активности минералов, скармливаемых цыплятам.
Изучение проблемы применения цеолитов в сельском хозяйстве в России начали с середины 70-х годов, но только с 1987 года были развёрнуты комплексные исследования (Романов Г.А., 1993 ).
По данным В. Фисинина и др. (1990), строительной единицей цеолита является тетраэдр, центр которого занят атомами Si и А1, а в вершинах расположены четыре атома кислорода. Каждый атом кислорода является общим для двух тетраэдров. Замена четырёхвалентного кремния на трёхвалентный алюминий определяет отрицательный заряд каркаса, который компенсируется зарядами одно- и двухвалентных катионов, расположенных вместе с молекулами воды в каналах структуры. Катионы, находящиеся в каналах, легко замещаются, поэтому их называют обменными в отличие от кремния и алюминия, которые не обмениваются в обычных условиях и называются каркасными. В полости по каналам могут проникать и задерживаться различные катионы и молекулы. Таким образом, цеолиты способны выполнять роль «молекулярных сит», отделяя малые молекулы, которые могут проникать в их каналы, от более крупных, которые туда не проникают.
Е.И. Ромашевская, Б.Т. Величковский (1990), В.И. Бгатов, А.М. Паничев (1985 ), Л. Врзгул (1986 ), А.М. Шадрин и др. (1986) считают, что влияние природного цеолита на организм птицы зависит как от их физико-химических свойств, так и от условий кормления и содержания птицы. Геометрия кристаллов важный фактор, определяющий возможность их использования. У клиноптилолита и гейландита, имеющих таблетчатую или пластинчатую форму кристаллов, не обнаружено отрицательных эффектов на уровне целого организма. С другой стороны, эрионит, имеющий волокнистые кристаллы, попадает с пылью в верхние дыхательные пути и может вызвать раковые опухоли (Artvinli M., Baris T.J, 1979 , Nikolova S.,1981,PoolA. et. al., 1983). На ионообменные и молекулярно-ситовые свойства природных цеолитов, по мнению В.И. Бгатова и А.М. Паничева (1985), влияют молекулярная структура цеолита (размер пор, геометрия каналов, внутренний объём и поверхность), внешние условия, химический состав растворов, химуса или газовой смеси, температура и давление.
Учёными ВНИТИП и ВНИИФБП (Методические рекомендации, 1990, Калачнкж Г.И., 1990) установлена эффективность действия цеолитовых туфов в зависимости от уровня минерала в них. Для птицы содержание цеолита в породе должно быть не менее 55%. Скармливание туфов с содержанием цеолита ниже этого уровня не даёт положительного эффекта, хотя и не оказывает выраженного отрицательного действия на обмен веществ и продуктивность. В многочисленных исследованиях было доказано, что чем лучше сбалансирован рацион по питательным веществам, тем ниже эффект от добавления цеолита.
Зависимость эффективности действия цеолита от качества кормов сложна и неоднозначна. Во-первых, при низком содержании сырого протеина в корме у птицы —13% отмечаются не высокие эффекты (увеличение прироста яйцекладки), но экономия кормов существенная - 5-7% (Шадрин A.M. и др., 1986, 1986, Шадрин A.M. и Подъяблонский A.M., 1984).
Скармливание цеолитов при высоком содержании протеина в кормах (17-18%) вызывает значительно выше прирост у бройлеров (до 14-16%) (Русских А.П. и др., 1986, Николаев В.Н., 1988,1988). «Зона оптимальности» по содержанию протеина в кормах птицы - 13-18 %.
В методических рекомендациях «Природные цеолиты в кормлении животных» (1991) обращается особое внимание на крупность помола цеолитовой муки. Цеолитовая мука с крупностью помола менее 0,5 мм при положительном действии на рост птицы вызывает повышенную запылённость комбикорма, что снижает его поедаемость. При измельчении до «респирабельных» размеров большинство цеолитов обладает цитотоксичностью, а при попадании в дыхательные пути вызывает фибротические процессы в лёгких. С другой стороны, порошок с величиной частиц более 2 мм плохо смешивается с комбикормом. Оптимальный размер частиц цеолита составляет 1-2 мм.
Химический состав цеолитовых туфов на территории России имеет различие, следовательно, и биологическое действие их на живой организм будет не одинаковым.
Например, по данным В.К. Горохова и др. (1984), в кристаллической решетке цеолита Лютогского месторождения Сахалина ряд катионов ( Са2+, Mg 2+, K+, Na+) находятся в обменном состоянии и за счёт цеолита потребность цыплят в данных катионах будет покрываться на 30-100%. Существенную роль в минеральном питании цыплят природные цеолиты могут играть за счёт изменения микроэлементного состава пищи. Содержание большинства микроэлементов в цеолитах сопоставимо с содержанием их в осадочных породах (мел, известняк) (Виноградов А.П.,1962), однако в цеолитах они находятся в обменном состоянии и могут быть доступными для животных.
Природные цеолиты, по мнению В.К. Горохова и др.(1977), можно использовать как носители микроэлементов, если предварительно обработать их растворами солей. Статическая обменная ёмкость сахалинских цеолитов составляет 1-1,5 мг-экв/г, что позволяет вводить микроэлементы в количествах, полностью обеспечивающих потребность птицы.
Следует упомянуть и тот факт, что цеолиты полностью безвредны, не содержат ядовитых веществ и заражённость их микроорганизмами исключена ( Чонти Р. и др., 1979, Челищев П., Челищева Р., 1978 ). Кроме того, по данным В.В. Устенко и др. (1994), цеолиты снижают уровень свинца в мышечной ткани птиц в 1,5, а кадмия - в 12,6 раза.
В физиологических исследованиях В.В. Байракова и др. (1984) при добавлении к комбикорму цыплят-бройлеров 6% клиноптилолита установили, что он способствует лучшей переваримости основных питательных веществ корма. С помощью полуколичественного спектрального анализа были изучены изменения микро- и макрокомпонентного состава исходных и прошедших через желудочно-кишечный тракт цеолита, а также зол комбикорма, мяса, печени, костей, помёта и пришли к выводу, что цеолит выполняет роль ионообменника, регулируя соотношение в организме птицы кальция и натрия, улучшает снабжение мягких тканей железом и особенно печень.
В литературе имеются данные, что минералы такие как кварц в форме песка или гравия, влияют на прирост живой массы (Ван-Соест П., 1971).
Учитывая, что гравий является постоянным компонентом рациона сельскохозяйственной птицы К.Я. Мотовилов и В.И. Бгатов (1997) провели сравнительный эксперимент по эффективности использования в организме птицы гравия, цеолита, горного хрусталя и кварца. Если до скармливания размер частиц всех природных соединений был в приделах 3-4 мм, то в группе, где птица получала кварц, с помётом выделилось 55,9% частиц размером 3 мм и более, 19,6% - 2-Змм, 5,7% - 1-2мм, в группе с горным хрусталём соотношение частиц соответственно составило 26,3%, 53,2 и 5,7%. Наибольшие изменения наблюдались в группе с цеолитом; полностью отсутствовали частички размером 3 мм и более, фракции 2-Змм было всего 7,4%, в основном выделялись частицы от 0,1 до 2мм. Гравий, попадая в пищеварительный тракт птицы, под воздействием соляной кислоты, пищеварительных соков, сокращений мышечного желудка претерпевает существенные изменения. Соединения кальция, входящие в состав гравия, растворяются и на их месте образуются впадинки. Изучив химический состав до и после прохождения его через желудочно-кишечный тракт авторы установили достоверное обогащение цеолита фосфором, калием, стронцием и натрием, количество которых соответственно возросло в 3,9%; 2,6; 1,9 и на 33%. Однако, в цеолите снизилось содержание кальция на 46,8%, железа на 22,7, титана на 21,7 , марганца на 18,7 и кремния на 1,19%. Кварцит и горный хрусталь по химическому составу до и после прохождения пищеварительного тракта существенно не различались. Образование минеральных веществ в растворимой части помёта обусловлено тем, что кварцевые соединения в пищеварительном тракте птицы образуют в водной среде гидроксилизированную плёнку кремниевой кислоты, обладающую мощными сорбционными свойствами. В результате механического воздействия плёнка легко срывается и на вновь образованной поверхности частичек минерала образуется новая плёнка, вступающая в хемосорбционные процессы. В связи с тем, что дробление, истирание веществ происходит беспрерывно, идёт постоянное образование в растворимой части адсорбатов. Наиболее высокое количество минеральных соединений в растворимой части при потреблении кварца и горного хрусталя связано, по-видимому, с тем, что содержание кремния в цеолите значительно ниже, чем в кварцитах, меньше образуется и кремневой кислоты, сорбирующей соединения натрия, калия, фосфора и др.
Т. Ленкова и О. Синцерова (1985) считают, что само минеральное вещество цеолитов не участвует в пищеварении, однако благодаря высокой способности связывать воду и ионы процесс пищеварения катализируется. Скорость прохождения корма по желудочно-кишечному тракту птицы под влиянием цеолитов замедляется, что способствует более полному перевариванию и использованию питательных веществ. С другой стороны цеолит выполняет роль ионообменника и регулирует соотношение в организме кальция и натрия, улучшает снабжение мягких тканей железом, в печёночных клетках отмечается накопление гликогена, а в ультраструктурах наблюдаются изменения, указывающие на активизацию внутриклеточного биосинтеза. Повышение усвояемости корма под влиянием цеолита может быть связано, по мнению авторов, с внесением легкоусвояемой подвижной формы калия и кальция, некоторых микроэлементов, буферным эффектом туфа, стабилизирующим кислотность желудочного сока, а также поглощением и выносом из организма токсических продуктов пищеварения и токсинов кормов (Челищев Н.Ф.,1984, Челищев Н.Ф., Челищева Р.В.,1978,1980)
Аналогичные результаты, свидетельствующие об усилении минерального обмена в организме птиц под влиянием цеолита, сопровождающиеся повышенным содержанием макро- и микроэлементов в тканях и органах, были получены В.Н. Николаевым (1988) и S. Fugii (1974).
За счёт локализации в полостях и каналах каркаса катионов натрия, калия, кальция, магния и других элементов цеолиты изменяют ионный состав химуса, нормализуя таким образом рН среды и оптимизируя условия деятельности пищеварительных ферментов. Ряд авторов ( Миначев Х.М, 1977, Врзгула Л.,1986, Калюжнов В.Т. и др., 1988) отмечают повышение протеолитической и амилолитической активности химуса кишечника, что косвенно свидетельствует о каталитической функции цеолитов в кишечнике.
По данным Н.Ф. Челищева (1984) и Д. Брэк (1976), A.M. Караджян и др. (1984), изменение ионного состава химуса, связанное с повышением содержания в нём кальция, способно усилить всасывание слизистой оболочкой продуктов расщепления белков, жиров и углеводов, о чём свидетельствует увеличение коэффициента усвоения этих компонентов кормов.
Активная поверхность природных цеолитов оказывает своё влияние на ферментативную активность пищеварительных ферментов, а молекулярно - ситовые свойства и сорбционная активность способствует связыванию цеолитами токсинов кишечника и крови (Цицишвили Г.М. и др.,1985, Кобидзе Т.С. идр.,1984).
В методических рекомендациях «Использование природных цеолитов в птицеводстве» В. Фисинин с соавторами (1990) отмечают, что при введении 6% клиноптилолита Сокирницкого и Крайниковского месторождения в комбикорм бройлеров переваримость органического вещества повышается с 77,4% до 81,6%, сырого протеина с 90,5% до 94,8%, жира - с 63,6 до 74,7%, а усвоение азота увеличивается с 48,8 до 64,3%. В опытах на бройлерах с туфом Дзегвского месторождения использование энергии рациона возрастает на 4-5%, усвоение азота на 2-3, переваримость сырого протеина на 1-1,5, жира - на 3,0, сырой клетчатки - на 10-15%. В тушках убитых бройлеров масса мышечного желудка к массе тела была на 8,6-12,5% больше, по сравнению с контрольной группой, а отношение массы желудочно-кишечного тракта к массе тела превосходила контроль на 3,4-7,0%. Различий в массе печени, сердца и почек к массе тела между группами отмечено не было. Гистологические исследования показали стимулирующее влияние цеолита на двигательную функцию кишечных ворсинок, что способствовало лучшему усвоению питательных веществ корма.
Кроме этого, цеолит в рационе цыплят-бройлеров способствовал лучшему усвоению жирорастворимых витаминов. Так, содержание в печени бройлеров опытной группы витамина А составила 55,4-114,0 мкг/г, а каратиноидов - 2,56-11,56 мкг/г, по сравнению с контрольной группой, у которой витамина А было на уровне 54,5 мкг/г, а каратиноидов -2,32 мкг/г. Добавка цеолита в рацион кур-несушек увеличивает содержание витамина Е в яйце на 6,5-19,6%, витамина А- 5,4-6,9%.
Газожидкостные хроматографические исследования некоторых аминокислот и пептидов, извлечённых из кормовой массы отдельных участков желудочно-кишечного тракта цыплят показали, что ввод в их рацион цеолита значительно снижает процесс брожения, происходящий в кормовой массе в период переваривания. За счёт локализованных в полостях и каналах каркаса катионов натрия, калия, кальция, магния и других элементов цеолиты изменяют ионный состав химуса, нормализуют рН и оптимизируют условия деятельности пищеварительных ферментов. При этом повышается активность ферментов желудочно-кишечного тракта - липазы и пепсина. Они стимулируют гипертрофию митохондрий и канальцев гладкой цитоплазматической сети, ответственной за синтез гликогена в печени.
З.А. Зубарашвили и Н.Т. Макаридзе (1985) установили, что в кормовом комке дистального участка желудочно-кишечного тракта цыплят, получавших природные цеолиты, уровень гетероциклических аминокислот был достоверно ниже, чем в контрольной группе, не получавшей цеолиты. Содержание общего белка и ЛЖК в содержимом праксимальных отделов желудочно-кишечного тракта, по сравнению с контролем, уменьшилось, щелочная реакция кормового кома увеличилась, а концентрация спиртов -понизилась. Природные цеолиты одинаково угнетали бродильные процессы в дистальных отделах кишечника, о чём свидетельствовала более щелочная реакция содержимого, а также снижение в нём концентрации суммарных ЛЖК. Природные цеолиты увеличивают коэффициент переваримости протеина, липидов, повышают уровень большинства свободных аминокислот плазмы крови цыплят, за исключением глутамина, цистеина и цистина. Описанные сдвиги начинали проявляться с 6-10 дня с начала опыта и становились наиболее выраженными в 35-50 - дневном возрасте цыплят.
Н.М. Головина (1992) на гусятах гибридах итальянской и рейнской породы испытывала 5 доз цеолита от 1,7% до 7% к основному рациону. Добавка пегассина повысила в рационе содержание кальция и фосфора в 2-3,5 раза. В результате исследований было установлено, что в опытных группах количество отложенного в организме азота и конверсия его в белок были выше, чем в контроле на 9,6 и 5,7%, резервная щёлочность крови увеличилась на 13,0-23,0%, содержание витамина А в печени возросло в 1,2-1,4 раза, кроме того в печени наблюдалось повышение калия в 1,6 раза, железа в 2,7, марганца - в 1,54, меди - в 3,4, цинка - в 1,6 раза. В тушках гусят опытных групп белка в мышечной ткани было больше на 3,0% по сравнению с контрольной группой.
В.Т. Калюжнов и др. (1991,1992) считают, что основное кормосберегающее действие цеолитов обусловлено повышением переваримости и усвояемости питательных веществ корма. При добавке 6% цеолита к рациону переваримость органического вещества у кур повышается с 67,1 до 72,0 %, использование азота корма с 45,6 до 46,8%, а добавка 3% цеолита в кормосмесь бройлеров увеличивает переваримость органического вещества с 68,1 до 69,5, использование азота - с 38,7 до 46,8%. Однако, авторы не рекомендуют скармливать цеолит совместно с солями микроэлементов. В данном случае усвоение азота, фосфора и микроэлементов снижается на 8,2 - 21,2%, по сравнению с группами, получавшими один цеолит и одни только соли микроэлементов.
В остром опыте на курах-несушках, продолжавшемся 60 дней, Т. Григорьева и Г. Иванов (1997) установили, что добавка к рациону пермаита Чувашского месторождения не отразилась отрицательно на патологоанатомической картине вскрытия. Печень у кур опытной группы имела нормальный объём, однородный темно-коричневый цвет, упругую паренхиму, в то время как в контрольной группе она была увеличена в объёме, имела тупые закруглённые края, тестообразную консистенцию, легко разрывалась и имела желто-коричневый, серо-жёлтый, глинистый цвет. В крови кур, получавших цеолит, количество эритроцитов увеличилось на 6-12%, гемоглобин на 4-20%, лизоцимная активность возросла на 59%, по сравнению с контрольной группой. На основании вышесказанного авторы пришли к выводу, что пермаит является сорбентом продуктов обмена веществ при эндогенной и экзогенной интоксикации птицы. Аналогичные данные были получены в своих исследованиях С.Д. Назаровым (1998).
О.А. Донченко (1999) при скармливании курам- несушкам 4% сахаптина к основному рациону установил увеличение переваримости сырого протеина на 2,5%, жира- на 12,2, клетчатки- на 14, БЭВ- на 2,2 и азота- на 5,76%.
Комплексные исследования, проведённые лично А.М. Шадриным (2001) и с сотрудниками (1987, 1988), показали, что добавка пегассина в рацион кур-несушек увеличивает трансформацию азота в яйцо на 1,5% от принятого с кормом и на 7,0% - от переваренного. При сопоставлении результатов химического анализа цеолита по 12 элементам на «входе» и на «выходе» достоверные результаты были получены по фосфору, кальцию и калию. В пищеварительном тракте происходит обогащение цеолита фосфором, калием и обеднение кальцием. Ионный обмен в системе «цеолит-организм» направлен на формирование нужного организму птицы соотношение ионов.
Добавка к рациону бройлеров и кур-несушек хонгурина способствует повышению в организме переваримости жира на 9,2-11,2%, БЭВ- на 2,2%, пролонгируется действие витаминов и ферментов, обеспечивается равновесие ионного обмена макро- и микроэлементов, улучшается процесс пищеварения. При ежедневном вводе природных цеолитов в рационы цыплят яичного направления продуктивности, цыплят-бройлеров, кур-несушек у них в крови повышается уровень общего белка и глобулинов, щелочной резерв и кальций, усвоение каротина и синтез витаминов А и Bi. При кормлении птицы недоброкачественными (токсичными) кормами предупреждается падёж в 2,02 раза, вынужденный убой - в 1,65 раза.
По своему химическому составу цеолиты разных месторождений отличаются друг от друга. Даже в одном месторождении разная глубина пласта имеет своё отличие по химическому составу. По данным С.Н. Байкова (2000), в среднем в цеолитах содержится: окиси кремния - 65,3 %, окиси алюминия - 12,0, окиси железа - 1,9, окиси магния - 1,2, окиси кальция - 3,1, окиси калия - 1,2, окиси марганца - 0,04, прочие элементы - 12,0 %.
Название глауконита происходит от греческого «глаукос» - голубовато-зелёный. По решению Международного номенклатурного комитета глауконитом следует называть железистую диоктаэдрическую слюду, неразбухающую, с (Al, Fe3+) lv>oi, ( Fe3+,Al)1V 1,2, Fe3+> Al. Содержание основных компонентов обычно находится в пределах (%): КЮ - 6-8; MgO - 3-4; АЮз -5-10; РезОз - 15-22; SiCh - 47-50,5; №0 - 7- 9; FeO - 2 - 4; CaO - О -0,8;Na>2>O-0-0,5.
Микрозондовые анализы частично показывают резкую неоднородность состава даже в образцах, однородных под микроскопом, разница в содержании отдельных компонентов составляет: 3% KiO, 4 % -Ре>2>0з, 3 % -А1>2> О з, 5 % - SiCh.
Процесс глауконитизации - процесс эволюционный.
Глауконитовые месторождения по своему происхождению относятся к осадочным, в то время как цеолиты имеют вулканическое происхождение.
Самыми крупными месторождениями глауконитовых песков в России являются Кимовское с запасом 15954 тыс.м3, Ростовское - 150 млн.т. Концентрация глауконитовых зёрен доходит в них до 30-50% (Байков С.Н.,2000).
Большие залежи глауконитовых песков распространены на Атлантическом побережье США - в окрестностях Мэриленда, Нью-Джерси, Дилавера и имеют минеральный состав (%): глауконит -80, кварц - 10-15, обломки раковин фораменифер - 5-10, карбонаты и глинистые минералы -1-3.
По данным В.Н. Удачина (1997), в составе глауконитов выделяются две группы, различающиеся цветом, - светло-зелёные и тёмно-зелёные. Темно-зелёные глаукониты характеризуются повышенной плотностью, высокой степенью совершенства структуры с низким содержанием разбухающих межслоевых промежутков, повышенным содержанием калия, закисного железа, пониженным содержанием обменных катионов, низкотемпературной воды. Светло-зелёные разности глауконитов имеют неупорядоченную структуру с повышенным содержанием разбухающих слоев, пониженную ёмкость катионного обмена и содержания низкотемпературной воды.
Ёмкость катионного обмена глауконитов прямо пропорциональна содержанию в их структуре разбухающих слоев. Поглощение глауконитом катионов переходных элементов четвёртого периода происходит по механизму ионного обмена, сопровождающегося вытеснением ионов калия.
В отличие от цеолитов глаукониты имеют не каркасное, а слоистое строение. Часть внутримолекулярных сил не уравновешена взаимодействием с расположенными в полости одного такого слоя ионами химических элементов. Эти силы могут вступать во взаимодействие с ионами химических веществ, содержащимися в растворах или в воздухе. В результате они скапливаются на активных поверхностях пластиночек, составляющих общий кристалл. Площадь активной поверхности значительно увеличивается и в этом, по всей вероятности, основное различие биологического действия глауконита по сравнению с цеолитом.
При использовании глауконита в качестве кормовой добавки его оптимальная норма составляет в десять раз меньше, по сравнению с цеолитом (0,25-0,5% от сухого вещества).
Добавка оптимальной дозировки глауконита в рацион супоросных свиноматок в исследованиях Е.В.Иванова (2000, 2000, 2001) позволила в первые две трети супоросности повысить переваримость протеина на 6,0%, жира - на 2,0, клетчатки - на 5,1 и БЭВ - на 3,9 %, а в последнюю треть супоросности эти показатели были выше контрольной группы
соответственно на 3,6 %; 2,3 ; 5,5 и 4,0 %. В крови животных данной группы зарегистрированы изменения анаболического характера в показателях белкового и липидного обмена, повышение клеточного и гуморального иммунитета; в последнюю треть супоросности происходит увеличение количества Т- и В-лимфоцитов на 4,6 и 5,6 %, секретируемого лизоцима - на 3,6 мкг/л, а общего лизоцима — 17,4 %, циркулирующих иммунных комплексов- на 39,7 %.
Г.А. Джинджихадзе и др. (2001,2001) установлено, что дозировка глауконита 0,25% от сухого вещества рациона свиньи на откорме повышает переваримость протеина на 4,3%, сырого жира - на!9,7 %, а среднесуточное отложение азота в теле было на 2,2 г больше. При этом в крови животных количество общего белка увеличилось на 4,3 -8,3 %, аминного азота - на 8,3-22,2 %, общих липидов на 22,9 - 26,1 %, (3-липоротеидов - на 10,9- 37,4 %, а мочевины уменьшилось на 5,2- 7,6 %. Кроме этого, в крови свиней наблюдалось повышение уровня основных биогенных микроэлементов ( кобальта, железа, цинка, марганца).
В исследованиях А.А.Замятина (2000) скармливание глауконита курам-несушкам в дозировках 0,15 , 0,25 и 0,5 % от сухого вещества рациона повысило переваримость органического вещества на 2,0 - 3,4%, сырого протеина - на 7,6- 7,8, жира - на 18,9 -19,9, клетчатки - на 10,8-10,9 и БЭВ -на 0,1 %. Использование азота корма у кур увеличивается на 7,9 - 8,9 %, кальция - на 9,1 -11,8 и фосфора - на 0,3 - 0,6% от принятого.
Следовательно, природные алюмосиликаты за счёт своих уникальных свойств вызывают в живом организме изменения, приводящие к сдвигу обмена веществ с преобладанием процессов анаболизма, что в конечном итоге сказывается на продуктивности и экономических показателях отросли.
1.4 Изменение продуктивности птицы под влиянием кормовой добавки цеолитов
Анализ доступной отечественной и зарубежной литературы показывает, что научных работ по исследованию цеолитов в качестве кормовой добавки в рационах сельскохозяйственной птицы имеется в достаточном количестве. Но единого мнения в отношении дозировок цеолитов нет. Очевидно, это зависит от типа цеолита, его содержания в породе и наличия других примесей (В. Калюжнов и др. , 1991). Хотя научные исследования, выполненные в разных странах, подтвердили положительный эффект от добавления цеолитов в рационы как молодняка, так и взрослой птицы. Введение 3-6% цеолитов в комбикорма не отражается на его потреблении, увеличение же дозы до7- 10% снижает поедаемость корма птицей и отрицательно сказывается на её продуктивности (Ленкова Т., Синцерова О., 1985).
По данным Т. Григорьевой и Г. Иванова (1997), включение в рацион кур - несушек Чувашского трепела в количестве 1,2 и 3% яйценоскость кур за 60 дней опыта увеличилась соответственно на 5,18 и 19 %.Сохранность поголовья возросла с 93,1 до 98,6 %. При этом оптимальной дозировкой было 3% цеолита. А при включении в кормосмесь кур-несушек 8-21 недельного возраста Дзегвского цеолита в дозе 3,5 и 7% В. Великанов и др. (1983) получили такую же сохранность поголовья - 98,8%.
Аналогичные результаты приводит в своём аналитическом обзоре В. Фисинин с соавторами (1990). В частности, клиноптилолиты Дзегвского и Тедзамского месторождений в кормлении молодняка кур яичных линий повышают жизнеспособность птицы на 1-2%, темпы роста - на 4-12 %, расход корма на единицу продукции уменьшается на 4-16 %. При этом оптимальная норма ввода считается 3 и 5% в рационе. Но с повышением дозировки с 5 до 7% яйценоскость кур снижается на 1,8%. Испытание гейлатдина пегасского месторождения в количестве 3-6% к рациону увеличивает яйценоскость птицы на 1,5-3,0%, сохранность поголовья на 3,3-5,0%, снижает расход корма в расчёте на 10 яиц на 4,6-8,3%. Включение 5% пегассина в комбикорм гусят-бройлеров увеличивает живую массу птицы на 5,1%, снижает затраты корма на 1 ц прироста на 7,3%.
Широкая производственная апробация, проведённая И.А. Чонка (1984) на 5459 головах кур-несушек по скармливанию по 50 и по 100 г на 1кг корма цеолита Сокирницкого месторождения, показала, что яйценоскость кур повысилась на 5 - 7,5%, прочность скорлупы яйца - на 65-82%, сохранность поголовья - на 0,4%. Лучшая доза оказалась 50 г на 1кг комбикорма. При этом экономический эффект составил 735,4 тыс. руб. в год.
При добавлении в корм бройлерам Закарпатских туфов С.А. Водолажченко и др. (1980) установили снижение расхода корма на 1 кг прироста при дозе 4% - на 4,2, а при 6,0-8% - на 8,3%. При этом не установлено вредного действия цеолитов на жизнеспособность птицы и качество получаемой продукции. Добавка этих минералов в рацион кур -несушек оказала положительное влияние на сохранность, прирост живой массы, яйценоскость и выводимость (Мерабишвили М.С. и др., 1980).
По данным F.Mumpton и P. Fishman (1974), введение цеолита в корм птицы повышает интенсивность её роста, улучшает конверсию корма, жизнеспособность, снижает потребление корма и воды.
Н. Квашали и др. (1980), N. Kvashali et. al.(1980) на цыплятах яичных линий и бройлеров испытывался вариант уменьшения весовой части рациона за счёт ввода в его состав клиноптилолита и вариант, когда клиноптилолит являлся составной частью рациона и входил в его состав как отдельный ингредиент. С практической точки зрения заслуживает внимания первый вариант, так как в этом случае без ухудшения зоогигиенических показателей достигается удешевление и экономия кормов.
В исследованиях Н.В. Мухиной (1991,1992,1993) добавка 5% пегассина в рацион ремонтного молодняка кур повышает сохранность
поголовья на 2,1 %, расход комбикорма уменьшается на 8%. Использование цеолита позволило сохранить расход ракушки на 0,5% и полностью исключить из рациона гравий.
Положительные результаты были получены Г.А. Романовым (1993) при скармливании шивыртуина гусятам-бройлерам. В данной группе сохранность поголовья была самой высокой и составила 90,2-92,7%, в то время как в группе, получавшей пегассин, она равнялась 83,7%, а в группе, где цеолиты не применялись, - 81,3%. При этом расход кормов у гусей по группам соответственно был 3,20; 3,67 и 3,88 кг.
A.M. Шадриным (2000,2001), A.M. Шадриным и Г.А. Жуковым (2000), A.M. Шадриным и др. (1987) рекомендуются оптимальные нормы ввода для кур-несушек сахаптина 4%, пегассина 4-6, хонгурина-6%. При этом яйценоскость птицы повышается на 7,5 %, сохранность поголовья - на 2,3-5,6 %, расход корма снижается на 17-21%. Для цыплят - бройлеров оптимальной дозой считается 6% хонгурина. При этом среднесуточный прирост увеличивается на 6,5 %, а затраты снижаются на 13%. Для ремонтного молодняка рекомендуется включать в рацион 6% пегассина, это способствует интенсивности роста птицы на 5-8%, повышает её сохранность на 5%, одновременно сокращая затраты кормов на 5%. Природные цеолиты снижают заболеваемость и падёж птицы на 1,7-3,7%.
А.В. Якимовым с коллегами (1999,1997) при скармливании бройлерам цеолита Марийской ССР шатрашанита в количестве 2,0 и 4,0 % от массы комбикорма увеличили прирост их живой массы соответственно на 4,2 и 7,1%. Увеличение нормы ввода цеолита до 6%, наоборот, способствовало снижению их живой массы.
О.А. Донченко (1999, 2002), скармливая курам-несушкам 4% к массе комбикорма сахаптина, увеличил яйценоскость птицы на 3,31%, массу яйца -на 0,48 г, затраты кормов в расчёте на 1000 яиц уменьшились на 7,55%. У цыплят яичного направления продуктивности аналогичная дозировка
цеолита способствовала повышению прироста живой массы на 4,83%, их сохранности на 2,6 %, снижению затрат корма на 1кг прироста на 15,4%. Полученный экономический эффект составил 28,8 руб. на 1 руб. скормленного цеолита.
В исследованиях Г. Иванова и др. (1997) при добавлении в рацион кур - несушек 3% пермаита их продуктивность удалось повысить на 13%, сохранность поголовья - на 5,5%, а выбраковку птицы уменьшить на 5,5%.
С.Н. Касумов и др. (1987) рекомендуют клиноптилолит Айдагского месторождения Азербайджана вводить в состав рациона кур-несушек, цыплят - бройлеров, индюшат и ремонтного молодняка в количестве 5%, взрослой птице -3%. При этом экономический эффект составляет 460 руб. в расчёте на 1000 несушек.
В то же время С.А. Водолажченко и др. (1984) считают, что клиноптилолит с величиной частиц менее 2мм можно без ущерба для продуктивности использовать в кормлении кур - несушек в количестве 6-12% от массы комбикорма. За 270 дней яйценоскости добавка 4 и 8% цеолита снизила яйценоскость на курицу-несушку с 126,8 шт. в контроле до 122,5 и 113,8 штук в опытных группах. Однако, оплодотворяемость яиц в опытных группах была выше на 1,3-1,7 %, прочность скорлупы повысилась с 2,357 г/мм до 2,523 г/мм в опытных группах. Полученные данные совпадают с результатами В. Фисинина и др. (1985).
По данным американских исследователей (Mupton F., 1978), включение в рацион цыплят 3,0; 5,0 и 8,0 % клиноптилолита увеличивает эффективность использования корма и на 25% уменьшает содержание воды в помёте.
В опытах Н.Ф. Квашали и З.Г. Микаутадзе (1980) на цыплятах при включении в их рацион 5% цеолита наблюдается увеличение их живой массы на 3,4 %, уменьшение расхода кормов на 4,4%, повышения сохранности поголовья на 2,9 %. При этом экономия кормов составляет 3475 кг,
себестоимость продукции снижается на 1,5 %, а экономический эффект в расчёте на 1000 голов составляет 211,6 руб.
Цеолиты, помимо того, что могут использоваться в рационах сельскохозяйственной птицы в чистом виде, также могут служить основой для производства биологически активных добавок. Так, А.Р. Мацерушка (1997) предлагает приготовлять яично-цеолитовую муку (25-30 кг яичной массы на 100 кг цеолита) и кровяно-цеолитовую муку (25-30 кг цеолита на 100 кг крови). При влажности 8-12 % их включают в количестве 6-9 % в замен комбикорма в рацион цыплят -бройлеров. При этом переваримость клетчатки увеличивается на 2,3-5,3 %, БЭВ - на 1,6 и 2,2%, усвоение азота повышается на 4,3- 9,3%. Бройлеры, получавшие такую подкормку, имели выше живую массу на 2,9-6,4 %, убойный выход - на 2,2 - 2,6 %, индекс мясности- на 11,1-4,8 %, бедра -на 11,8 - 11,6 %, содержание жира в фарше -на 25,2 и 20,4%.
Н.Р. Бисикенов (1993) предлагает использовать цеолит с витаминными премиксами, применение которых повышает сохранность цыплят на 2-3 %, уменьшает затраты корма на единицу продукции на 3,1 -6,7 % и способствует увеличению живой массы цыплят на 1,1- 2,1 %.
Л.С. Кудряшовым и др. (1998) из всех изучаемых дозировок при добавлении в рацион цыплят- бройлеров 8% пегассина увеличение убойного выхода тушек не превышало 1,8 %, в то время как добавка 6% того же цеолита увеличила коэффициент мясности с 2,3 до 2,7.
А.В. Якимов и др. (1996) при добавлении к рациону цыплят -бройлеров 2,4 и 6% цеолита шатрашанах позволили получить среднесуточный прирост соответственно на 4,0 , 7,2 и 2,4 % выше чем в контрольной группе. Авторы считают оптимальную дозу 4% от массы комбикорма.
В исследованиях Н.Е. Тен и др. (1992) включение 5% пегассина в рацион кур-несушек повысило их сохранность на 2,6%, способствовало увеличению живой массы на 10,8 , яичной продуктивности - на 7,0, интенсивность яйцекладки - на 4,1 , выход инкубационных яиц - на 2,1 и инкубационные качества яйца - на 5,5 %. При этом затраты корма на производство яйца снизились на 3,7 %, а себестоимость - на 12,3 %. В исследованиях А.А. Замятина (2000) кормовая добавка глауконита в рационах кур увеличила их продуктивность на 6,5 - 13,3%, массу яйца на 2,1 -5,3 , толщину скорлупы на 0,9 - 2,6 и уменьшило её деформацию на 3,7 -14,2 %. Подводя итог, следует сказать, что оптимальные дозировки цеолитов в рационе сельскохозяйственной птицы оказывают положительное влияние на её продуктивность, рост, развитие и экономически выгодно.
1.5 Заключение по обзору литературы
Развитию отечественного птицеводства в настоящее время уделяется должное внимание, так как это одна из рентабельных отраслей и первая перешла на промышленную основу развития.
Из всех видов сельскохозяйственной птицы гусеводству по его хозяйственно-полезным признакам уделяется должное внимание. Особенно заслуживает внимания вопрос выращивания гусят-бройлеров, у которых в 60-70 дневном возрасте можно получить живую массу от 3,5 до 4,5 кг.
При организации мясного откорма молодняка гусей важными факторами являются правильно выбранная порода, способ и условия их содержания, кормление полноценными и сбалансированными комбикормами. Из 20 пород и породных групп гусей, разводимых в нашей стране, большинство учёных (Салеев П., Ионова Е.И., Шонина С. и др.) рекомендуют для выращивания гусят-бройлеров использовать помесную (двух-трёхпородную) птицу, хорошо адаптированную к местным природно-климатическим условиям. Причём в первые три недели выращивания оптимальным считается содержание в комбикорме 20% сырого протеина, а в последующие дни - 18%. Концентрация обменной энергии в течение всего периода выращивания не должна быть ниже 280 ккал на 1% сырого протеина.
Из всех применяемых как в нашей стране, так и за рубежом, способов содержания гусят наиболее технологичным с физиологической точки зрения многих учёных (Покровский В.А., Салеев П., Ионовой Е.И., Bartels К., Dayen М.и Fiedler H.) является выращивание гусят-бройлеров в помещениях на глубокой подстилке со свободным доступом к водоёму. Это способствует более быстрому росту пухо-перьевого покрова и физиологическому развитию птицы.
Скорейшего роста и развития птицы можно добиться, используя в их рационах в качестве кормовой добавки различных природных алюмосиликатов-цеолитов. Высокая их ионообменная способность в живом организме позволяет восполнить дефицитные макро- и микроэлементы рациона, улучшить их усвоение и направить обмен веществ в организме в анаболическом направлении. Это явление характерно практически для всех цеолитов, залежи которых находятся на территории России, стран ближнего и дальнего зарубежья. При этом важным является вопрос о правильности нормы ввода цеолита в рацион сельскохозяйственной птицы.
Научно-обоснованной нормой ввода цеолитов в рационы различных производственных групп и видов птицы, по данным A.M. Шадрина, В. Фисинина, О.А. Донченко, Г.М. Цицишвили, F. Mumpton и других, считается от 3 до 6% от массы комбикорма. Это способствует повышению переваримости органического вещества на 4,2%, сырого протеина - на 4,3 жира - на 11,1%, а усвоение азота - на 15,5%. При этом в печени птиць содержание витамина А увеличивается в 2 с лишним раза, каратиноидов - в четыре раза, в яйце количество витамина Е возрастает в 3 раза. Добавка цеолитов повышает в организме птицы окислительно-восстановительные процессы.
По своим физико-химическим характеристикам глауконит, как и цеолиты, относится к алюмосиликатам. Но его слоистое строение в отличие от цеолитов, позволяет увеличить активную поверхность в пять и более раз. В результате чего ионообменные свойства глауконита наиболее выше по сравнению с цеолитами. В проведённых исследованиях Е.В. Иванова, Г.А. Джинджихадзе оптимальная дозировка глауконита в рационах свиней была на порядок ниже, чем для цеолитов. Впоследствии это было подтверждено в исследованиях А.А. Замятина на курах-несушках. Добавка 0,25% глауконита в рацион кур-несушек увеличила переваримость органического вещества на 2,0 - 3,4 %, сырого протеина - на 7,8, жира - на 18,9-19,9, клетчатки - на 10,8 — 10,9 %. При этом лучше использовался азот, кальций и фосфор рациона. В результате чего продуктивность кур возросла на 6,5 - 13,3 %, масса яйца - на 2,1 - 5,3, толщина скорлупы - на 0,9 - 2,6 %. Аналогичные результаты отмечены многими исследователями при скармливании цеолитов.
Однако как в отечественной, так и в зарубежной литературе имеются лишь отдельные научные данные по использованию в качестве кормовой добавки в рационах гусей природных алюмосиликатов. Поэтому основной целью наших научных исследований являлось изучить возможность использования глауконита в качестве кормовой добавки в рационах гусят-бройлеров.
2. СОБСТВЕННЫЕ ИССЛЕДОВАНИЯ
2.1 Методика и условия проведения исследований
2.1.1 Химический состав используемого глауконита
Изучение химического состава глауконита Карийского месторождения Челябинской области проводил институт минерологии Уральского отделения Академии наук, находящийся в городе Миассе Челябинской области (Удачин В.Н., 1997).
Химический состав природных алюмосиликатов меняется в зависимости от местонахождения и глубины залегания пласта. В таблице 1 приводятся средние значения содержания отдельных микроэлементов в глауконите Карийского месторождения.
Данные минерального состава свидетельствуют, что содержание основных микроэлементов в глауконите находится ниже предельных допустимых концентраций для данного сырья.
1. Содержание отдельных микроэлементов в глауконите, мг/кг
|
Микроэлемент |
Глауконит |
пдк |
|
Медь |
5,9 |
55,0 |
|
Цинк |
37,0 |
100,0 |
|
Кобальт |
17,3 |
50,0 |
|
Железо |
- |
- |
|
Марганец |
21,0 |
1500,0 |
|
Свинец |
10,2 |
32,0 |
|
Никель |
15,5 |
85,0 |
|
Кадмий |
0,98 |
3,0 |
|
Ртуть |
0,04 |
2,0 |
|
Мышьяк |
3,8 |
5,0 |
Следует отметить, что в очищенном концентрате Карийского месторождения собственное содержание глауконита составило 94 %. Ионообменная ёмкость глауконита составляет 0,4 мг-экв/г, что ниже по сравнению с цеолитами (0,7 мг-экв/г), но слоистое строение глауконита позволяет увеличить количество разбухающих слоев от 5 до 55% при рН 8 и более.
2.1.2 Методика, схема и техника проведения исследований
Для решения поставленных задач нами в период с апреля по август 2001 и 2002 годов на гусеводческой ферме АОЗТ «Птицевод» Октябрьского района Челябинской области был проведён научный, научно-хозяйственный и производственный эксперимент на помесных гусятах шадринской и итальянской пород. Группы, на научный и научно-хозяйственный опыт, были подобраны по принципу аналогов; с учётом живой массы, породы, возраста и состояния здоровья.
В каждой группе в физиологическом опыте было по 5 голов гусят-бройлеров, в научно-хозяйственном - по 500, в производственном: в контрольной 950, а в опытной 975 голов.
В период проведения исследований хозяйство было благополучно по инфекционным и инвазионным заболеваниям. В зоогигиеническую оценку помещения в первые две недели входило: учёт плотности посадки птицы, освещённость, продолжительность светового дня, исследование температуры и относительной влажности воздуха психрометром Августа, аммиака — универсальным газоанализатором УГ-2, освещённость - люксметром Ю- Ю-116 (Кузнецов А.Ф. и др., 1999).
Поедаемость корма - путём ежедневного учёта дачи и остатков комбикорма. Химический состав и питательность комбикорма проводили в межкафедральной лаборатории УГАВМ по общепринятым методикам (Лебедев Л.Т., Усович А.Т.,1976, Малахов А.Г. и др., 1994, Петухова Е.А и др., 1989): первоначальную воду - методом высушивания навески корма в сушильном шкафу при температуре 65° С до постоянной массы; гигроскопическую воду - методом высушивания навески корма в сушильном шкафу при температуре 100 - 105° С до постоянной массы; «сырую» золу -сжиганием навески корма в муфельной печи при температуре 500-600° С; «сырой» протеин - по методу Кьельдаля; «сырую» клетчатку - кипячением в слабых растворах кислот и щелочей по методу Геннеберга и Штомана; «сырой» жир - экстракцией сернокислым эфиром в аппаратах Сокслета; кальций - трилонометрическим методом (Холод В.М., Ермолаев Г.Ф., 1988); фосфор - методом колориметрии (Холод В.М., Ермолаев Г.Ф., 1988). Содержание аминокислот взято из данных удостоверения качества комбикормов ПК - 4, завозимых на гусиную ферму с Магнитогорского комбината хлебопродуктов.
Микроэлементы Си, Fe, Zn, Co, Mn, а также макроэлемент Mg, определяли на атомно-адсорбционном спектрофотометре (Сырье и продукты пищевые, 1997).
Калорийность рационов и расчет использования энергии корма гусятами рассчитывали косвенным методом по методике ВИЖ (Щеглов В.В. и др., 1991).
2.1.3 Методика проведения балансового опыта
Для изучения влияния различных дозировок глауконита на переваримость и использование питательных веществ кормосмеси был проведён балансовый опыт в конце периода откорма гусят, т.е. в 50-дневном возрасте на 5 аналогичных по массе бройлерах из каждой группы по методике ВНИТИП и по методу М.И. Дьякова. Продолжительность балансового опыта 13 дней, из которых 5 дней учётных.
Птица содержалась в отдельных клетках с сетчатым дном, под которым установлены каркасы из полиэтиленовой плёнки для сбора помёта.
В течение балансового опыта ежедневно учитывали количество съеденного корма, путём учёта остатков корма от заданного и количество выделенного помёта. Помёт собирали дважды в день (утром и вечером), взвешивали, помещали в двойные полиэтиленовые пакеты (тщательно закрываемые), заливали 0,1 н раствором щавелевой кислоты (2 мл на 50 г помёта) для связывания аммиака. Количество пошедшей кислоты учитывали при определении первоначальной воды. Помёт хранили в холодильнике на нижней полке. Химический состав кормосмеси, помёта проводили в межкафедральной лаборатории УГАВМ по общепринятым методикам, указанным выше. Азот кала определяли по методу М.И. Дьякова (Маелиев И.Т, 1968).
Коэффициенты переваримости, балансы азота, кальция и фосфора вычисляли по общепринятым методикам (Томмэ М.Ф., 1969).
В конце балансового опыта был проведен контрольный убой 5 гусят-бройлеров из каждой группы. При этом собранная для исследования кровь была доставлена в отдел биохимического анализа межкафедральной лаборатории УГАВМ.
2.1.4 Методики лабораторных исследований
В межкафедральной лаборатории УГАВМ в цельной крови определяли:
- гемоглобин - гемоглобинцианидным методом при помощи набора химических реактивов для определения массовой концентрации гемоглобина крови (Пименова М.Л., Дервиз Г.В., 1974). Гемоглобин при взаимодействии с железосинеродистым калием окисляется в метгемоглобин, образующий с ацетонциангидридом окрашенный гемоглобинцианид, интенсивность окраски которого пропорциональна содержанию гемоглобина;
- подсчёт эритроцитов и лейкоцитов — проводили в камере Горяева (Кондрахин И.П. и др., 1985) путём подсчёта клеток белой и красной крови соответственно в 5-ти больших квадратах и 5-ти полосах.
Из биохимических показателей в сыворотке крови по общепринятым методикам определяли:
общий белок - рефрактометрическим методом на рефрактометре типа «RL-2», в основу которого положено определение показателя преломления исследуемого вещества. В сыворотке крови величина рефракции в первую очередь зависит от количества белка (Кондрахин И.П. и др., 1985);
холистерин - при помощи набора «БИО-ЛА-ТЕСТ» (Антонов Б.И. и др., 1991). Холестерин в присутствии уксусного ангидрида и смеси уксусной и серной кислот даёт изумрудно-зелёное окрашивание, интенсивность которого прямо пропорциональна его концентрации; общие липиды - фотоколориметрическим методом по Бурштейну. В основе метода лежит реакция избирательного осаждения бета-липопротеидов гепарином в присутствии двухвалентных катионов (Дрозденко Н.П. и др., 1981);
(3 - липопротеиды - фотоколориметрическим методом по Бурштейну. В основе метода лежит реакция избирательного осаждения бета-липопротеидов гепарином в присутствии двухвалентных катионов (Дрозденко Н.П. и др., 1981);
аминный азот - по реакции с нингидрином. Аминокислоты при взаимодействии с нингидрином подвергаются окислению, при этом образуется соединение, окрашенное в фиолетовый цвет, интенсивность окрашивания при определённых условиях пропорциональна количеству свободных аминокислот (Кондрахин И.П. и др., 1985); кальций - трилонометрическим методом с индикатором флюорексоном по Вичеву и Каракашеву (Холод В.М., Ермолаев Г.Ф., 1988). Метод основан на различной прочности комплексных соединений, образуемых кальцием, флуорексоном и трилоном Б;
фосфор - определение в безбелковом фильтрате крови с ванад-молибденовым реактивом (по Пулсу в модификации В. Ф. Коромыслова и Л. А. Кудрявцевой). Метод основан на том, что фосфор в безбелковом фильтрате дает лимонно-желтое окрашивание (Холод В.М., Ермолаев Г.Ф., 1988).
Микроэлементы крови - определяли методом атомно-адсорбционной спектрофотометрии (Сырье и продукты пищевые, 1997).
Для исследования минерального состава костяка брали левую и правую большеберцовые кости у 5-ти бройлеров из подопытных групп, в которых определяли золу, кальций, фосфор, микроэлементы по описанным выше методикам.
Мясную продуктивность определяли в конце опыта путем проведения контрольных убоев 5-ти гусят-бройлеров из каждой группы по методике ВНИТИП (Маслиева О.И.,1970 и Матрозова С.И.,1977).
При этом определяли предубойную, убойную массу, массу тушки, массу съедобных и несъедобных частей, внутреннего жира, химический состав мяса.
Калорийность мяса определяли расчетным путем по химическому составу и калорическим коэффициентам: 1 г жира = 9,3 ккал, 1 г белка = 4,1 ккал. Энергетическая ценность мяса (кДж) рассчитали исходя из того, что 1ккал соответствует 4,186 кДж.
Расчет конверсии протеина и энергии корма в продукцию проводили по методике Лепайые Л.К. и др. (1983).
Результаты опытов обрабатывались биометрически на микрокалькуляторе и персональном компьютере с программированным обеспечением. Достоверностью считали разницу при Р<0,05.
2.1.5 Содержание и кормление гусят-бройлеров
В АОЗТ «Птицевод» Октябрьского района Челябинской области, на базе которого проводились исследования, всё поголовье гусей с суточного возраста выращивается на глубокой подстилке.
Птичник, в котором находились контрольная и опытные группы, представляет собой приспособленное помещение размером 1800 кв.м с центральным проходом. Всё помещение было разделено на восемь секций по 250 гусят в каждой, и каждая секция имела свой выход на выгульный дворик. Плотность посадки гусят в первую половину выращивания составляла 10 голов на 1 кв. м, в последние 30 дней — 5 голов. Перед заполнением птичника гусятами его бетонный пол был посыпан гашёной известью, сверху которой был положен слой измельчённой соломы из расчёта 0,8 кг на кв.м. В течение первых двух недель безвыгульного содержания гусят сухой подстилочный материал добавлялся через день. В результате чего к 14 дню его толщина составила 12 см. Вдоль всего птичника с обеих сторон от центрального прохода имелись продольные поилки, которые заполнялись водой из водопроводной сети через поплавковую камеру, которая соединяется шлангом с подводящей линией. Кормление гусят осуществлялось посредством его подачи из бункера накопителя через шнековый механизм по закрытым кормопроводам в бункерные кормушки, к которым гусята имели свободный доступ. Температура в помещении и относительная влажность, контролировалась с помощью психрометра Августа. За счёт круглосуточной автоматической работы теплогенератора ТГ-25. В первые семь дней температура в помещении поддерживалась на уровне 30 С°, в последующие семь дней - 26С°. Относительная влажность воздуха была на уровне 75%. Закладка яиц на инкубацию и вывод первой партии гусят в АОЗТ «Птицевод» приурочивают к тому времени, когда после двухнедельного содержания в помещении гусят можно постепенно, с 30-40 минут, приучать к прогулкам на солнце. Выгульная площадка птичника, в котором проводился эксперимент, была разделена мелкоячеистой сеткой на число секций птичника. Каждая маленькая выгульная площадка имела огороженный доступ к водной поверхности озера, по периметру которого расположены производственные помещения Акционерного общества.
С возрастом гусята большую часть времени светового дня находились у водоёма, а кормление их проводилось в помещении.
Таким образом, для всей подопытной птицы в эксперименте были созданы одинаковые условия содержания и ухода по принятому в хозяйстве распорядку дня (таблица 2).
2. Распорядок дня при выращивании гусят на мясо в АОЗТ «Птицевод»
|
№ п/п |
Наименование работы |
Начало, час-мин. |
Окончание, час-мин. |
|
1. |
Подготовка к работе |
8-00 |
8-30 |
|
2. |
Кормление, осмотр поголовья, сбор павшей птицы |
8-30 |
9-00 |
|
3. |
Сдача падежа |
9-00 |
9-30 |
|
4. |
Выгон гусей на выгулы |
9-30 |
10-00 |
|
5. |
Техобслуживание оборудования, уборка помещений |
10-00 |
12-00 |
|
6. |
Перерыв на обед |
12-00 |
14-00 |
|
7. |
Техобслуживание оборудования, осмотр птицы |
14-00 |
14-30 |
|
8. |
Загон гусей в помещение |
14-30 |
15-30 |
|
9. |
Кормление и выполнение разовых рабог |
15-30 |
17-30 |
|
10. |
Заключительные работы и уборка рабочего места |
17-30 |
18-00 |
В течение всего опыта кормление подопытной птицы проводилось в соответствии с нормами ВНИТИП с учётом возраста.
В течение опыта кормление гусят осуществляли по двум периодам: 1-20 день и 21-66 дней. Полнорационный комбикорм, который получали все подопытные гусята, был приготовлен путём добавления к зерновой части БВМД в количестве 20% от общей массы комбикорма.
Состав комбикорма, применяемого при кормлении подопытных гусят, представлен в приложении (табл. 28).
Среднее потребление комбикорма и питательных веществ гусятами в течение научно-хозяйственного опыта представлено в таблице 3. Состав и питательность комбикорма в период выращивания гусят в первые двадцать дней позволили в 100 г комбикорма иметь 311,0 ккал, или 1,30 МДж обменной энергии, 21,6 г сырого протеина. В результате чего энергопротеиновое отношение составило 144,5 ккал в расчёте на 1% сырого протеина, что соответствует научно-обоснованным нормам, так как при узком энергопротеиновом отношении избыток протеина будет расходоваться на энергетические цели, а при широком - снижается продуктивность птицы.
Жир необходим птице, как энергетический материал, так и среда для растворения жирорастворимых витаминов. В первый период выращивания его концентрация составила 5,1 % в 100 г комбикорма. Основная роль сырой клетчатки в организме не только сельскохозяйственной птицы, но и животных заключается в формировании каловых масс. От её количества во многом зависит заполнение кормовыми массами желудочно-кишечного тракта. Для молодняка птицы её уровень намного ниже по сравнению с взрослой. Поэтому в первый период выращивания гусят её количество находилось на уровне 4,2%.
3. Потребление корма и питательных веществ гусятами за период опыта (в среднем на голову в сутки)
|
Показатель |
Возраст, дн. |
|
|
1-20 |
21-66 |
|
|
Полнорационный комбикорм, г |
65,5 |
276,0 |
|
В кормосмеси содержится : |
||
|
обменной энергии: ккал МДж |
203,7 0,85 |
831,0 3,48 |
|
сырого протеина, г |
14,1 |
51,3 |
|
сырой клетчатки, г |
2,8 |
17,0 |
|
сырого жира, г |
3,3 |
12,4 |
|
лизина, мг |
786 |
2650 |
|
метионина + цистина, мг |
590 |
1932 |
|
кальция, мг |
655 |
2760 |
|
фосфора, мг |
459 |
1794 |
|
натрия, мг |
65,5 |
276 |
|
меди, мг |
0,33 |
2,26 |
|
железо, мг |
1,97 |
4,86 |
|
цинка, мг |
3,28 |
12,4 |
|
кобальта, мг |
0,06 |
0,80 |
|
марганца, мг |
6,55 |
11,59 |
|
линолевой кислоты, г |
1,51 |
5,24 |
|
ЭПО, ккал на 1 % сырого протеина |
144,5 |
162,0 |
Содержание основных нормируемых для сельскохозяйственной птицы аминокислот характеризует биологическую полноценность протеина рациона. К основным аминокислотам относятся лизин и метионин с цистином, количество которых было на уровне соответственно 1,2 и 0,9 мг в 100 г комбикорма.
Для птицы, в отличие от сельскохозяйственных животных, одним из нормируемых показателей является линолевая кислота. Она способствует повышению усвояемости питательных веществ рациона, улучшению ожиренности и сортности тушки. В комбикорме первого периода выращивания её содержание составило 2,3 %.
Во второй период выращивания гусят (21-66 дней) концентрация вышеперечисленных питательных веществ в 100 г комбикорма составила 301,1 ккал, или 1,26 МДж обменной энергии, 18,6 г сырого протеина, 4,5 г сырого жира, 6,15 г сырой клетчатки, 1,9 г линолевой кислоты, 0,96 мг 1 лизина и 0,70 мг метионина с цистином. Энерго-протеиновое отношение в этот период было на уровне 162 ккал в расчёте на 1% сырого протеина. В комбикорме первого и второго периодов выращивания, содержание кальция и фосфора соответственно составило 1,0 и 0,7 %, 1,32 и 0,65 %, натрия- 0,1%. Количество основных нормируемых микроэлементов соответствовало норме кормления.
Жиро — и водорастворимые витамины восполнялись в рационе гусят за счёт витаминно-минерального премикса, норма ввода которого составляла 0,5 и 1,0% от общей массы комбикорма.
На фоне такого основного рациона кормления, который получала птица всех групп, гусятам II - IV опытных групп добавлялся глауконитовый концентрат в количестве соответствующим схеме опыта. Фактическое потребление глауконита гусятами в зависимости от съеденного комбикорма представлено в таблице 4.
4. Потребление корма и глауконита гусятами за период опыта (в среднем на голову в сутки)
|
Скормлено по группам глауконита, мг |
|||||
|
Возраст, |
Потреблено |
||||
|
дней |
комбикорма, |
||||
|
г |
I |
II |
III |
IV |
|
|
1-10 |
35 |
- |
30 |
60 |
90 |
|
11-17 |
90 |
- |
76 |
152 |
228 |
|
18- 24 |
110 |
- |
93 |
186 |
279 |
|
25-31 |
221 |
- |
187 |
374 |
561 |
|
32-38 |
270 |
- |
229 |
458 |
687 |
|
39-45 |
280 |
- |
238 |
476 |
714 |
|
46-52 |
328 |
- |
279 |
558 |
837 |
|
53-59 |
338 |
- |
287 |
574 |
861 |
|
60-66 |
320 |
- |
272 |
544 |
816 |
|
ИТОГО: |
14049 |
- |
11777 |
23554 |
35331 |
Глауконит гусятам опытных групп задавался ежедневно при утреннем кормлении в зависимости от их количества в группе путём равномерного смешивания его с комбикормом.
В результате чего за весь период откорма в расчёте на одного гусёнка было затрачено 14,049 кг комбикорма, а количество скормленного глауконита составило: во II группе 11,777 г, в III - 23,554 и в IV группе -35,331г.
Добавка глауконитового концентрата в рацион гусят опытных групп оказала определённое влияние на их рост и развитие.
3. РЕЗУЛЬТАТЫ СОБСТВЕННЫХ ИССЛЕДОВАНИЙ
3.1 Динамика живой массы и сохранность гусят-бройлеров за период откорма
Сложный процесс индивидуального развития организма представляет совокупность количественных и качественных изменений, происходящих после акта оплодотворения яйцеклетки и образования зиготы, на протяжении всей жизни особи, в соответствии с унаследованным генотипом и нормой реакции.
Ещё Ч. Дарвин представлял индивидуальное развитие организма как совокупность двух процессов: роста и развития. «Всякий организм, - писал он (цитата по Кравченко Н.А.,1973)- достигает зрелости после более или менее длительного периода роста и развития: первый из этих терминов означает только увеличение размеров, термин же «развитие»- изменение строения».
Использование глауконитового концентрата в рационах гусят-бройлеров на фоне основного рациона кормления позволило получить данные, представленные в таблице 5.
5.Динамика живой массы гусят, г (X ±т\, п =50).
|
Возраст, дней |
Группа |
|||
|
I |
II |
III |
IV |
|
|
1 |
95,9±0,16 |
96,1±0,15 |
96,2±0,16 |
96,0±0,14 |
|
11 |
381,9±3,23 |
416,1±2,99*** |
393,2±3,15** |
410,0±4,05*** |
|
21 |
812,9±9,10 |
873,1±15,12*** |
875,2±13,91*** |
877,5±14,99*** |
|
31 |
1379,9 ±14,77 |
1468,1±12,80*** |
1539,2±16,45*** |
1514,0±17,12*** |
|
41 |
201 1,9 ±42,51 |
2133, 1± 39,90* |
2272,2± 40,70*** |
2186,0±44,13** |
|
51 |
2762,9 ±17,02 |
2895,1±19,43*** |
3074,2± 21,22*** |
2962,0±18,17*** |
|
61 |
3563,9±24,36 |
3749,1±25,01*** |
3947,2±30,30*** |
3801,1±27,23*** |
|
66 |
3990,4±48,62 |
4194,0±47,21** |
4419, 7±53,13*** |
4265,5±50,68*** |
|
Абсолю |
||||
|
тный |
||||
|
прирост |
3894,5±48,61 |
4098,5±48,02** |
4323,5±50,06*** |
4166,5±49,41*** |
|
в % к I |
||||
|
группе |
100 |
105,2 |
111,0 |
107,0 |
Периодическое индивидуальное взвешивание гусят в течение всего периода выращивания и откорма показывает, что глауконит в изучаемых
дозировках оказывает не одинаковое влияние на рост и развитие птицы. При одинаковой постановочной живой массе живая масса гусят в конце первой десятидневки имела различие. Причём лучшие результаты показала низкая дозировка глауконита. Так, если в контрольной группе средняя живая масса одной головы была 381,9 г, то во второй группе она была выше на 34,2 г, или на 9%. В то время как в третьей и четвёртой группах различие составило соответственно 11,3 и 28,1 г, или 3,0 и 7,4 % (Р< 0,01, Р< 0,001).
В 21-дневном возрасте средняя живая масса одного гусёнка в опытных группах была практически одинаковой ( 873,1 - 877,5 г ), но превосходила первую контрольную группу на 60,2 - 63,6 г ( Р< 0,001). С третьей недели опыта и до его конца наибольшую интенсивность роста имели гусята третьей группы, получавшие с рациона 0,2% глауконита. В месячном возрасте их живая масса превосходила аналогов первой контрольной группы на 11,5%, в 41 - дневном - на 12,9, в 51-дневном -на 11,3, в конце опыта- на 10,8% ( Р< 0,001).
Низкая дозировка глауконита (II группа) позволила в аналогичные возрастные периоды увеличить живую массу гусят по сравнению с I группой на 4,8-6,4% (Р< 0,05, Р< 0,01, Р< 0,001). А высокая дозировка глауконита в рационах гусят IV группы повысила их живую массу по сравнению с I контрольной группой в возрасте 31, 41, 51 и 66 дней соответственно на 9,7%, 8,7, 7,2 и 6,9 %.
В результате чего в среднем за опыт абсолютный прирост живой массы гусёнка в I группе составил 3894,5 г, в то время как во II группе -4098,5, в III группе - 4323,5 , в IV группе - 4166,5 г, что соответственно выше по сравнению с первой группой на 5,2%, 11,0 и 7,0%.
Интенсивность роста молодняка птицы определяется по относительному приросту и её живой массы. Результаты расчёта относительного прироста живой массы гусят за период опыта составлены в таблице 6.
6. Относительный прирост живой массы гусят, % ( п = 50 )
|
Возраст, дн |
Группа |
|||
|
I |
II |
III |
IV |
|
|
11 |
119.7 |
124.9 |
121.4 |
124.1 |
|
21 |
72.1 |
70.9 |
76.0 |
72.6 |
|
31 |
51.7 |
50.8 |
55.0 |
53.2 |
|
41 |
37.3 |
36.9 |
38.5 |
36.3 |
|
51 |
31.5 |
30.3 |
30.0 |
30.1 |
|
61 |
25.3 |
25.7 |
24.9 |
24.8 |
|
66 |
11.3 |
11.2 |
11.3 |
11.5 |
Вышеприведённые данные изменений относительного прироста живой массы гусят показывают, что самая высокая их интенсивность роста наблюдается в первые 10 дней жизни. Если в I группе она составила 119,7%, то в опытных группах она была на уровне 121,4 - 124,9%.
К шестьдесят шестому дню, т.е. к окончанию эксперимента, интенсивность роста гусят снизилась до 11,2 - 11,5% во всех группах. Характерно отметить, что низкая дозировка глауконита у гусят II группы с 21 по 51 день обеспечивала интенсивность роста, уступающего аналогам I группы. Средняя дозировка глауконита (III группа ) позволила до 41 дня иметь интенсивность роста гусят на 1,2-3,9 % выше, чем в I группе, а в последующем на 0,4 - 1,5 % уступала им. Высокая дозировка глауконита ( IV группа ) только в течение первого месяца жизни гусят обеспечивала их более высокую интенсивность роста, а в последующем она уступала аналогам I контрольной группы.
Изменение абсолютного и относительного прироста живой массы гусят в течение научно - хозяйственного опыта объясняется разницей в среднесуточном приросте. Проведённый расчёт среднесуточного прироста гусят за период опыта представлен в таблице 7 и на рисунке 1.
7. Динамика среднесуточного прироста гусят, г ( X ± m>x>, n =50)
|
Возраст, дн |
Группа |
|||
|
1 |
II |
III |
IV |
|
|
1 |
- |
- |
- |
- |
|
11 |
28,6 ±0,27 |
32,0±0,23*** |
29,7 ±0,25** |
31,4±0,29*** |
|
21 |
43, 1± 0,64 |
45,7 ±0,82** |
48,2±0,79*** |
46,7±0,80*** |
|
31 |
56,7± 2,94 |
59,5 ± 2,40 |
66,4 ±2,65** |
63,7 ±3,02 |
|
41 |
63,2± 2,93 |
66,5± 2,85 |
73,3 ±3,13* |
67,2 ± 3,44 |
|
51 |
75,1±3,13 |
76,2 ±3,35 |
80,2 ±3,51 |
77,6 ± 3,22 |
|
61 |
80,1 ±2,46 |
85,4 ±2,39 |
87,3 ±2,61 |
83,9 ±2,48 |
|
66 |
85,3 ±3,07 |
89,1± 2,91 |
94,5 ±3,17* |
92,3 ±3,55 |
|
В целом за опыт |
59,9± 0,75 |
63,0± 0,70** |
66,5±0,81*** |
64,1±0,79*** |
|
В % к I группе |
100 |
105,2 |
111,0 |
107,0 |
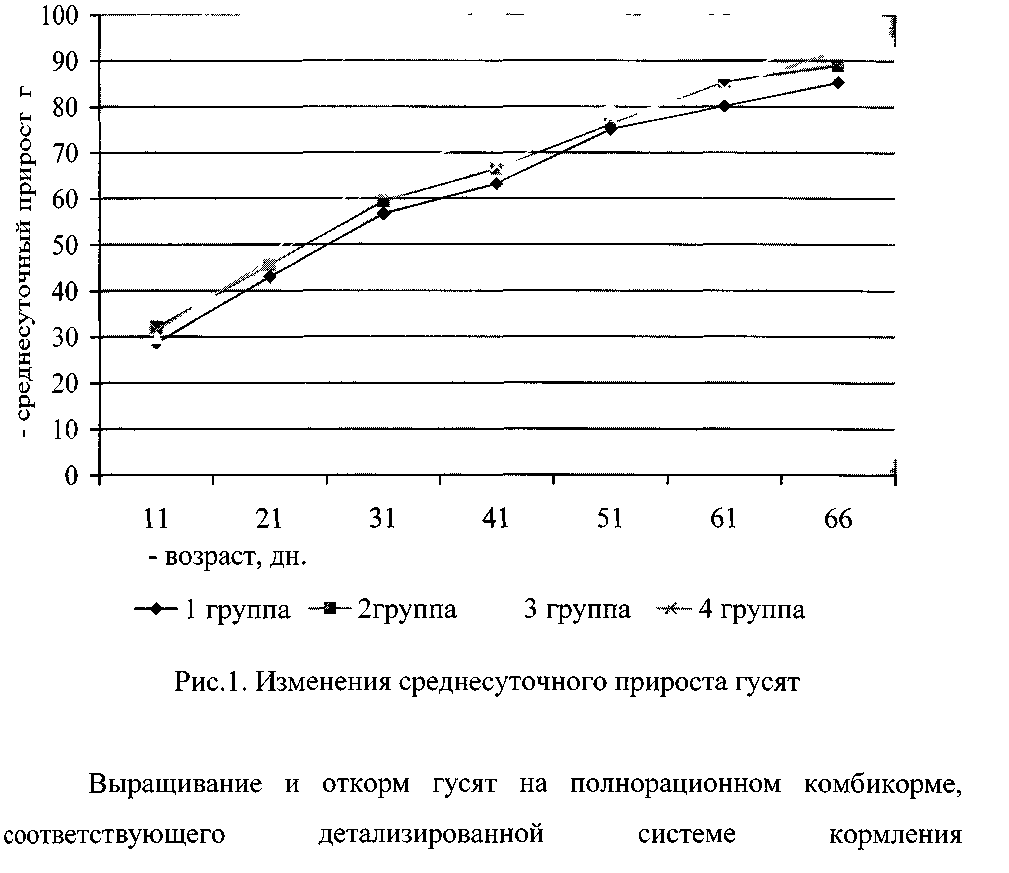
сельскохозяйственной птицы, позволяет ежедневно увеличивать прирост гусят. Если к концу первого периода выращивания (21 день) среднесуточный прирост гусят I группы был на уровне 43,1 г, то к концу откорма (66 день) он возрос до 85,3 г.
Высокие ионообменные свойства глауконита определённым образом повлияли на обменные процессы в организме птицы. Если в конце первой декады наиболее положительное воздействие на организм проявили низкая и высокая дозировки глауконита (Пи IV группы ), в результате чего по сравнению с I контрольной группой разница в среднесуточном приросте составила 3,4 и 2,8 г ( Р< 0,001 ), то средняя дозировка глауконита ( III группы) обеспечила различие всего лишь в 1,1 г ( Р< 0,01).
Однако к концу первого периода выращивания (к 21 дню) наибольший среднесуточный прирост наблюдается в III группе, по сравнению с I контрольной, II и IV опытными группами. Так, если в I группе среднесуточный прирост составил 43,1 г, то во II группе он был выше на 2,6 г ( Р< 0,01), в III - на 5,1 и в IV группе на 3,6 г ( Р< 0,001 ). Данная закономерность сохранилась до конца опыта, при чём с достоверными различиями у гусят III группы по сравнению с I группой в возрасте 31 день на 9,7 г ( Р< 0,01), в 41 день - 10,1 г (Р< 0,5 ), в 66 дней на 9,2 г ( Р< 0,5 ). Во II и IV группах в течение одного возрастного периода среднесуточный прирост гусят был выше чем в I группе, но не имел достоверных различий.
В конечном итоге в целом за опыт среднесуточный прирост гусят I группы составил 59,9 г, во II группе он был выше на 3,1 г ( Р< 0,01), или на 5,2 %, в III - 6,6 г ( Р< 0,001 ), или на 11% и в IV группе - на 4,2 г( Р< 0,001 ), или на 7% по сравнению с I группой.
На протяжении всего научно-хозяйственного опыта нами ежедневно проводился учёт павшей птицы и постановка патологоанатомического диагноза. Падёж и выбраковка гусят в течение эксперимента представлены в таблице 8.
8. Падёж и сохранности гусят за период опыта, гол. ( п = 500 )
|
Возраст, дн. |
Группа |
|||
|
I |
II |
III |
IV |
|
|
1-10 |
8 |
4 |
3 |
3 |
|
11-20 |
4 |
3 |
1 |
3 |
|
21-30 |
2 |
- |
1 |
2 |
|
31-40 |
9 |
4 |
- |
- |
|
41-50 |
1 |
1 |
- |
- |
|
51-60 |
2 |
1 |
- |
- |
|
61-66 |
- |
- |
- |
- |
|
Итого: |
26 |
13 |
5 |
8 |
|
Сохранность, % |
94,8 |
97,4 |
99,0 |
96,4 |
Следует отметить, что в первую декаду жизни гусят основной причиной их выбраковки явилась токсическая дистрофия. По всей вероятности, родительское стадо гусей в ЗАО «Птицевод» перед и в момент яйцекладки получало не доброкачественные корма, что в конечном итоге сказалось на новорожденных гусятах. Если в I контрольной группе за данный период отход гусят составил 8 голов, то добавка глауконита снизила падёж во второй группе на 4 головы, в III и IV группах по 3 головы.
В следующие возрастные периоды ( 11-20 и 21-30 дней ) основная причина падежа гусят связана с их травмированием при перегонах из помещения на выгульные площадки. Но при этом в опытных группах выбраковка гусят была меньше по сравнению с I контрольной группой.
Всплеск падежа гусят в возрастной период 31-40 дней, когда в I контрольной группе пало 9 голов, а во II - 4 объясняется ветеринарной службой хозяйства, завозом недоброкачественной партии корма, что послужило появлению у гусят энтерита. Глауконит, в результате его высокой сорбционной способности к токсинам, снизил падёж гусят, а в III и IV группах вообще его не было. Единичные случаи выбраковки гусят в I и II группах в последующие 26 дней были связаны с травмированием птицы. В конечном итоге за период опыта в I контрольной группе из всего поголовья по различным причинам выбыло 26 гусят, во II -13, в III -5 и в IV группе - 8 голов. В результате чего если сохранность поголовья в I группе составила 94,8 %, то во II группе она увеличилась до 97,4, в III - до 99,0 и в IV группе - 96,4 %. Объяснить изменение живой массы гусят под влиянием кормовой добавки глауконита возможно постановкой и проведением балансовых опытов с расчётом коэффициентов переваримости и использования основных питательных веществ корма.
3.2 Физиологические исследования
На течение процессов пищеварения в организме птицы оказывают влияние многие факторы, такие как возраст, пол, кросс птицы, направление продуктивности, рацион. Основными показателями изучения физиологии пищеварения в научных работах по вопросам полноценного кормления птицы является определение коэффициентов переваримости питательных веществ рациона. Одной из поставленных перед нами задач являлось рассчитать в балансовом опыте коэффициенты переваримости рациона гусят-бройлеров под влиянием кормовой добавки глауконита.
3.2.1 Влияние глауконита на переваримость питательных веществ рациона
В период проведения балансового опыта гусята контрольной и опытных групп получали одинаковый рацион в равном количестве. Полная поедаемость кормосмеси обеспечила ежесуточное потребление питательных веществ подопытной птицей, количество которых представлено в таблице 9.
9. Химический состав кормосмеси и среднесуточное потребление питательных веществ гусятами
|
Показатели |
Химический состав кормосмеси,% |
Содержится в 325 г кормосмеси, г |
|
Сухое вещество |
85,4 |
278,0 |
|
Органическое вещество |
81,1 |
264,0 |
|
Сырой протеин |
18,6 |
60,0 |
|
Сырой жир |
4,5 |
14,6 |
|
Сырая клетчатка |
6,15 |
20,0 |
|
БЭВ |
51,85 |
169,0 |
|
Кальций |
1,32 |
4,29 |
|
Фосфор |
0,668 |
2,17 |
Добавка глауконита в кормосмесь гусят II -IV опытных групп оказала определённое влияние на течение обменных процессов, что в свою очередь отразилось на содержании питательных веществ в помёте гусят (табл.10).
10. Содержание питательных веществ в помёте гусят ( в среднем за сутки, )г (X ±nix, n=5)
|
Показатель |
Группа |
|||
|
I |
II |
III |
IV |
|
|
Сухое вещество |
67,17±1,06 |
65,80±1,28 |
63,43±1,57 |
63,19±2,19 |
|
Органическое вещество |
58,65±1,13 |
55,32±1,33 |
52Д4±1,92* |
53,06±2,09 |
|
Сырой протеин |
6,89±0,20 |
5,48±0,22 |
3,88±0,24*** |
4,74±0,37*** |
|
Сырой жир |
6,98±0,31 |
5,22±0,17*** |
5,80±0,11** |
5,56±0,08** |
|
Сырая клетчатка |
14,16±0,08 |
13,90±0,12 |
13,86±0,17 |
13,64±0,32 |
|
БЭВ |
30,62±1,34 |
30,72±1,67 |
28,60±2,01 |
29,12±2,35 |
Вышеприведённые данные таблицы 10 свидетельствуют, что с увеличением дозировки кормовой добавки глауконита в рационе гусят II и IV fопытных групп, по сравнению с I контрольной группой, наблюдается
тенденция к снижению сухого вещества в помёте. Так, если в I группе его содержание в помёте было на уровне 67,17 г, то во II группе оно снизилось на 1,37 г, в III - на 3,74 и в IV группе - на 3,98 г. Аналогичная закономерность наблюдается и количеству органического вещества в помёте. Причём, если во II группе, по сравнению с I, различие составило 3,33 г (Р >0,05 ), в IV -на 5,59 г (Р >0,05 ), то в III группе оно было самым высоким-6,51 г (Р<0,05). Снижение содержания органического вещества в помёте гусят II-IV опытных групп произошло в основном за счёт уменьшения количества сырого протеина и сырого жира.
По сравнению с помётом гусят I группы содержание сырого протеина во II группе уменьшилось на 1,41 г , или на 20,5%, в III- на 3,01 г, или 43,7%, и в IV группе на 2,15 г, или 21,2% (Р <0,001 ). Самое низкое содержание сырого жира наблюдалось в помёте гусят II группы и составило 5,22 г, затем в IV - 5,56 и в III группе - 5,80 г, что было ниже по сравнению с I группой собственно на 25,2%, 16,9 и 19,3% (Р <0,01, Р<0,001).
По содержанию в помёте гусят сырой клетчатки наблюдается тенденция к её уменьшению в группах, где птица получала кормовую добавку глауконит. Данное различие было в пределах 0,26 - 0,52 г. По группе БЭВ уменьшение наблюдалось только в III и IV группах и не имело достоверного различия.
Имеющаяся разница в выделении питательных веществ с помётом при одинаковом их поступлении в организме гусят позволило рассчитать коэффициенты переваримости, представленные в таблице 11 и на рисунках 2
11 .Коэффициенты переваримости питательных веществ кормосмеси гусятами (X ± nix, n =5)
|
Показатель |
Группа |
|||
|
I |
II |
III |
IV |
|
|
Сухое вещество |
75,8±0,38 |
76,7±0,47 |
77,2±0,56 |
77,3±0,78 |
|
Органическое вещество |
77,8±0,43 |
79,0±0,51 |
80,2±0,72* |
79,9±0,79* |
|
Сырой протеин |
88,6±0,33 |
91,0±0,35«* |
93,6±0,41*** |
92,2±0,60*** |
|
Сырой жир |
52,2±2,12 |
64,2±1Д9*** |
60,3±0,78** |
61,9±0,55** |
|
Сырая клетчатка |
29,2±0,37 |
30,5±0,59 |
30,7±0,86 |
31,8±1,61 |
|
БЭВ |
81,9±0,78 |
81,8±0,98 |
83,1±1,19 |
82,8±1,39 |

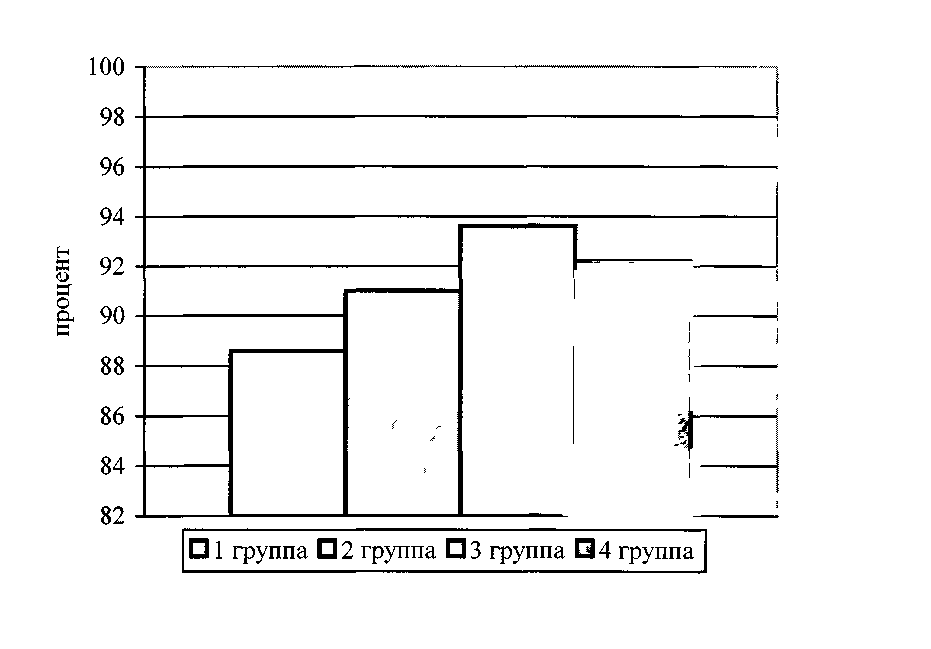 Рис.3.
Коэффициенты переваримости сырого
протеина гусятами
Рис.3.
Коэффициенты переваримости сырого
протеина гусятами
Переваримость питательных веществ, кормов, поступающих в организм, во многом зависит от ферментативной активности желёз внутренней секреции, секреторной функции отделов желудочно-кишечного тракта и отдельных органов. Биогенные микроэлементы, освобождающиеся из глауконита за счёт его ионообменных свойств, по всей вероятности, активизируют работу ферментов.
При кормлении гусят одним только полнорационным комбикормам (I группа) переваримость сухого вещества кормосмеси находилась на уровне 75,8%, добавка к рациону 0,1% глауконита (II группа) повышает его переваримость на 0,9%, увеличение дачи глауконита до 0,2% (III группа) - на 1,4, до 0,3 % (IV группа) - на 1,5%. Аналогичная закономерность наблюдается и с органическим веществом. Если в I группе его переваримость составила 77,8%, то во II группе она возросла на 1,2%, в III- на 2,4 (Р< 0,05 ) и в IV группе - на 2,1% (Р < 0,05).
Повышение переваримости органического вещества рациона гусят опытных групп произошло в основном за счёт переваримости сырого протеина и сырого жира. Самая высокая переваримость протеина наблюдалась в III группе и составила 93,6%, затем в IV - 92.2 и во II группе -91,0%, что выше по сравнению с I группой соответственно на 5,0%,3,6 и 2,4% (Р< 0,001).
Однако, низкая дозировка глауконита, в отличие от средней и высокой способствует более высокой переваримости сырого жира в организме птицы. Так, если в I группе переваримость сырого жира составила 52,2%, то во II группе она возросла на 12,0% (Р< 0,001 ) и составила 64,2%, в III группе - на 8,1% (Р < 0,01 ) и составила 60,3%, в IV группе- на 9,7% и составила 61,9% (Р < 0,01 ).Существенных различий между группами в переваримости сырой клетчатки и БЭВ не наблюдалось. Переваримость сырой клетчатки находилась в пределах от 29,2% у гусят I группы до 31,8%- в IV группе, а переваримость БЭВ - от 81,8% во II группе до 83,1% в III группе.
Таким образом, на переваримость питательных веществ рационов гусят-бройлеров глауконит оказывает не одинаковое влияние, низкая дозировка в большей степени влияет на переваримость сырого жира, средняя - сырого протеина, высокая дозировка — на переваримость сырого протеина и жира, но в меньшей степени, чем низкая и средняя дозировки.
3.2.2 Баланс и использование азота
Азотистые вещества корма, попадая в желудочно-кишечный тракт животного, подвергаются гидролизу до свободных аминокислот, которые используются для роста и развития растущего организма, восстановления изношенных тканей, роста и развития плода.
Поэтому об эффективности использования протеина корма в различные возрастные периоды у сельскохозяйственных животных и птицы можно судить по балансу азота корма.
Анатомические особенности пищеварительной системы и органов мочевыделения у птицы позволяют одномоментно учитывать весь выделяющий из организма азот, а количество отложенного азота в теле рассчитать по формуле:
N отложенный в теле = N корма - (N кала + N мочи )
N кала + N мочи = N помёта
Результаты проведённого расчёта баланса азота у гусят, получавших на фоне основного рациона кормовую добавку глауконит в изучаемых дозировках, представлен в таблице 12 и на рисунке 4.
12 Баланс азота у гусят ( в среднем за сутки, г) (X±mx, n =5)
|
Показатель |
Группа |
|||
|
I |
II |
III |
IV |
|
|
Принято с кормом |
9,70 |
9,70 |
9,70 |
9,70 |
|
Выделено с помётом |
6,30± 0,04 |
6,19±0,03* |
6,00± 0,03*** |
6,14±0,02** |
|
Отложено в теле |
3,40± 0,04 |
3,51± 0,03* |
3,70± 0,03*** |
3,56 ±0,02** |
|
Использовано, % от принятого |
35,1±0,47 |
36,1±0,31 |
38,1±0,33*** |
36,7± 0,25* |
|
ЗРис. 4. Среднесуточное отложение азота в теле гусят При одинаковом среднесуточном поступлении азота с кормом в организм гусят различия в его переваримости и усвоения под влиянием изучаемых дозировок глауконита привели к тому, что если в I контрольной группе с помётом из организма выделялось 6,3 г азота, то во II группе его потери уменьшились на 0,11 г (Р < 0,05 ), в III - на 0,3 (Р < 0,001 ) и в IV группе- на 0,16 г (Р< 0,01 ). В результате чего на аналогичную величину в опытных группах возросло среднесуточное отложение азота в теле и составило: в I группе 3,40 г, во II - 3,51, в III - 3,70 и в IV группе - 3,56 г. Соответственно и самое высокое использование азота отложенного в теле гусят в расчёте от принятого было в III группе и составило 38,1%, в меньшей степени в IV и в III — 36.7 и 36,1%, самое низкое использование азота наблюдалось в I контрольной группе - 35,1%. Следовательно, самое высокое использование азота в теле гусят-бройлеров наблюдается при включении в состав основного рациона кормовой добавки глауконит в количестве 0,2 % от сухого вещества рациона. 3.2.3 Баланс кальция и фосфора / Кальций и фосфор относятся к макроэлементам не органической части корма и являются одними из важных минеральных элементов детализированной системы кормления. Биологическая роль данных элементов питания для организма птицы очень велика. Кальций содержится в основном в костях в виде фосфорнокислых и углекислых солей. Ионы кальция необходимы для нормальной деятельности сердца, участвуют в регуляции мышечной и нервной деятельности, повышают защитные функции организма. Дефицит кальция в рационе молодняка приводит к возникновению рахита. В основе заболевания лежит расстройство процессов минерализации кости. Это приводит к нарушению роста, искривлению позвоночника, рёбер, трубчатых костей и клюва. У взрослой птицы при недостатке кальция развивается остеопороз. Это обусловлено нарушением эндокринных механизмов, в первую очередь гипофизарного и паратиреоидного. Предрасполагающим фактором служит ограниченное движение птицы. Поэтому одной из поставленных в наших исследованиях задач являлось рассчитать баланс кальция в рационе гусят-бройлеров при скармливании им кормовой добавки глауконита. Среднесуточный баланс кальция в организме гусят представлен в таблице 13. 13. Баланс кальция у гусят (в среднем на голову в сутки ,г) (X ±nix, n =5) Показатель |
Группа |
|||
|
I |
II |
III |
IV |
|
|
Принято с кормом |
4,29 |
4,29 |
4,29 |
4,29 |
|
Выделено с помётом |
1,86 ±0,05 |
1,89±0,06 |
1,39 ±,06*** |
1,47±0,08** |
|
Отложено в теле |
2,43± 0,05 |
2,40± 0,06 |
2,90 ±0,06*** |
2,82± 0,08** |
|
Использовано, % от принятого с кормом |
56,6±1,11 |
55,9± 1,35 |
67,6± 1,33** |
65,7± 1,94** |
При одинаковом поступлении кальция в организм гусят-бройлеров его потери с продуктами выделения в помёте между группами были не одинаковы. Так, если в I контрольной группе общее количество кальция выделенного с пометом было на уровне 1,86 г в сутки, то низкая дозировка глауконита у гусят II группы не повлияла на его потери с помётом (1,89 г), средняя дозировка глауконита (III группа) способствовала снижению его выделения с помётом на 0,47 г (Р < 0,001 ), а высокая дозировка в рационе гусят IV группы обеспечила данное различие в 0,39 (Р < 0,01 ) по сравнению с I группой. В результате чего, если в I и II группах среднесуточное отложение кальция в теле гусят составило соответственно 2,43 и 2,40 г, то в III группе оно было на уровне 2,90 г, а в IV группе - 2,82 г. Поэтому в III и IV группах наблюдалось самое высокое использование кальция по отношению к принятому с кормом (67,6 и 65,7 %), в то время как в I группе оно составило 56,6%, а во II группе- 55,9%.
Фосфор, как и кальций, содержится во всех тканях организма и является непременным компонентом его внутренней среды. Основная часть фосфора в виде фосфорнокислого кальция находится в костяке.
Фосфорная кислота входит в состав многих коэнзимов. Макроэргические фосфорные соединения (АТФ, АДФ, АМФ) являются универсальными аккумуляторами энергии, исключительную роль играет АТФ в мышечной деятельности. Дефицит фосфора в рационах молодняка приводит к развитию рахита, а у взрослой птицы вызывает остеопороз.
Расчёт баланса фосфора у гусят, получавших кормовую добавку глауконит, представлен в таблице 14
14. Баланс фосфора у гусят (в среднем на голову в сутки, г) (X±mx, n =5)
|
Показатель |
Группа |
|||
|
I |
II |
III |
IV |
|
|
Принято с кормом |
2,17 |
2,17 |
2,17 |
2,17 |
|
Выделено с |
||||
|
помётом |
1,05±0,06 |
1,01±0,05 |
0,94± 0,04 |
1,1 0± 0,06 |
|
Отложено в теле |
1,12±0,06 |
1,16±0,05 |
1,23±0,04 |
1,07±0,06 |
|
Использовано, % |
||||
|
от принятого с |
||||
|
кормом |
51,6±3,97 |
53,5 ±2,50 |
56,7 ±1,67 |
49,3 ±2,80 |
При среднесуточном потреблении гусятами 2,17 г фосфора в сутки его потери с помётом были практически одинаковыми : 1,05 г в I группе, 1,01- во II, 0,94 в III и 1,10 г - в IV группе. В результате чего среднесуточное отложение фосфора в теле гусят I группы было на уровне 1,12 г, во II - 1,16,
в III - 1,23 и в IV группе 1,07 г, а его использование от принятого по группам соответственно составило 51,6%, 53,5, 56,7 и 49,3%.
Следовательно, глауконит в меньшей степени оказывает влияние на переваривание и отложение в теле фосфора, в то время как наибольшее отложение кальция в организме наблюдается при средней дозировке изучаемого алюмосиликата, в меньшей степени высокой, и низкая дозировка глауконита не оказывает никакого влияния на повышение его ретенции в организме гусят.
3.3 Гематологические исследования
Кровь как жидкая ткань вместе с лимфой, спинномозговой и межтканевой жидкостями составляют то единое целое, которое называют внутренней средой организма. Кровь практически омывает все клетки, доставляя к ним необходимые вещества и унося от них продукты жизнедеятельности. Состав крови свидетельствует о нормальных и патологических процессах, происходящих в организме. Как внутренняя среда она осуществляет связь всех органов и тканей, создавая необходимый режим их существования.
О направленности обменных процессов в живом организме можно судить по содержанию отдельных метаболитов в крови. Их количество может меняться в зависимости от кормления, то есть под влиянием отдельных кормовых добавок.
В наших исследованиях была поставлена задача изучить изменения отдельных морфологических и биохимических показателей крови гусят при скармливании им кормовой добавки глауконит. Полученные результаты представлены в таблице 15.
15. Отдельные морфологические и биохимические показатели крови гусят (Х± m>x>, n =5)
|
Показатель |
Группа |
|||
|
I |
II |
III |
IV |
|
|
Гемоглобин, г/л |
87,5±1,12 |
87,5± 1,75 |
93,7±0,95** |
88,7± 0,01 |
|
Эритроциты, 1012/л |
2,34± 0,26 |
2,34± 0,33 |
2,59±0,18 |
2,64± 0,17 |
|
Лейкоциты,! О9 /л |
25,7±ОД8 |
25,3±0,11 |
27,5± 0,67* |
24,7± 0,30 |
|
Общий белок, г/л |
63,5±3,56 |
64,5± 2,50 |
82,5±5,17** |
71,0±3,10 |
|
Аминный азот, г/л |
0,06± 0,001 |
0,06± 0,001 |
0,07± 0,001*** |
0,06± 0,001 |
|
Общие липиды, г/л |
4,64±0,11 |
5,24±0,17* |
4,89± 0,21 |
5,10±0,28 |
|
Холестерин, моль/л |
2,70± 0,22 |
2,75±0,19 |
2,70±0,17 |
2,50±0,18 |
|
В - липопротеиды, г/л |
2,62±0,31 |
2,82± 0,20 |
3,46± 0,12* |
3,08±0,16 |
|
Кальций, ммоль/л |
2,67±0,11 |
2,90± 0,08 |
2,90± 0,07 |
2,78± 0,08 |
|
Фосфор, ммоль/л |
1,81±0,17 |
2,00± 0,06 |
2,35± 0,23 |
1,94±0,05 |
Эритроциты являются важными форменными элементами крови. Обладая большой удельной поверхностью, эритроциты могут адсорбировать на себе многочисленные органические и минеральные вещества, в том числе и газы, и транспортировать их к тканям. Кормовая добавка глауконит в рационах гусят-бройлеров опытных групп не оказала существенного влияния на количество эритроцитов, хотя в III и IV группах по сравнению с I имеется тенденция к их повышению на 0,25- 0,30х 1012 /л, или на 10,7 - 12,8%.
Основная физиологическая функция эритроцитов – дыхательная, неразрывно связана со свойствами содержащегося в них белка гемоглобина. В лёгких гемоглобин присоединяет к себе кислород и транспортирует его к тканям. В тканях, отдав кислород, эритроциты связывают углекислый газ и транспортирует его к лёгким.
Если в I группе гусят-бройлеров содержание гемоглобина в крови было на уровне 87,5 г/л, то во II группе его было аналогичное количество, что и в I группе, в то время как в III группе оно возросло на 5,2 г/л ( Р< 0,01), а в IV группе- на 1,2 г/л ( Р> 0,05).
Лейкоциты относятся к форменным элементам крови и играют очень важную роль в защите организма. Они могут захватывать и переваривать бактерии и другие инородные тела, попавшие в организм. Анализируя содержание лейкоцитов в крови гусят-бройлеров, видно, что только в III группе, по сравнению с остальными, количество лейкоцитов достоверно увеличилось и составило 27,5х 109 /л, что на 7,0 % больше чем в I группе, на 8,7- чем во II и на 11,3% по сравнению с IV группой.
Содержание общего белка в крови во многом зависит от уровня сырого протеина в рационе его переваримости в организме птицы. Различия в коэффициентах переваримости протеина под влиянием кормовой добавки глауконит привело к тому, что если в I группе количество общего белка в сыворотке крови составило 63,5 г/л, то в остальных группах его было больше: во II - на 1,0 г/л, в III- на 19,0 (Р< 0,01), и в IV группе - на 7,5 г/л.
Одним из важных показателей белкового обмена является определение в сыворотке крови аминного азота, т.е. азота свободных аминокислот, поступивших в кровь после гидролиза белка в желудочно-кишечном тракте. У всей подопытной птицы, за исключением III группы, количество аминного азота в крови было одинаковым - 0,06 г/л, а в III группе оно увеличилось на 16,7% и составило 0,07 г/л (Р<0,001).
Определение в сыворотке крови отдельных метаболитов липидного обмена показало, что самое высокое содержание общих липидов наблюдалось у гусят II группы, получавших низкую дозировку глауконита, и составило 5,24 г/л, что на 0,6 г/л было выше аналогов I контрольной группы (Р< 0,05). В III и IV группах гусят количество общих липидов в крови превосходило I группу соответственно на 0,25 и 0,46 г/л (Р > 0,05). Основными переносчиками липидов в крови являются В-липопротеиды. Их содержание в крови подопытных гусят имело существенные различия.
Если в I группе количество В - липопротеидов было на уровне 2,62 г/л, то во II группе их содержание повысились на 7,6%, в III - 32,1(Р< 0,05) и в IV гpynne - на 17,6 % , что соответственно составило 2,82 г/л, 3,46 и 3,08 г/л.
Содержание холестерина в сыворотке крови гусят всех групп было практически одинаковое и изменялось от 2,50 ммоль/л в IV группе до 2,75 ммоль/л - во II группе ( Р > 0,05).
Различия содержания в сыворотке крови гусят основных макроэлементов - кальция и фосфора не носило достоверного характера, хотя можно сказать о имеющейся тенденции в повышении их количества у гусят опытных групп. Так, если в I группе количество кальция в крови гусят составило 2,67 ммоль/л, то во Пи III группах оно увеличилось до 2,90, а в IV - до 2,78 ммоль/л. Содержание фосфора в крови I группы гусят составило 1,81 ммоль/л, во II группе оно увеличилось до 2,00, в III - до 2,35 и в IV группе - до 1,94 ммоль/л.
Высокие ионообменные свойства глауконита оказывают влияние не только на лучшее усвоение макроэлементов из рациона гусят, но и также определённым образом влияют на поступление в организм и микроэлементов, выполняющих в организме важные функции в обмене веществ. Содержание отдельных микроэлементов в цельной крови гусят-бройлеров представлено в таблице 16.
|
Показатель |
Группа |
|||
|
I |
II |
III |
IV |
|
|
Медь |
29,30±2,31 |
27,82± 1,18 |
32,32± 1,55 |
33,40± 1,16 |
|
Кобальт |
0,10±0,01 |
0,09±0,01 |
0,13±0,01* |
0,09±0,01 |
|
Цинк |
5,98 ±0,45 |
7,54 ±0,51* |
6,42± 0,33 |
4,35± 0,28 |
|
Железо |
1,85±0,25 |
1,93 0±,28 |
2,43 ±0,1 8 |
2,01±0,11 |
|
Марганец |
0,10 ±0,02 |
0,06± 0,03 |
0,08 ±0,02 |
0,09 ±0,01 |
Если в I группе гусят его количество в крови составило 5,98 мг/л, то во II группе по содержанию он увеличился на 26,1% и был на уровне 7,54мг/л (Р< 0,05). Достоверных различий по содержанию в крови гусят II группы меди, кобальта, железа и марганца отмечено не было, имеющаяся разница была в пределах ошибки средней арифметической величины.
Средняя дозировка глауконита в рационе гусят III группы, по сравнению с I, достоверно повысила содержание кобальта в крови с 0,10 до 0,13 мг/л (Р< 0,05). При этом у данной группы гусят следует отметить позитивные сдвиги в увеличении количества в крови меди на 10,3%, цинка -на 7,4 и железа - на 31,4% (Р> 0,05).
Добавка к рациону гусят-бройлеров IV группы высокой дозировки глауконита привело к тому, что в их крови по сравнению с I группой количество меди повысилось с 29,30 мг/л до 33,40 мг/л, железа - с 1,85 до 2,01 мг/л и наблюдается снижение кобальта и марганца с 0,10 до 0,09мг/л, цинка- с 5,98 до 4,35 мг/л (Р> 0,05).
Следовательно, средняя дозировка глауконита в рационах гусят-бройлеров по сравнению с низкой и высокой оказывает стимулирующее влияние на эритропоэз в организме птицы, повышает окислительно-восстановительные реакции обмена веществ с выраженным анаболическим характером и защитные функции организма.
3.4 Результаты контрольного убоя гусят-бройлеров
Мясная продуктивность - важнейшие хозяйственно полезное свойство птицы. Она характеризуется массой и мясными качествами птицы в убойном возрасте, а также пищевыми достоинствами - качеством мяса.
Мясо птицы, как и других сельскохозяйственных животных, состоит из белков, ряда азотистых и безазотистых соединений, углеводов, жиров, витаминов, ферментов, минеральных веществ и воды. Значение и ценность мяса, а также других пищевых продуктов для рационального питания человека определяются его калорийностью, вкусовыми свойствами и степенью усвояемости. Пищевая ценность мяса обуславливается соотношением входящих в его состав тканей. Чем больше в мясе мышечной ткани, тем выше его питательная ценность. Жировая ткань является благоприятным фактором только при определённом её соотношении с мышечной и соответствующем распределении в тушке. При большом количестве жировой ткани уменьшается относительное содержание белков и снижается усвояемость мяса. По мере увеличения количества соединительной ткани, содержащей неполноценные белки, снижается количество мяса, уменьшается его нежность и ухудшается вкус. Кости также понижают пищевую ценность мяса, так как в них мало питательных веществ.
Результаты контрольного убоя птицы, проведённого в конце периода откорма представлены в таблице 17.
17. Анатомическая разделка тушек гусей, в % к предубойной массе (п=5)
|
Показатель |
Группа |
|||
|
I |
II |
III |
IV |
|
|
Предубойная масса, г |
3854 ±25,0 |
4072±24, 6*** |
4300 ±42,4*** |
4146±38,4*** |
|
Масса не потрошённой тушки, г в % к предубойной массе |
3426± 44,4 88,9 |
3632±34,3** 89,2 |
3853 ±32,9*** 89,6 |
3698 ±44,6** 89,2 |
|
Масса полупотрощённой тушки, г |
3106±36,4 |
3290 ±37,4** |
3492±29,1*** |
3362±9,8*** |
|
Убойный выход полу потрошённой тушки, % |
80,6 |
80,8 |
81,2 |
81,1 |
|
Масса потрошенной тушки ,г |
2821 ±16,2 |
2989±33,9** |
3173±20,7*** |
3064±26,8*** |
|
Убойный выход потрошённой тушки, % |
73,2 |
73,4 |
73,8 |
73,9 |
|
Выход съедобных частей, г % |
2337 ±17,9 68,2 |
2508 ±15,9*** 69,1 |
2660 ±25,3*** 69,0 |
2581± 19,9*** 69,8 |
|
Выход не съедобных частей тушки, г % |
1089±38,8 31,8 |
1124±22,7 30,9 |
11 93 ±38,5 31,0 |
1117±22,7 30,2 |
|
Индекс мясных качеств |
2,15 |
2,23 |
2,23 |
2,31 |
Представленные данные таблицы 17 показывают, что предубойная живая масса гусят-бройлеров была близкой к средней живой массе гусят в ростовом опыте: 3854 г в I группе, 4072 г - во II, 4300 - в III и 4146 г - в IV группе.
Масса непотрошённой тушки, то есть за вычетом из предубойной массы массу крови, пера и пуха, в относительном выражении изменялась от 88,9% в I группе до 89,6 - в III группе. А в абсолютном выражении, если в I группе масса непотрошённой тушки весила 3426 г, то во II группе она была больше на 206 г (Р< 0,01), в III - на 427 г (Р< 0,01) и в IV группе - на 272 г
Полупотрошённая тушка гусят опытных групп также превосходила по массе контрольную группу. Причём данная разница составила в пользу II группы на 184 г, или на 5,9% (Р< 0,01), в III группе - на 386 г , или 12,4% (Р< 0,01) и в IV группе - на 256 г, или на 8,2% (Р< 0,01).
Проведение полного потрошения тушек гусят показало, что масса потрошённой тушки в I группе была на уровне 2821 г, во II группе 2989, в III - 3173 и в IV группе - 3064 г, или по сравнению с I группой данное различие составило в пользу опытных групп соответственно на 168 г (Р< 0,01), 352 г(Р< 0,001) и 243 г (Р< 0,001). При этом убойный выход потрошённой тушки относительно предубойной массы во всех группах был одинаковым - 73,2-73,9%.
Проведённый расчёт выхода съедобных частей в тушках подопытных гусят существенно отличался в опытных группах по сравнению с контрольной. Так, если в I группе масса съедобных частей в тушке гусят составила 2337 г, то во II группе она была выше на 171 г, в III - на 323 и в IV группе - на 244 г (Р< 0,001). В относительном выражении выход съедобных частей к предубойной массе гусят составил: в I группе 68,2%, во II - 69,1, в III -69,0 и в IV группе - 69,8%.
Более высокая живая масса гусят в опытных группах, по сравнению с контрольной, повысила в их тушках не съедобных частей: во II группе на 35 г, в III - на 104 и в IV - на 28,0 г.
Отношение съедобных частей в тушках гусят к не съедобным позволило рассчитать индекс мясных качеств, который повысился с 2,15 в I группе до 2,23 - во II и в III, до 2,31 - в IV группе.
При проведении контрольного убоя одним из важных качественных показателей является изучение морфологического состава тушек гусят, которое позволяет судить о влиянии кормового фактора на развитее основных тканей организма. Результаты обвалки тушек гусят представлены в таблице 18.
18. Морфологический состав тушек гусят (X±m>x>, n =5)
|
Показатель |
Группа |
|||
|
I |
II |
III |
IV |
|
|
Масса потрошённой |
||||
|
тушки, г |
2821± 16,2 |
2989±33,9** |
3173±20,7*** |
3064±26,8*** |
|
В том числе: |
||||
|
мышц,г |
1543±6,6 |
1650±7,2*** |
1824±18,5*** |
1752±6,4*** |
|
% |
54,7 |
55,2 |
57,5 |
57,2 |
|
Масса внутреннего |
||||
|
жира, г |
79 ±2,3 |
108± 1,2*** |
92±2,9*** |
95± 1,8*** |
|
% |
2,8 |
3,6 |
2,9 |
3,1 |
|
Масса кожи с подкожным жиром, г |
649± 4,7 |
681±3,7*** |
670 ±2,4** |
663± 5,5 |
|
% |
23,0 |
22,8 |
2,11 |
21,6 |
|
Масса костей, г |
550 ±5,1 |
550± 4,7 |
587±3,2*** |
554± 5,5 |
|
% |
19,5 |
18,4 |
18,5 |
18,1 |
|
Мясо-костный индекс |
2,81 |
3,00 |
3,11 |
3,16 |
Полученные данные свидетельствуют, что масса потрошённой тушки у гусят II - IV групп превалировала над I контрольной группой. Причём данное различие составило во II группе на 168 г (Р< 0,01), в III - на 352 г (Р< 0,001)и в IV группе - на 243 г (Р< 0,001). Данное различие произошло за счёт развития всех тканей организма. Так, если в I группе в тушках гусят мышечная ткань в абсолютном выражении составила 1543 г, или 54,7%, от массы потрошённой тушки, то во II группе её количество увеличилось на 107 г (Р< 0,001), или на 0,5%, в III группе соответственно её масса возросла на 281 г и 2,8% (Р< 0,001), а в IV группе - на 209 г и 2,5%.
Самое высокое содержание в тушках внутреннего жира наблюдалось у гусят II группы - 108 г, затем в IV группе - 95 и в III - 92 г, что превосходило I группу соответственно на 29 г, 16 и 13 г (Р< 0,001).
Масса кожи с подкожным жиром аналогичным образом была больше всего во II группе - 681 г, затем в III и IV группах ( 670 и 663 г ) и по сравнению с I группой разница составила соответственно 32 г (Р< 0,001), 21 (Р< 0,01) и 14 г. Хотя в относительном выражении масса кожи с подкожным жиром в I группе была самой высокой и составила 23,0% от массы потрошённой тушки, в то время как во II группе она понизилась до 22,8%, в III - до 21,1 и в IV группе до 21,6%.
Существенных различий по содержанию в тушках гусят I, II и IV групп костной ткани не было, в то время как в III группе её масса на 37 г превосходила I группу г (Р< 0,001).
Важным показателем качества мяса гусят-бройлеров является расчёт мясокостного индекса, то есть отношение мышечной ткани к костной. Добавка глауконита в рацион гусят II - IV групп повысила мясокостный индекс в тушках. Если в I группе он составил 2,81, то во II группе он увеличился до 3,00, в III - до 3,11 и в IV группе - до 3,16.
При анатомической разделке тушки гусят-бройлеров была изучена масса таких жизненно важных органов, как сердце, печень, лёгкое, почки, а также мышечного желудка, результаты данных исследований представлены в таблице 19.
19. Масса внутренних органов гусят, г (X±m>x>, n =5)
|
Орган |
Группа |
|||
|
I |
II |
III |
IV |
|
|
Сердце |
30± 1,52 |
32± 0,83 |
31±0,61 |
33± 0,60 |
|
Печень |
75±2,15 |
78± 0,99 |
76± 1,16 |
74± 0,55 |
|
Мышечный желудок |
127 ±1,75 |
126± 1,47 |
130 ±1,33 |
125± 1,21 |
|
Лёгкое и почки |
35± 0,25 |
33±0,17*** |
37±0,12*** |
38±0,13*** |
Из данных таблицы 19 видно, что глауконит не оказал существенного влияния на массу основных паренхиматозных органов у подопытных гусят. Так, масса сердца изменялась в пределах от 30 г в I группе до 33 г — в IV, масса печени от 74 г в IV группе до 78 г — во П. Однако следует отметить, что масса лёгкого и почек была выше у гусят опытных групп. Если в I группе масса данных органов была на уровне 35 г, то во II группе была на 2 г меньше (Р< 0,001), а в III и IV группах на 2 и 3 г больше (Р< 0,001).
Питательные и вкусовые качества мяса гусят в значительной степени зависят от химического состава и прежде всего от сухого вещества и содержания в нём протеина и жира. В свою очередь от количества в мясе протеина и жира во многом зависит его энергетическая ценность.
Химический состав мяса гусят и его энергетическая ценность, представлены в таблице 20.
20. Химический состав мяса гусят, % ( n =5)
|
Показатель |
Группа |
|||
|
I |
II |
III |
IV |
|
|
Общая вода |
76,06± 1,01 |
74,33 ±1,32 |
73,35 ±1,15 |
74,39± 1,35 |
|
Сухое вещество |
23,94 ±1,01 |
25,67±1,32 |
26,65 ±1,15 |
25,61±1,35 |
|
Протеин |
18,11±0,45 |
18,60 ±0,56 |
20,11 ±0,77* |
19,12±0,31 |
|
Жир |
5,22±0,17 |
6,46± 0,22** |
5,80±0,11* |
5,58±0,14 |
|
Зола |
0,61±0,01 |
0,61± 0,03 |
0,74± 0,05* |
0,91±0,04*** |
|
Энергетическая ценность 1 ООг мяса, ккал кДж |
122,8 514,0 |
136,3 570,7 |
140,4 587,8 |
130,3 545,4 |
Анализируя полученные данные, можно сказать, что в мясе гусят, получавших с кормом глауконит, просматривается тенденция к снижению влагоёмкости. Если в I группе содержание общей воды составило 76,06%, то во II группе оно снизилось на 1,73%, в III - на 2,71 и в IV группе- на 1,67%. Одновременно на аналогичную величину в данных группах увеличилось содержание в мясе сухого вещества. Причём, в органической части сухого вещества мяса наблюдаются существенные изменения в количественном содержании протеина и жира. Так, в I группе гусят содержание протеина в мясе составило 18,11%, во II группе оно повысилось до 18,60 %, или на 0,49%, в III группе количество протеина в мясе составило 20,11%, или на 2,0% выше чем в I группе (Р< 0,05), а в мясе гусят IV группы данный показатель был на уровне 19,12%, что на 1,01% превысило аналогов I группы.
Содержание жира в мясе гусят, получавших испытуемые дозировки глауконита, превосходило контрольную группу, выращенную на одном полнорационном комбикорме. В результате чего среднее содержание жира в мясе гусят II группы составило 6,46%, III - 5,80 и в IV группе - 5,58%, что было выше содержания жира в мясе гусят I группы соответственно на 1,24%(Р< 0,01), 0,58%(Р< 0,05) и на 0,36%.
Неорганическая часть сухого вещества мяса гусят, представленная минеральными элементами, особенно отличалась в двух последних опытных группах (III и IV). Если в I и II группах содержание сырой золы в мясе было на уровне 0,61%, то в III группе её количество повысилось до 0,74% (Р< 0,05), а в IV группе - до 0,91%(Р< 0,001), то есть на 0,13 и 0,30%.
Расчёт энергетической ценности мяса, проведённый на основании калорийности протеина и жира свидетельствует, что в 100 г мяса гусят I группы общее содержание энергии составило 122,8 ккал, или 514,0 кДж, то в опытных группах его энергетическая ценность была выше и составила: во II группе 136,3 ккал и 570,7кДж, в III группе соответственно 140,4 и 587,8, а в IV группе - 130,3 ккал и 545,4 кДж.
Следовательно, кормовая добавка глауконит в рационе гусят опытных групп за счёт повышения переваримости питательных веществ корма и их лучшему использованию оказывает положительное влияние на развитие мышечной и жировой ткани в организме, большему отложению в мясе протеина и жира, а также увеличивает его энергетическую ценность.
Учитывая, что гусята в течение всего периода выращивания и откорма получали полнорационный комбикорм, сбалансированный за счёт БВМД всеми нормируемыми биологически активными макро - и микроэлементами, определение их содержания в мышечной ткани, костной ткани и печени представляет определённый интерес.
В таблице 21 приводится содержание отдельных макро - и микроэлементов в изучаемых тканях и печени.
Полученные результаты исследований свидетельствуют, что в мышечной ткани гусят-бройлеров кормовая добавка глауконит снижает концентрацию меди в ткани. Если в I контрольной группе содержание меди в мышечной ткани составляет 15,6 мг/кг, то во II группе её количество уменьшается до 6,34 мг/кг, или в 5,3 раза, и в IV группе - до 2,82 мг/кг, или в 5,5 раза (Р< 0,05). И, в то же время, количество кобальта повышается с 1,22 мг/кг в I группе и до 1,86 мг/кг - во II и в III группах (Р< 0,05), до 2,42 мг/кг -в IV группе (Р< 0,05), т.е.практически в 1,5-2,0 раза. Содержание цинка в мышечной ткани гусят опытных групп, по сравнению с I контрольной группой, увеличилось в 2,6-3,0 раза. При этом самое высокое содержание цинка (26,04 мг/кг ) наблюдалось во II группе и с повышением дозировки глауконита снижалось до 24,04 мг/кг в III и 23,12мг/кг - в IV группе. Изменения в количественном содержании железа в мышечной ткани имеет аналогичную закономерность; с 28,44 мг/кг у гусят I группы увеличивается до 44,74 мг/кг во II группе (Р< 0,05), до 39,04 - в III группе(Р< 0,05) и до 32,04 мг/кг - в IV группе (Р< 0,05).
Самое высокое содержание марганца в мышечной ткани было отмечено в мышечной ткани гусят III группы - 0,48 мг/кг, что на 0,40 мг/кг превосходило I группу. Низкая дозировка глауконита в рационах гусят II группы обеспечила практически такую же концентрацию марганца, что и в III группе — 0,42 мг/кг, в то время как самая высокая дозировка глауконита у гусят IV группы на половину снизила содержание марганца в мышечной ткани, чем во II и III группах.
21.Содержание отдельных макро- и микроэлементов в мышечной и костной тканях, печени гусят, мг/кг (X±m>x>, n =5)
|
Показатель |
Группа |
|||
|
I |
II |
III |
IV |
|
|
Мышечная ткань: |
||||
|
Медь |
15,6±2,31 |
6,34± 3,30* |
2,94± 4,15* |
2,82± 3,75* |
|
Кобальт |
1,22±0,16 |
1,86±0,18* |
1,86 ±0,14* |
2,42 ±0,21** |
|
Цинк |
8,74± 1,25 |
26,04±2,16*** |
24,04 ±3,18** |
23, 12 ±3,55** |
|
Железо |
28,44±ЗД5 |
44,74±4,17* |
39,04 ±3,00* |
32,04± 2,99 |
|
Марганец |
0,08 ±0,02 |
0,42 ±0,1 7 |
0,48 ±0,15* |
0,20±0,09 |
|
Магний |
17,40± 1,04 |
22,12 ±1,13* |
22,06 ±1,35* |
21,66±2,06 |
|
Костная ткань: |
||||
|
Медь |
4,88±0,17 |
2,82±0,55** |
3,64± 0,66 |
2,60± 0,98* |
|
Кобальт |
1,20 ±0,01 |
1,26± 0,01** |
1,04±0,01*** |
1,10±0,01*** |
|
Цинк |
46,80 ±1,19 |
48,60 ±1,32 |
37,7 ±1,56** |
29,00±2,04*** |
|
Железо |
9,70 ±1,09 |
12,80± 0,98* |
7,68± 0,75 |
9,28± 0,88 |
|
Марганец |
0,52± 0,03 |
0,40±0.01* |
0,42± 0,04 |
0,52± 0,06 |
|
Магний |
22,74± 0,89 |
24,28 ±0,53 |
20,00± 1,01 |
22,24± 0,88 |
|
Печень: |
||||
|
Медь |
98,00± 3,03 |
89,66 ±4,1 5 |
126,00±5,07*** |
93,36±1,18 |
|
Кобальт |
1,34±0,11 |
1,34±0,10 |
1,40±0,13 |
2,66±0,18*** |
|
Цинк |
13,72 ±1,78 |
20,62 ±1,56* |
20,72 ±1,75* |
20,94 ±2,09* |
|
Железо |
44,68±5,16 |
60,64±4,18* |
60,72 ±3,85* |
53,00± 2,01 |
|
Марганец |
0,52± 0,03 |
0,80 ±0,09* |
0,76±0,13 |
0,58 ±0,09 |
|
Магний |
19,12±0,43 |
20,76±0,15** |
10,74±0,66*** |
10,98±0,39*** |
Магний мышечной ткани находится в осмотически не деятельном состоянии, т.е. относительно прочном соединении с белками. Он поровну распределён между актином и миозином. Соединённые с белками ионы магния притягивают к себе АТФ, вызывая пространственную деформацию её молекул и тем самым обеспечивают активное воздействие на неё АТФ-центра молекулы миозина. По сравнению с I контрольной группой гусят, в мышечной ткани которых содержание магния составляло 17,40 мг/кг, в опытных группах его количество увеличилось от 21,66 мг/кг в IV группе до 22,13 мг/кг - во II группе.
Прочность костной ткани скелета, его высокие механические качества обусловлены большой концентрацией минеральных солей, стабилизирующих органическую основу костной ткани. Минеральные соли, депонируемые в скелете, не являются инертным балластом - они принимают активное участие в поддержании гомеостаза макро- и микроэлементов, а также ряда катионов в организме животного.
Имеющиеся результаты анализов костной ткани показывают, что глауконит в рационе птицы приводит к снижению содержания в ней меди. Если в I группе её количество было на уровне 4,88 мг/кг, то во II группе она уменьшилась до 2,82 мг/кг, в III - до 3,64 и в IV группе - до 2,60 мг/кг или соответственно на 2,6 мг/кг (Р< 0,01), 1,24 и 2,28 мг/кг ( Р< 0,05).
Низкая дозировка глауконита во II группе гусят по сравнению с I повысила содержание кобальта в костной ткани до 1,26 мг/кг, или на 5,0%(Р< 0,01), а средняя и высокая дозировки ( III и IV группа) уменьшили его содержание до 1,04 - 1,10 мг/кг (Р< 0,001). Аналогичная картина наблюдается и по цинку; его количество в костной ткани снижается с 46,80 мг/кг в I группе до 37,70 в III и до 29,00 мг/кг - в IV группе (Р< 0,001).
Самое высокое содержание железа в костной ткани наблюдалось во II группе - 12,80 мг/кг, что превосходило I группу на 3,10 мг/кг (Р< 0,05), а в III и в IV группе его количество снизилось до 7,68 и 9,28 мг/кг. Печень является одним из важных паренхиматозных органов живого организма, в котором присутствуют практически все минеральные элементы.
Так, печень является основным депо лабильной меди в организме и концентрация в ней этого элемента служит, по мнению В.И.Георгиевского и др.(1979), индикатором усвоения меди из корма и обеспеченности им организма животных. Полученные данные свидетельствуют, что самая высокая концентрация меди в печени наблюдалась у гусят III группы -126,00 мг/кг и превосходило I контрольную группу на 28,00 мг/кг (Р< 0,001), в то время как во II и IV группах её количество было на уровне соответственно 89,66 и 93,36 мг/кг.
Основное депонирование железа в печени происходит в форме ферритина. По мере расхода железа плазмы для синтеза гемоглобина, миоглобина и ферментов железо из депо поступает в плазму. Если в печени гусят I группы железа содержалось 44,68 мг/кг, то во II группе оно повысилось до 60,64 мг/кг, и ли на 35,7%(Р< 0,05), в III -до 60,72, или на 35,9% (Р< 0,05), а в IV группе - до 53,00 мг/кг, или на 18,6%(Р> 0,05).
В печени в обычных условиях, примерно, половина всего кобальта приходится на витамин Bi2, причём соотношение кобаламиновых и не кобаламиновых соединений варьирует в зависимости от обеспеченности животных кобальтом (при его недостатке практически весь кобальт печени содержится в форме витамина). Нарастающее содержание кобальта в печени гусят наблюдается только при средней и высокой дозировке глауконита в рационе. Так, в I и во II группах среднее содержание кобальта в печени было одинаковым - 1,34 мг/кг, в III группе оно увеличилось до 1,40 мг/кг, или 4,5%(Р> 0,05), а в IV группе - до 2,66 мг/кг, или на 98,5% (Р< 0,001).
В организме нет тканей с преимущественной локализацией цинка; элемент распределяется в них довольно равномерно. Тем не менее, наиболее насыщены цинком костная ткань, печень, кожа и перья. В качестве структурного компонента цинк входит в молекулы карбоангидразы, панкреатической карбоксипептидазы, дегидрогеназ глутаминовой и молочной кислот. В наших исследованиях все изучаемые дозировки глауконита повысили содержание цинка в печени гусят с 13,72 мг/кг в I группе до 20,62 мг/кг во II группе , 20,72 - в III и до 20,94 мг/кг - в IV группе (Р< 0,05), что соответственно составило50,3%, 51,0 и 52,6%.
В печени на долю марганца приходится около 17-18% общего содержания марганца в организме. Он принимает активное участие в окислительно-восстановительных процессах, тканевом дыхании, костеобразовании, оказывает влияние на рост, размножение, кроветворение, функцию эндокринных органов. Характерно отметить, что только в печени гусят II группы низкая дозировка глауконита способствовала повышению его концентрации до 0,80 мг/кг, что превосходило I группу на 0,28 мг/кг (Р< 0,05) и с повышением дозировки глауконита в III и IV группах наблюдается снижение его содержания соответственно до 0,76 и 0,58 мг/кг.
Содержание магния в печени гусят-бройлеров имело существенное различие. Если в I группе содержание магния было на уровне 19,12 мг/кг, то во II группе отмечено его повышение до 20,76 мг/кг, или на 1,64 мг/кг (Р< 0,01), в то время как в III группе его количество уменьшилось до 10,74 мг/кг, а в IV группе -до 10,98 мг/кг, т.е. в 1,7-1,8 раза (Р< 0,001).
Следовательно, глауконит в рационах гусят-бройлеров оказывает определённое влияние на кумуляцию в печени основных биогенных микроэлементов, причём в большей степени это наблюдается в III группе, гусята которой получали дозировку глауконита 0,2% от сухого вещества рациона.
3.5 Конверсия протеина и энергии корма в питательные вещества мясной продукции
Для нормальной жизнедеятельности и продуктивности сельскохозяйственных животных и птицы необходимо постоянное поступление в организм энергии, источником её являются все органические вещества корма.
На эффективность использования валовой энергии корма в организме оказывают влияние кормовые добавки природного и синтетического происхождения, биологически активные вещества.
22. Поступление и использование энергии корма гусятами, МДж/гол. в сутки
|
Показатель |
Группа |
|||
|
I |
II |
III |
IV |
|
|
Валовая энергия Переваримая энергия Обменная энергия, % Обменность энергии |
5,40 4,02 3,81 70,6 |
5,40 4,12 3,91 72,4 |
5,40 4,18 3,98 73,7 |
5,40 4,15 3,95 73,2 |
Добавка глауконита в рацион гусят-бройлеров оказала определённое влияние на использование энергии корма. Проведённый в таблице 22 косвенный расчёт использования энергии корма гусятами показал, что при одинаковом поступлении с кормом во всех группах - 5,4 МДж на голову в сутки позволило получить различия в переваримости органической части корма у гусят опытных групп под влиянием глауконита. Так, если в I группе переваримая энергия составила 4,02 МДж, то во II она увеличилась до 4,12 МДж, или на 2,5%, в III - до 4,18, или на 4,0% и в IV - группе - до 4,15 МДж, или на 3,2%.
Количество обменной или физиологически полезной энергии, которая идёт в организме на поддержание жизнедеятельности и образование продукции, было также выше у гусят опытных групп. В рационе гусят I контрольной группы количество обменной энергии было на уровне 3,81 МДж, в то время как во II группе 3,91 МДж, в III - 3,98 и в IV группе - 3,95 МДж, что в расчёте от валовой энергии соответственно составило 70,6%, 72,4, 73,7, 73,2%.
Определение показателей мясной продуктивности гусят - бройлеров и расчёт затрат корма на единицу произведённой продукции не дают объективной оценки эффективности конверсии птицей питательных веществ
корма в продукцию и не характеризуют его способности к максимальному производству пищевого белка.
Предложенный Л.К. Лепайые и др. (1982) методы определения эффективности конверсии животными и птицей питательных веществ корма в питательные вещества мясной продукции, то есть по выходу пищевого белка и жира, во многом подтвердили результаты контрольного убоя гусей.
Показателями оценки мясной продуктивности гусят - бройлеров являются коэффициенты конверсии кормового протеина в белок ткани организма (ККП) и коэффициент конверсии энергии в энергию отложения белка и жира (ККЭ). Полученные данные по конверсии кормового протеина и энергии в продукцию представлены в таблице 23.
Проведённый расчёт показывает, что у гусят в убойном возрасте, то есть при достижении ими 60-дневного возраста среднесуточное отложение в тканях тела белка и жира составило: в I контрольной группе 414,4 г белка и 322,4 г жира, в опытных группах по сравнению с I отложение данных питательных веществ было выше соответственно во II группе на 38,4 и 64,6 г, вIII-на 94,5 и 44,5 г, в IV группе на 61,5 и 37,8 г.
23. Трансформация протеина и энергии корма в продукцию (в среднем на голову, г)
|
Показатель |
Группа |
|||
|
I |
II |
III |
IV |
|
|
Предубойная живая масса |
3854 |
4072 |
4300 |
4146 |
|
Отложилось в тканях тела: |
||||
|
белка |
414,4 |
452,8 |
508,9 |
475,9 |
|
жира |
322,4 |
387 |
366,9 |
360,2 |
|
Выход на 1 кг живой массы: |
||||
|
белка |
107,5 |
111,2 |
118,4 |
114,8 |
|
жира |
83,7 |
95,0 |
85,3 |
86,9 |
|
ККП, % |
15,8 |
17,2 |
19,3 |
18,1 |
|
ККЭ, % |
12,8 |
14,7 |
15,0 |
14,4 |
В расчёте на 1 кг живой массы выход белка и жира, отложенного в тканях тела, также различался. Если у гусят I группы, получавших за период
откорма один только полнорационный комбикорм в среднем за сутки в расчёте на 1 кг живой массы откладывалось в теле 107,5 г белка и 83,7 г жира, то добавка глауконитового концентрата к рациону гусят II-IV опытных групп по сравнению с I повысила выход белка во II группе на 3,7 г, в III на 10,9 и в IV группе - на 7,3 г, жира соответственно на 11,3г, 1,6 и 3,2 г.
Самый высокий коэффициент конверсии протеина 19,3% наблюдался в III группе, затем в IV (18,1%), во II (17,2%) и в последнюю очередь в I группе - 15,8%. Аналогичная закономерность просматривается и при расчёте коэффициента конверсии энергии в энергию отложения белка и жира. В I группе ККЭ составил 12,8%, во II -14,7, в III - 15,0 и в IV группе - 14,4%.
Следовательно, из всех изучаемых дозировок глауконита самая высокая трансформация протеина и энергии корма в продукцию наблюдается при добавлении глауконита в количестве 0,2% от сухого вещества рациона.
3.6 Затраты корма и экономическая эффективность проведённых исследований
В работе любого сельскохозяйственного предприятия важным условием является цена реализации произведённой продукции, которая во многом зависит от себестоимости. В структуре себестоимости наибольший удельный вес занимают корма и заработная плата. Поэтому, одним из важных экономических показателей, характеризующих эффективность ведения отрасли птицеводства, является расчёт затрат кормов на получение единицы продукции. Чем ниже будут затраты корма на получение продукции, тем ниже будет её себестоимость. А удешевление производства продукции птицеводства является одним из путей интенсивного ведения отрасли.
Проведённый нами расчёт фактических затрат кормов за период опыта по выращиванию и откорму гусят-бройлеров с использованием кормовой добавки глауконит представлен в таблице 24.
Полученные данные в таблице 24 показывают, что при одинаковом расходе корма в расчёте на 1 голову во всех группах подопытных гусят общее количество скормленного корма и содержащихся в нём питательных веществ между группами существенно различалось. Основная причина данного различия - это разная сохранность поголовья гусят в группах. Если в I группе за весь период опыта было скормлено 6659,23 кг корма, то во II оно превосходило на 182,63 кг, в III - на 295,03 и в IV группе - на 252,88 кг. В опытных группах, по сравнению с I контрольной, гусятам было скормлено больше: обменной энергии на 2257 - 3646 МДж, сырого протеина - на 33,6 -54,4 кг.
24. Затраты кормов на получение одного кг прироста живой массы гусят (в среднем по группе)
|
Показатель |
Группа |
|||
|
I |
II |
III |
IV |
|
|
Среднее поголовье гусят в группе, голов |
474 |
487 |
495 |
492 |
|
Скормлено корма на 1 голову, г |
14049 |
14049 |
14049 |
14049 |
|
Скормлено всего: кормов, кг |
6659,23 |
6841,86 |
6954,26 |
6912,11 |
|
обменной энергии, МДж |
82286 |
84543 |
85932 |
85411 |
|
сырого протеина, кг |
1227,7 |
1261,3 |
1282,1 |
1274,3 |
|
Прирост живой массы 1 головы, г |
3798,6 |
4002,4 |
4227,3 |
4070,5 |
|
Валовый прирост живой массы по |
||||
|
группе, кг |
1800,54 |
1949,17 |
2092,51 |
2002,69 |
|
Затрачено на 1 кг прироста живой |
||||
|
массы: кормов, кг |
3,70 |
3,51 |
3,32 |
3,45 |
|
в % к I группе |
100 |
94,9 |
89,7 |
93,2 |
|
обменной энергии, МДж |
45,7 |
43,4 |
41Д |
42,6 |
|
в % к I группе |
100 |
95,0 |
89,9 |
93,2 |
|
сырого протеина, г |
682 |
647 |
613 |
636 |
|
в % к I группе |
100 |
94,9 |
89,9 |
93,3 |
Прирост живой массы 1 гусёнка за опыт позволил рассчитать валовой прирост живой массы гусят в целом по группе. В результате чего в I группе валовой прирост живой массы гусят составил 1800,54 кг, во II - 1949,17, в III - 2092,51 и в IV группе - 2002,69 кг. Рассчитанное количество скормленных кормов, питательных веществ и валовой прирост живой массы гусят позволили провести расчёт затрат на произведённую единицу продукции. Так, при откорме гусят на одном полнорационном комбикорме (I группа) на получение 1 кг прироста живой массы затрачивается 3,7 кг корма, с добавкой низкой дозировки глауконита (II группа) - 3,51, средней (III группа)- 3,32 и с высокой дозой глауконита (IV группа) - 3,45 кг корма, что в относительном выражении соответственно составляет- 100%, 94,9, 89,7 и 93,2%.
Затраты обменной энергии в I группе гусят были на уровне 45,7 МДж и снизились в опытных группах; на 2,3 МДж во II группе, на 4,6 - в III и на 3,1 МДж - в IV группе. Аналогичная тенденция просматривается и по сырому протеину, затраты которого на 1 кг прироста живой массы составили в I группе 682 г, в то время как во II группе они уменьшились до 647 г, в III -до 613 и в IV группе - до 636 г, что по сравнению с I группой данная разница составила соответственно 35 г, 69 и 46 г.
Интенсивный путь развития любой отрасли животноводства, и в том числе птицеводства, предусматривает внедрение в производство новых научных разработок, позволяющих сельскохозяйственному предприятию при определённых дополнительных затратах получать дополнительную продукцию, превышающую стоимость вложенных денежных средств, то есть работать рентабельно.
В наших исследованиях изучалось влияние природного алюмосиликата глауконита на рост и развитие гусят-бройлеров, переваримость и использование питательных веществ корма, показаний контрольного убоя и проведён расчёт экономической эффективности, подтверждающий целесообразность проведения данных мероприятий (таблица 25).
25.Экономическая оценка результатов опыта
|
Показатель |
Группа |
|||
|
I |
II |
III |
IV |
|
|
Валовой прирост живой массы |
||||
|
гусят, кг |
1800,54 |
1949,17 |
2092,51 |
2002,69 |
|
Скормлено кормов всего, кг |
6659,23 |
6841,86 |
6954,26 |
6912,11 |
|
Скормлено глауконита, г |
- |
5772 |
11733 |
17494 |
|
Стоимость скормленных |
||||
|
кормов, руб. |
46615 |
47893 |
48680 |
48385 |
|
Стоимость глауконита, руб. |
- |
57,7 |
117,3 |
174,9 |
|
Общая стоимость кормов и глауконита, руб. |
46615 |
47950,7 |
48797,3 |
48559,3 |
|
Стоимость 1кг прироста живой массы, руб. |
25,89 |
24,60 |
23,32 |
24,25 |
|
Произведено прироста живой |
||||
|
массы, кг: |
||||
|
на каждые скормленные 1 00 |
||||
|
кг корма |
27,04 |
28,49 |
30,09 |
28,97 |
|
в % к I группе |
100 |
105,4 |
111,3 |
107,1 |
|
на каждые скормленные 1 00 |
||||
|
руб. корма |
3,86 |
4,06 |
4,29 |
4,12 |
|
- в % к I группе |
100 |
105,2 |
111,1 |
106,7 |
|
Получено дополнительно прироста живой массы гусят, кг |
. |
148,63 |
291,97 |
202,15 |
|
Стоимость прироста живой массы, руб. |
_ |
8323,3 |
16350,3 |
11320,4 |
Добавка глауконита к основному рациону гусят-бройлеров в количестве 5772 г во II группе, 11733 г в III и 17494 г - в IV группе привело к удорожанию стоимости скормленных кормов на сумму соответственно 57,7;
117,3 и 174,9 руб. В результате чего в I контрольной группе общая стоимость кормов за весь период опыта составила 46615 руб., а в опытных группах -47950,7 руб. во II группе, 48797,3 руб.- в III и 48559,3 руб. - в IV группе.
Однако, не смотря на это, более высокие среднесуточные приросты у гусят-бройлеров в опытных группах за счёт кормовой добавки глауконит оказали определённое влияние на стоимость единицы прироста живой массы. Если в I группе стоимость 1 кг прироста живой массы была равна 25,89 руб., то во II группе она снизилась до 24,60 руб., или на 5%, в III группе - до 23,32 руб., или на 9,9%, и в IV группе - до 24,25 руб., или на 6,3%.
Оплата корма продукцией является одним из экономических показателей, характеризующим производство продукции на единицу вложенных натуральных и стоимостных средств. При этом в натуральном выражении, то есть на каждые скормленные 100 кг корма в I группе было получено 27,04 кг прироста живой массы, в то время как во II группе - 28,49 кг, в III - 30,09 и в IV группе - 28,97 кг, что по сравнению с I группой было выше соответственно на 5,4%, 11,3 и 7,1%. В стоимостном выражении, то есть в расчёте на каждые скормленные 100 руб. корма, в I группе было произведено 3,86 кг прироста живой массы, во II группе - 4,06 кг, или на 5,2% больше, в III - 4,29 кг, или 11,1%, в IV группе - 4,12 кг, или 6,7%.
Полученные различия в валовом приросте живой массы в опытных группах, по сравнению с контрольной, за счёт скармливания кормовой добавки глауконит даёт возможность получить дополнительный прирост живой массы во II группе 148,63 кг, в III - 291.97 ив IV группе - 202,15 кг, что в стоимостном выражении соответственно составило 8323,3 руб., 16350,3 и 11320,4 руб.
Следовательно, с экономической точки зрения наиболее выгодной в рационах гусят-бройлеров является кормовая добавка глауконит в количестве 0,2% от сухого вещества рациона.
4. Результаты производственной проверки
Испытание новых кормовых добавок на небольшом поголовье сельскохозяйственных животных и птицы при проведении научных, а также научно-хозяйственных опытов требует обязательной апробации полученных результатов на большом поголовье, то есть требуется постановка и проведение производственного опыта.
Вышеприведенные результаты научного и научно-хозяйственного опытов по определению оптимальной дозировки кормовой добавки глауконитового концентрата в рационе гусят-бройлеров показали, что наиболее целесообразной является доза глауконита 0,2% от сухого вещества.
26. Результаты производственной проверки
|
Показатель |
Группа |
|
|
I |
II |
|
|
Среднее поголовье гусят: |
||
|
в начале опыта |
950 |
975 |
|
в конце опыта |
884 |
951 |
|
Сохранность, % |
93,1 |
97,5 |
|
Средняя живая масса 1 головы, г: |
||
|
в начале опыта |
96,5 |
96,4 |
|
в конце опыта |
3821,0 |
4184,9 |
|
Прирост живой массы, г |
3724,5 |
4088,5 |
|
Среднесуточный прирост, г |
57,3 |
62,9 |
|
Валовой прирост живой массы, кг |
3292,46 |
3888,16 |
|
Скормлено кормов, всего, кг |
12429,92 |
13372,01 |
|
Скормлено глауконита, кг |
- |
22,73 |
|
Затраты корма на 1 кг прироста живой массы, кг |
3,78 |
3,44 |
|
Получено дополнительного прироста живой массы, |
||
|
кг |
- |
595,7 |
|
Стоимость прироста живой массы, руб. |
- |
31870,0 |
В I контрольной группе за период опыта их 950 гусят на убой было отправлено 884 головы. При этом сохранность поголовья составила 93,1%. Во II опытной группе из 975 голов к концу опыта осталось 951 голова, то есть сохранность поголовья составила 97,5%.
Добавка глауконита в рацион гусят опытной группы позволила за период откорма получить прирост живой массы 4088,5 г, или на 364 г больше, чем в контрольной группе. Данное различие объясняется среднесуточным приростом, который в опытной группе составил 662,9 г, что ; на 9,8% выше чем в контрольной группе. В результате чего за период опыта в контрольной группе был получен валовой прирост живой массы 3292,46 кг, в то время как в опытной группе - 3888,16 кг, или на 595,7 кг больше.
На основании фактического количества скормленных кормов и полученного прироста живой массы был проведен расчет затрат кормов на единицу прироста живой массы. Если в контрольной группе на 1 кг прироста живой массы было затрачено 3,78 кг комбикорма, то в опытной группе они уменьшились на 3,44 кг, или на 9,0%.
Кормовая добавка глауконит в рационе гусят-бройлеров по сравнению с группой, получавшей только один комбикорм, позволила получить дополнительно прироста живой массы 595,7 кг на общую сумму 31,87 тыс. руб.
Таким образом, производственная апробация подтвердила результаты научно-хозяйственного опыта о целесообразности использования в рационах гусят-бройлеров кормовой добавки глауконит в оптимальной дозировке -0,2% от сухого вещества рациона.
5. Обсуждение результатов собственных исследований
Одним из путей повышения продуктивности сельскохозяйственной птицы является использование в их рационе кормовой добавки природных алюмосиликатов. Почти сорок лет учёными разных стран мира отрабатываются нормы ввода природных алюмосиликатов в рационы разных видов, возрастных и породных групп сельскохозяйственной птицы.
Самыми распространёнными как в нашей стране, так и за рубежом природными алюмосиликатами являются цеолиты. Детальное изучение их
физико-химических свойств показало, что они обладают высокими сорбционными, молекулярно-ситовыми и ионообменными свойствами. Именно эти качества цеолитов лежат в основе их биологического действия на живой организм. За счёт локализации в полостях и каналах каркаса цеолита катионов натрия, калия, кальция, магния и других элементов, цеолита изменяет ионный состав химуса, нормализуя таким образом рН и оптимизируя условия деятельности пищеварительных ферментов (протеолитических и амилолитических). В результате чего переваримость органического вещества повышается в среднем на 4,0%, сырого протеина на 3-4, сырого жира на 3,0, сырой клетчатки — на 10-15%, усвоение азота на 2-3%. Как следствие, яйценоскость птицы увеличивается на 7-8%, сохранность поголовья - на 2-6%, расход корма снижается на 17-21%. У цыплят-бройлеров при добавлении им в корм цеолитов среднесуточный прирост возрастает на 6,5%, а затрата корма уменьшается на 13%.
Многие учёные (Фисинин В., Шадрин A.M., Донченко О.А., Mumptjn F., Fishman P. и др.) считают оптимальной нормой ввода природных цеолитов в рацион сельскохозяйственной птицы от 3 до 6% от массы комбикорма.
К группе природных алюмосиликатов относится и глауконит, который в отличие от цеолита имеет не каркасное, а слоистое строение. В результате чего активная поверхность алюмосиликата увеличивается в 5 и более раз. Следовательно, и биологическое действие глауконита на много выше, чем у цеолита. Это подтверждают научные работы Е.В. Иванова, Г.А. Джинджихадзе, И.Р. Мазгарова, в которых установлена оптимальная норма ввода глауконита в рацион свиней - не более 0,25% от сухого вещества рациона.
Исследования, произведённые А.А. Замятиным на курах несушках, подтвердили оптимальную дозировку глауконита для птицы- 0,25% от сухого вещества рациона. При этом переваримость органического вещества корма
увеличивалась на 2,0 - 3,4%, сырого протеина - на 7,6-7,8, сырого жира 18,9-19,9, сырой клетчатки - на 10,8%, использование азота корма - на 8-9%.
Однако разные виды сельскохозяйственной птицы отличаются друг от друга не только в анатомическом строении органов пищеварения, но и в функциональной активности желёз внутренней секреции. Так, у гусей в отличие от кур, количество панкреотического сока во время кормления увеличивается в 2,0 - 2,2 раза, а между кормлением его выделяется меньше чем у кур. В то же время амилазы у кур намного выше, чем у гусей. В мышечном желудке у гусей корм подвергается более основательной механической обработке, чем у других видов птицы. Длина тонкого отдела кишечника у гусей в среднем на 100 см, а слепой кишки на 14 см длиннее аналогичных отделов кишечника у кур. Всё это накладывает свой отпечаток на переваримость и использование питательных веществ в корме.
Поэтому целью нашей работы явилось изучение влияния глауконита на продуктивные качества гусят-бройлеров. При этом ставились вопросы определить оптимальную дозировку кормовой добавки глауконита в рационах гусят, установить её влияние на переваримость и использование основных питательных веществ корма, конверсию энергии и протеина в продукцию, рост и сохранность поголовья, показатели мясной продуктивности с экономическим основанием проведённых исследований.
Для чего в условиях АОЗТ "Птицевод" Октябрьского района Челябинской области в период 2001-2002 годов проведён научный, научно-хозяйственный и научно-производственный опыты на гусятах-бройлерах. Четыре группы гусят в научно-хозяйственном опыте продолжавшемся 65 дней получали одинаковый рацион кормления, в 100 г которого в первые 20 дней содержалось 1,3 МДж обменной энергии, 21,6 г сырого протеина, 5,1 г сырого жира, 4,2 г сырой клетчатки, 1,2 мг лизина и 0,9 мг метионина с цистином, 2,3 г линолевой кислоты. Во второй период откорма (21-66 дней) концентрация питательных веществ в 100 г комбикорма составила: 1,26 МДж
обменной энергии, 18,6 сырого протеина, 4,5 г сырого жира, 6,15 г сырой клетчатки, 1,9 г линолевой кислоты, 0,96 мг лизина и 0,70 мг метионина с цистином. ЭПО по периодам откорма составило 144,5 и 162 ккал в расчёте на 1% сырого протеина. На фоне основного рациона гусятам опытных групп ежедневно добавляли концентрат глауконита в количестве 0,1%, 0,2 и 0,3% от сухого вещества. В результате чего, при среднем расходе комбикорма на одного гусёнка за период опыта 14,049 кг в опытных группах скормлено 11,78 г, 23,55 и 35,33г глауконита.
Выращивание гусят-бройлеров на полнорационном комбикорме, соответствующего детализированной системе нормализованного кормления сельскохозяйственной птицы, позволяет за 65 дней при среднесуточном приросте 59,9 г получить абсолютный прирост живой массы 3894,5 г. При этом переваримость питательных веществ рациона была на уровне: сухого вещества 75,8%, органического вещества -77,8, протеина -88,6, жира-52,2, клетчатки-29,2 и БЭВ-81,9%. В теле гусят среднесуточное отложение азота равнялось 3,40 г, кальция -2,43 г, фосфора - 1,12г, что соответственно составило 35,1%, 56,6 и 51,6% от принятого с кормом.
При изучении морфологических и отдельных биохимических показателей крови отклонений от нормы не было. Содержание отдельных микроэлементов в крови зависело от их количества в витаминно-минеральном премиксе комбикорма.
Проведённый контрольный убой гусят по завершению научно-хозяйственного опыта показал, что убойный выход потрошенной тушки был на уровне 73,2%, выход съедобных частей составил 68,2%, в результате чего индекс мясных качеств был равен 2,15. В потрошённой тушке на долю мышечной ткани пришлось 54,7%, внутреннего жира- 2,8, кожи с подкожным жиром - 23,0, костей - 19,5%. Энергетическая ценность ЮОг мяса, рассчитанная на основании химического состава, составила 122,8 ккал, или514,ОМДж.
При расчете трансформации протеина и энергии корма в продукцию количество обменной энергии от валовой энергии корма составило 70,6%. Выход на 1 кг живой массы белка составил 107,5 г, жира -83,7 г. В результате чего коэффициент конверсии протеина в продукцию (КПП) был на уровне 15,8%, коэффициент конверсии энергии в продукцию (ККЭ)-12,8%.
Откорм гусят на полноценном комбикорме позволил получить на 1 кг прироста живой массы затраты кормов 3,70 кг, обменной энергии - 45,7 МДж, сырого протеина - 682 г, а в стоимостном выражении - 25,89 руб. При ] этом оплата корма продукцией в расчёте на каждые скормленные 100 кг J корма и 100 рублей истраченные на корма составила соответственно 27,04 и 3,86 руб.
Испытание низкой дозировки глауконита в количестве 0,1% от сухого вещества основного рациона позволило за период откорма при среднесуточном приросте 63,0 г получить абсолютный прирост живой массы 4098,5 г, что на 204 г, или на 5,2% было выше по сравнению с гусятами, получавшими только один полнорационный комбикорм. При этом сохранность поголовья в группе возросла до 97,4%, в то время как в контрольной группе она была на уровне 94,8%.
Увеличение прироста живой массы гусят, получавших низкую дозировку глауконита, объясняется тем, что она в организме птицы способствует повышению переваримости сырого протеина на 2,4%, а сырого жира — на 12,0%. В результате чего в теле гусят среднесуточное отложение азота возросло на 0,11 г и составило 3,51 г, против 3,40 г в группе, не получавшей глауконит. При этом в сыворотке крови гусят намечалась тенденция к повышению уровня общего белка, хотя количество аминного азота не изменилось. Различия в переваримости сырого жира привело к тому,
что в сыворотке крови содержание общих липидов возросло на 0,6 г/л, а из определяемых основных биогенных микроэлементов достоверно повысилось количество цинка на 1,56 мг/л, или на 26,1%. Учитывая, что цинк входит в состав секретируемых ферментов поджелудочной железы, по всей вероятности, произошло повышение активности липолитических ферментов повлиявших на переваримость сырого жира.
Различия в живой массе гусят, получавших низкую дозировку глауконита, привели к тому, что в их тушках при убойном выходе 73,4%, выход съедобных частей составил 2508 г, или 69,1% от массы не потрошённой тушки, что на 0,9% больше, чем в тушках гусят не получавших глауконит. Увеличение съедобных частей тушек произошло в основном за счёт мышечной ткани на 107 г, или на 0,5%, внутреннего жира - на 29г, или на 0,8% и кожи с подкожным жиром - на 32 г. В мышечной ткани количество жира возросло на 1,24% при одинаковом содержании протеина, что и в мясе гусят не получавших глауконит. В результате чего энергетическая ценность мяса составила 136,3 ккал, или 570,7 МДж, что соответственно на 13,8 ккал и 56,7 МДж было выше, чем мясо гусят контрольной групп
Высокие ионообменные свойства глауконита обеспечили в мышечной ткани гусят, получавших низкую дозировку глауконита, повышение содержания меди, кобальта, цинка, железа и магния, в костной ткани - меди, кобальта, железа и марганца, в печени — цинка, железа, марганца и магния.
Полученная разница в переваримости питательных веществ рациона привела к тому, что в организме гусят количество обменной энергии от валовой возросло до 72,4%, против 70,6% у гусят, выращенных и откормленных на одном полнорационном комбикорме.
Низкая дозировка глауконита в рационах гусят позволило обеспечить выход на 1 кг живой массы белка - 111,2 г, жира 95,0 г при ККП - 17,2%, ККЭ - 14,7%, что соответственно выше гусят контрольной группы на 3,7 и 11,3 г, 1,4 и 1,9%.
Кормовая добавка глауконит в количестве 0,1% от сухого вещества рациона обеспечила снижение в рационе на 1 кг прироста живой массы затраты кормов, обменной энергии и сырого протеина - на 5,0-5,1%. При этом стоимость 1 кг прироста снизилась на 1 руб. 29 коп., а оплата корма продукцией возросла в натуральном и стоимостном выражении на 5,2-5,4%. Дополнительно полученный прирост живой массы в размере 148,63 кг позволил получить прибыль 8323,3 руб.
Добавка к рациону гусят-бройлеров глауконита в количестве 0,2% от сухого вещества рациона позволила получать самый высокий прирост живой массы за период опыта - 4323,5г, что на 11,0% больше по сравнению с гусятами выращенных на одном полноценном комбикорме. При этом среднесуточный прирост живой массы составил 66,5 г.
Высокие сорбционные способности глауконита позволили, особенно в первую декаду выращивания, сократить падёж гусят. В результате чего сохранность поголовья составила 99% против 94,8% в группе не получавшей глауконит.
Кормовая добавка глауконит в дозе 0,2% от сухого вещества рациона обеспечила повышение переваримости органического вещества корма на 2,4%, протеина - на 5,0 и жира - на 8,1%. В результате чего у гусят данной группы в организм поступало самое высокое количество обменной энергии -3,98 МДж в сутки, а обменность энергии составило 73,7%. Глауконит в рационах гусят уменьшил выделение азота с помётом на 0,3 г, повысив тем самым на аналогичную величину его отложение в теле до уровня 3,70 г. Кроме этого в теле гусят баланс кальция на 0,47 г был выше по сравнению с гусятами не получавшими глауконит.
Данная дозировка глауконита повышает окислительно-восстановительные процессы в организме птицы, о чём свидетельствует увеличение эритроцитов в крови на 10,7 %, содержание гемоглобина - на 7,1%. Повышение числа лейкоцитов на 7,0% можно расценивать как повышение защитной функции организма. Наряду с этим наблюдаются изменения в метаболитах белкового и липидного обмена: содержание общего белка в сыворотке крови возросло на 29,9%, аминного азота - на 16,7, (3-липопротеидов - на 32,1%. Из минеральных элементов крови достоверно увеличилось количество кобальта на 30% и наметилась тенденция повышения меди, цинка и железа.
Более высокая трансформация переваренных питательных веществ через кровь в органы и ткани гусят привела к тому, что в расчёте на 1 кг живой массы в среднем за сутки в их теле откладывалось белка 118,4 г, жира - 85,3 г. В результате чего ККП составил 19,3%, ККЭ - 15%, что соответственно на 3,5 и 2,2% было выше по сравнению с гусятами не получавшими глауконит. В результате чего масса кормовая добавка глауконит увеличила массу потрошённой тушки на 352 г, в том числе выход съедобных частей — на 323 г. В основном это произошло за счёт мышечной ткани - на 281 г, внутреннего жира на 13, кожи с подкожным жиром на 21 и костной ткани на 37. При этом в мышечной ткани количество протеина возросло на 2%, жира на 0,58, а минеральных веществ - на 0,13%. В результате чего энергетическая ценность мяса составила 140,4 ккал, или 587,8 кДж. Из минеральных веществ в мышечной ткани наблюдается достоверное повышение меди, цинка, кобальта, железа, марганца и магния, в костной ткани — цинка и снижение кобальта, в печени — повышение меди, цинка и железа, снижение магния.
В конечном итоге добавка глауконита 0,2% от сухого вещества рациона позволила получить самые низкие затраты на 1 кг живой массы: кормов 3,32 кг, обменной энергии-41,1МДж, сырого протеина - 613,а стоимость 1 кг прироста живой массы - 23,32 руб. При этом оплата корма продукцией была самой высокой и составила в натуральном выражении 30,09 руб., а в стоимостном - 4,29 руб. Дополнительно полученный прирост живой массы исчислялся в 16,35 тыс. руб.
Высокая дозировка кормовой добавки глауконита-0,3% от сухого вещества рациона гусят-бройлеров при среднесуточном приросте 64,1г позволила получить абсолютный прирост живой массы за опыт 4166,5г, что на 7%превосходило гусят не получавших глауконит. При этом сохранность поголовья составила 96,4%. Данная дозировка обеспечила повышение переваримости органического вещества на 2,1% сырого протеина - на 3,6 и сырого жира - на 9,7% при отношении обменной энергии к валовой 73,2%. В результате чего с пометом из организма терялось меньше, а в теле откладывалось на 0,16 г азота больше. При одинаковом отложении фосфора в теле гусят, что и при других изучаемых дозировках, баланс кальция был положительным и его откладывалось на 0,39 г больше по сравнению с группой не получавшей глауконит. В результате чего в крови гусят наблюдается тенденция к увеличению отдельных метаболитов белкового и липидного обмена: общего белка - на 11,8%, общих липидов - на 9,9, Р -липопротеидов - на 17,6%. При этом морфологические показатели крови и содержания в ней отдельных макро - и микроэлементов не имели достоверной степени различия.
Всё это привело к тому, что масса потрошённой тушки, при убойном выходе 73,9%, превосходила тушку гусят не получавших глауконит только на 243 г с преобладанием количества съедобных частей на 244 г. В том числе за счёт мышечной ткани на 209 г, внутреннего жира - на 16 г, кожи с подкожным жиром - на 14 г. В мышечной ткани наблюдается достоверное повышение зольности на 0,30%, при этом из микроэлементов за счёт меди, кобальта и цинка. Энергетическая ценность 100 г мяса была ниже, чем в группах гусят получавших глауконит 0,1 и 0,2% от сухого вещества рациона, и составила 130,3 ккал, или 545,4 МДж. В костной ткани данная дозировка глауконита достоверно снизила содержание меди, кобальта и цинка. В печени наблюдается повышение кобальта и цинка, но снижение магния.
Расчёт трансформации протеина и энергии корма в продукцию показал, что на 1 кг живой массы выход белка составил 114,8 г, жира - 86,9 г, при ККП -18,1%, ККЭ - 14,4%. Это превосходило расчётные данные гусят не получавших глауконит соответственно на 7,3 и 3,9 г, 2,3 и 1,6%.
Кормовая добавка глауконит в количестве 0,3% от сухого вещества рациона гусят-бройлеров позволила сократить на единицу прироста живой массы затраты кормов, обменной энергии и протеина - на 6,7 - 6,8%, а стоимость 1 кг прироста живой массы - на 6,3%. При этом оплата корма продукцией повысилась на 7,1% в натуральном и на 6,7% в стоимостном выражении. Дополнительно полученный прирост живой массы по группе за счёт глауконита составил 202,15 кг на общую стоимость 11,32 тыс. руб.
Проведённая производственная апробация наиболее оптимальной дозировки кормовой добавки глауконита - 0,2% от сухого вещества рациона гусят показала, что она обеспечивает сохранность поголовья на уровне 97,5%, против 93,1% в группе не получавшей глауконит, увеличивает среднесуточный прирост на 9,8%, снижает затраты корма на единицу продукции на 9,0% и обеспечивает дополнительный прирост живой массы на сумму 31,87 тыс. руб.
На основании вышеизложенных результатов научного, научно-хозяйственного и научно-производственного опытов можно сделать следующие выводы и предложения производству.
ВЫВОДЫ
1. Кормление гусят-бройлеров полнорационным комбикормом позволяет к концу откорма получить прирост живой массы 3894,5 г при среднесуточном приросте 59,9 г и затратах корма на единицу продукции 3,70 кг, обменной энергии - 45,7 МДж, протеина - 682 г. При этом в потрошенной тушке выход съедобных частей составил 68,2% с содержанием в них мышечной ткани 54,7%, внутреннего жира - 2,8, кожи с подкожным жиром -23,0 и костей-19,5%.
Гусята, получавшие кормовую добавку глауконит в количестве 0,1% от сухого вещества рациона по среднесуточному приросту на 5,2% превосходили гусят, не получавших его. При этом переваримость протеина увеличилась на 2,4%, жира - на 12,0%, а отложение азота в теле было на 0,11 г выше. В съедобных частях потрошенной тушки мышечной ткани было больше на 107 г, внутреннего жира - на 29 и кожи с подкожным жиром - на 32г.
Добавка глауконита 0,2% от сухого вещества рациона гусят обеспечила увеличение среднесуточного прироста на 11,0% за счет большего переваривания протеина корма на 5,0%, жира - на 8,1% и повышенного среднесуточного отложения азота в теле на 8,8%. Это обеспечило более высокий выход в съедобных частях тушки мышечной ткани на 281 г, внутреннего жира - на!3, кожи с подкожным жиром - на 21 и костей - на 37г.
Увеличение дозировки глауконита до 0,3% от сухого вещества рациона гусят способствует повышению среднесуточного прироста живой массы на 7,0%, переваримость протеина - на 3,6 и жира - на 9,7%, а в тушках птицы увеличение мышечной ткани на 209 г, внутреннего жира — на 16, кожи в подкожным жиром — на 14г.
Самый высокий коэффициент конверсии протеина и жира рациона в продукцию обеспечивает дозировка глауконита 0,2% от сухого вещества рациона- 19,3 и 15,0%, соответственно в меньшей степени доза 0,1 и 0,3% - 17,2 и 14,7%, 18,1 и 14,4%, в то время как при откорме на одном полнорационном комбикорме ККП составил 15,8, ККЭ - 12,8%.
Глауконит в дозе 0,1% от сухого вещества рациона способствует увеличению содержания в мясе гусят жира на 1,24%, в дозе 0,2% - протеина
на 2,0%, жира - на 0,58 и золы - на 0,13%, добавка 0,3% глауконита -содержание золы на 0,30%.
Наиболее выраженные окислительно-восстановительные, а также анаболические процессы белкового и липидного обмена наблюдались в организме гусят, получавших 0,2% глауконита, в меньшей степени при 0,1 и 0,3% глауконита от сухого вещества рациона.
Откорм гусей с использованием кормовой добавки глауконит позволило уменьшить на единицу прироста затраты корма, обменной энергии и протеина на 10,1-10,3%, а оплату корма продукцией увеличить в натуральном и стоимостном выражении на 5,2 - 11,3%.
ПРЕДЛОЖЕНИЯ ПРОИЗВОДСТВУ
Для крупных промышленных гусеводческих ферм и фермерских хозяйств рекомендуем использовать в рационах гусят-бройлеров кормовую добавку глауконит в количестве 0,2% от сухого вещества рациона, что позволит повысить сохранность поголовья на 4,4%, прирост живой массы -на 9,8% и сократить затраты кормов на единицу продукции - на 9,0%.