Загальна характеристика і особливості життєдіяльності бактерій родини Enterobacteriaceae
Размещено на http://www.
Міністерство освіти і науки України
Дніпродзержинський державний технічний університет
Кафедра екології та біотехнології
Дисципліна Загальна мікробіологія
КУРСОВА РОБОТА
на тему
Загальна характеристика і особливості життєдіяльності бактерій родини Enterobacteriaceae
Дніпродзержинськ
2009
ЗМІСТ
Вступ
1. Морфологія бактерій родини Enterobacteriaceae
2. Фізіологія бактерій родини Enterobacteriaceae
3. Метаболізм бактерій родини Enterobacteriaceae
4. Генетика бактерій родини Enterobacteriaceae
5. Екологія бактерій родини Enterobacteriaceae
6. Антигени бактерій родини Enterobacteriaceae
7. Патогенність і токсиноутворення бактерій родини Enterobacteriaceae
8. Резистентність бактерій родини Enterobacteriaceae
9. Патогенез бактерій родини Enterobacteriaceae
10. Імунітет до бактерій родини Enterobacteriaceae
11. Лабораторна діагностика бактерій родини Enterobacteriaceae
12. Профілактика і лікування захворювань викликаних бактеріями родини Enterobacteriaceae
13. Динаміка зміни резистентності умовно-патогенних ентеробактерій шлунково-кишкового тракту до антимікробних препаратів
Перелік посилань
ВСТУП
Важливу роль в життєдіяльності людини відіграє мікрофлора товстого кишечника – своєрідний екстракопоральний орган. Вона є антагоністом гнилісної мікрофлори, так як продукує молочну, оцтову кислоти, антибіотики і інше. Відома її роль в водно-сольовому обміні, регуляції газового складу кишечнику, обміні білків, вуглеводів, жирних кислот, холестерину і нуклеїнових кислот, а також продуктів біологічно активних сполук – антибіотиків, вітамінів, токсинів та ін. Морфокінетична роль мікрофлори складається в її участі в розвитку органів і систем організму; вона приймає участь також в фізіологічному запаленні слизистої оболонки і зміні епітелію, перетравленні і детоксикації екзогенних субстратів і метаболітів, що порівнюється з функцією печінки. Нормальна мікрофлора виконує, крім того, антимутагену роль, руйнуючи канцерогенні речовини в кишечнику. В той же час деякі бактерії можуть продукувати потужні мутагени. Пристінкова мікрофлора кишечника колонізує слизову оболонку у вигляді мікроколоній, утворюючи своєрідну біологічну плівку, яка складається з мікробних тіл і екзополісахаридного матриксу. Екзополісахариди мікроорганізмів, які називаються глікокаліксом, захищають мікробні клітини від різних фізико-хімічних і біологічних впливів. Слизова оболонка кишечника також знаходиться під захистом біологічної плівки.
Великий вплив чинить мікрофлора кишечника на формування і підтримку імунітету. В кишечнику міститься близько 1,5 кг мікроорганізмів, антигени яких стимулюють імунну систему. Природнім неспецифічним стимулятором імуногенезу є мурамілдипептид, який утворюється із мікрофлори під дією лізоциму і інших ліричних ферментів, які знаходяться у кишечнику.
Важливою функцією нормальної мікрофлори кишечнику є її участь в колонізаційній резистентності, під якою розуміють сукупність захисних факторів організму, і конкурентних, антагоністичних і інших особливостей анаеробів кишечнику, які надають стабільність мікрофлорі і запобігають колонізації слизових оболонок сторонніми мікроорганізмами [1].
До групи кишкових бактерій відносяться дуже багато бактерії, зокрема родина Enterobacteriaceae. Це велика родина яка включає такі роди як Salmonella, Escherichia, Klebsiella, Proteus, Shigella. Більшість бактерій цієї родини складають значну частину нормальної аеробної флори кишечника і зазвичай не викликають захворювань, а навпаки, беруть участь у забезпеченні його нормального функціонування. Ці мікроорганізми стають патогенними лише у випадках проникнення їх у тканини організму, особливо в сечовидільні шляхи, легені, очеревину чи мозкові оболонки, де вони здатні викликати запальний процес. Вони можуть проникати в кров'яне русло (при використанні венозних чи уретральних катетерів); попадати в сечостатеві шляхи при проведенні інструментальних досліджень (нозокоміаль-на передача); викликати інфекційні ураження сечовидільних шляхів після введення контамінованих лікарських препаратів, особливо після внутрішньовенного їхнього застосування. Можливий статевий шлях передачі.
Бактерії родини Enterobacteriaceae часто обумовлюють вторинні інфекції, у тому числі пневмонії, ендокардити (запалення внутрішньої оболонки серця), бактеріємії, особливо в людей, що приймають наркотики, і в пацієнтів клінік.
Прояви інфекцій, викликаних бактеріями родини Enterobacteriaceae, залежать від локалізації патологічного процесу. Вони не можуть бути диференційовані з іншими бактеріальними інфекціями на підставі одних лише симптомів і ознак захворювання, тому дуже важливо визначити збудника інфекції, дати видову і типову характеристику, чутливість до антибактеріальних засобів. Системне вивчення видових, типових і інших особливостей мікроорганізмів − збудників інфекцій − має істотне значення для з'ясування патогенезу інфекцій і визначення методів їхньої терапії.
Залежно від локалізації патологічного процесу для дослідження використовують виділення статевих органів, сечу, кров, спинномозкову рідину та ін. У пофарбованих мазках виявляють грамнегативні короткі палички, що можуть утворювати ланцюжки; вони дуже схожі один на одного і лише наявність широких капсул має діагностичне значення для роду Klebsiella. Досліджуваний матеріал одночасно наносять на кров'яний агар і середовища, що містять спеціальні
барвники і вуглеводи, що дозволяє швидко диференціювати ферганські і неферганські колонії. Бактерії, виділені на цих середовищах, надалі ідентифікують за допомогою біохімічних і серологічних реакцій. Можлива швидка попередня ідентифікація при можливості мікроорганізмів ферментувати лактозу.
Виражену протимікробну дію щодо бактерій родини Enterobacteriaceae має ампіцилін, тетрацикліни, аміноглікозиди, поліміксини, сульфаніламіди. Однак чутливість до цих препаратів в окремих штамів різна, тому її необхідно визначати лабораторними методами. Нерідко зустрічається множинна стійкість до ліків.
При лікуванні ефект може бути досягнутий при одночасному застосуванні аміноглікозидів і фурагіну чи ампіциліну; триметоприму із сульфаметоксазолом і поліміксином. Доцільні й інші комбінації антибактеріальних засобів, призначення препаратів, що потенціюють дію антибіотиків, що підвищують захисні механізми організму хворих, що сприяють доставці антибактеріальних засобів до вогнищ ураження й забезпечують їхню терапевтичну концентрацію в тканинах і сироватці крові.
Профілактика й ефективність боротьби з цими інфекціями багато в чому залежать від дотримання чистоти рук, суворого дотримання правил асептики й антисептики (стерилізації інструментарію, дезінфекції устаткування), обережності в призначенні внутрішньовенних препаратів і особистої гігієни сечостатевих шляхів. Дуже важливо пам'ятати, що багато грамнегативних бактерій є патогенними і викликають розвиток захворювання в ослабленому організмі. У лікувальних установах ці бактерії нерідко переносяться персоналом, а також через інструменти чи препарати для парентерального введення [2].
1. МОРФОЛОГІЯ БАКТЕРІЙ РОДИНИ ENTEROBACTERIACEAE
Мікроорганізми родини Enterobacteriaceae являють собою палички довжиною 1-5 мкм і товщиною 0,4-0,8 мкм. Вони добре фарбуються аніліновими барвниками, грамнегативні. Багато з них рухливі завдяки наявності перетрихіально розташованих джгутиків і можуть мати капсулу. Спор не утворюють. У багатьох штамів, які належать до різних родів ентеробактерій: ешерихіям, сальмонелам, шигелам, клебсієлам, протеям, виявлені ворсинки (пілі) загального типу, а у деяких штамів – статеві ворсинки (F-пілі), з якими зв’язані їх адгезивні та інші властивості [3,4].
Ешерихії
Рід E. coli представлений єдиним видом і багато чисельними біоварами і сероварами, які розрізняються деякими біохімічними, антигенними і патогенними властивостями.
Кишкову паличку (рис. 1.1) вперше виділив із фекалій людини в 1885 році німецький педіатр і мікробіолог Теодор Ешерих. В подальшому було встановлено, що подібні бактерії широко розповсюдженні в природі. Вони зустрічаються в кишечнику майже всіх видів ссавців, птахів, риб, а також в воді, ґрунті і інших об’єктах навколишнього середовища.
За своїми морфологічними і тинкторіальними властивостями E. coli нагадує інші ентеробактерії. Це дрібні грамнегативні палички довжиною 2-3 мкм, товщиною 0,5-0,7 мкм з закругленими кінцями, в мазках розташовуються не упорядковано. Серед кишкових паличок зустрічаються рухливі і не рухливі варіанти. Деякі штами мають виражену капсулу і утворюють слизові колонії. S-форми E. coli на поживному агарі формують опалово-мутнуваті, слабко випуклі, вологі, з рівними краями і блискучою поверхнею колонії, які легко суспендуються в сольовому розчині. Перехідні S- і R-форми утворюють пласкі, з шершавою поверхнею і нерівними краями колонії, які погано суспендуються в сольовому розчині. Кишкова паличка викликає рівномірне помутніння рідини поживного середовища з утворенням невеликого осаду [3].
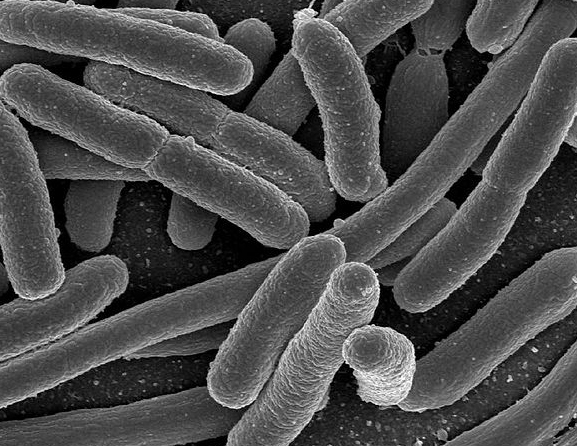
Рисунок 1.1 - Escherichia coli (за [5])
Сальмонели
До роду Salmonella (рис. 1.2) відносяться збудники черевного тифу, паратифу і сальмонельозів. Назва роду походить від імені американського дослідника Даніеля Сальмона. Сальмонели підрозділені на 4 підроду, які включають 65 серогруп і близько 2000 сероварів.
Сальмонели відповідають в основному тим ознакам, які присутні всім ентеробактеріям. При мікроскопії мазків, забарвлених за грамом, черевнотифозні і паратифозні бактерії мають вигляд паличок червоного кольору із заокругленими кінцями розміром 0,5-0,8×1-3 мкм, активно рухливі у висячій чи роздавленій краплі.
Ріст у МПБ супроводжується помутнінням. На МПА виростають ніжні, круглі, гладенькі, прозорі або напівпрозорі колонії розміром 2-4 мкм. Однак колонії тифозних мікробів, що мають Vi-антиген, каламутні. У S. Schottmuelleri колонії грубіші, через кілька днів по периферії колонії утворюється слизовий валик.
На середовищах Ендо, Левіна, Плоскирєва колонії безбарвні, прозорі, часом рожевуваті (Ендо) або злегка голубуваті (Левіна). На вісмут-сульфатному агарі черевнотифозні мікроби утворюють колонії чорного кольору, іноді зі світлим обідком. Паратифозні бактерії на цьому середовищі можуть утворювати коричневі або зеленуваті колонії. Після зняття колонії залишається чорний слід [3,6].
Рисунок 1.2 - Salmonella (за [7])
Шигели
Бактерії роду Shigella (рис. 1.3) є збудниками бактеріальної дизентерії або шигельозу.
Перші припущення про роль бактерій в етіології дизентерії були зроблені А. Шентемесом і Ф. Відалем в 1888 році.
Дизентерія – поліетіологічне захворювання. Його викликають різні види бактерій, які називаються шигелами. В теперішній час вони віднесені до роду Shigella, який підрозділяється на чотири групи: A,B,C і D – і більше двох десятків сероварів.
За своїми морфологічними властивостями шигел мало відрізняються від ешеріхій і сальмонел. Однак вони позбавленні джгутиків і тому є нерухомими бактеріями. Багато штамів шигел мають ворсинки загального типу і статеві ворсинки. Різні види шигел ідентичні за своїми морфологічними властивостями [3].
Рисунок 1.3 - Shigella (за [8])
Клебсієли
У мазках забарвлених за Грамом, капсульні форми клебсієл мають вигляд грамнегативних, еліпсоподібних, товстих паличок довжиною 5-8 мкм і шириною 3-5 мкм. Без капсульні форми мають менші розміри (0,3-0,6×1-3 мкм), поодиноке, парне або ланцюжкове розташування [6].
Протей
Усі види роду Proteus (рис. 1.4) являють собою грамнегативні прямі палички розмірами 0,4-0,6×1,0-3,0 мкм. Зустрічаються кокоподібні, нитковидні форми. Спор і капсул не утворюють, мають перетрихіально розташовані джгутики [3].
Рисунок 1.4 - P. vulgaris (за [9])
2. ФІЗІОЛОГІЯ БАКТЕРІЙ РОДИНИ ENTEROBACTERIACEAE
Усі ентеробактерії є факультативними анаеробами, добре ростуть на середовищах з м’ясним екстрактом. Вони володіють вираженою ферментативною активністю, яка пов’язана з утворенням багато чисельних сахаридолітичних, протеолітичних і інших ферментів, як конституційних, так і індуцибельних. Разом з тим відмінності які спостерігаються в перерахованих ознаках мають таксономічне значення і використовуються для родинної і видової диференціації і ідентифікації ентеробактерій. Деякі з цих ознак слугують підставою для визначення окремих біоварів серед одного і того ж виду бактерій.
До основних ознак, які вказують на належність ентеробактерій до одного з п’яти триб відносяться:
1) характер продукту, який утворюється при зброджуванні глюкози: кислоти, бутандіол, їх суміші;
2) реакція з метиловим червоним;
3) утворення ацетилметилкарбінолу і позитивна реакція Фогеса-Проскауера;
4) відновлення нітратів;
5) утворення уреази;
6) ріст в присутності KCN і деякі інші.
Головною біохімічною ознакою, яка слугує для визначення родової і видової належності ентеробактерій, є їх здатність ферментувати різні вуглеводи до утворення кислоти або газу, утворення індолу, сірководню, декарбоксилаз амінокислот (лізину, орнітину та ін.), утилізація цитрату, тартрату та інші [3].
Ешерихії
Це факультативний анаероб, добре росте на звичайних поживних середовищах при слабо лужній реакції середовища і оптимальній температурі 37°С. Ріст і розмноження бактерій можливі при доволі значних коливаннях рН середовища і температурного режиму.
Кишкова паличка має
найбільш високу ферментативну активність.
Вона утилізує ацетат в якості єдиного
джерела вуглецю, відновлює нітрати до
нітритів. Ферментує глюкозу та інші
вуглеводи з утворенням пірувату, який
потім перетворюється в молочну, оцтову,
мурашину кислоти виділяються CO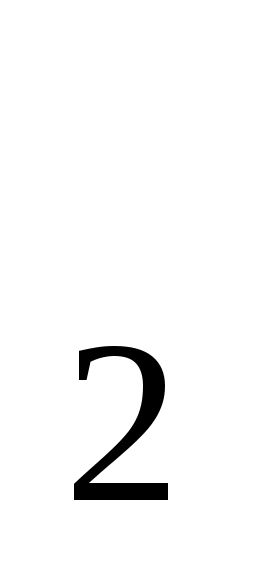 і H.
Більшість штамів ферментує лактозу.
Однак зустрічаються штами, які повільно
зброджують лактозу або зовсім не
володіють цією здатністю [3].
і H.
Більшість штамів ферментує лактозу.
Однак зустрічаються штами, які повільно
зброджують лактозу або зовсім не
володіють цією здатністю [3].
Сальмонели
Вони є ферментативно активними бактеріями, хоча деякі поступаються в цьому відношенні ешеріхіям. Диференціація багато чисельних представників даного роду проводиться на основі їх ферментативної активності і антигенної структури наведено в таблиці 2.1 [3].
Таблиця 2.1 - Ферментативні властивості ешерихій і тифозно-паратифозних бактерій (за [6])
|
Вид |
Лактоза |
Глюкоза |
Мальтоза |
Маніт |
Сахароза |
Індол |
H |
|
Escherichia coli Salmonella typhi Salmonella parathyphi A Salmonella schottmuelleri |
кг - - - |
кг к кг кг |
кг к кг кг |
кг к кг кг |
кг± - - - |
+ - - - |
± + - + |
Примітка: к-утворення кислоти; кг-утворення кислоти і газу; + - виявлення ознаки; − відсутність ознаки; ± - можливе виявлення ознак.
Шигели
Збудники дизентерії хемоорганогетеротрофні, невимогливі к поживним середовищам. На щільних середовищах при виділенні із організму хворого утворюються, як правило, S-форми колоній. Шигели виду sonnei утворюють два типи колоній − S- (I фаза) і R- (II фаза). Бактерії I фази при пересівах утворюють обидва типи колоній. Оптимальна температура росту 37°С, рН середовища 7,2.
Шигели менш ферментативно активні, ніж інші ентеробактерії: при зброджуванні глюкози та інших вуглеводів утворюються кислі продукти без газоутворення.
Шигели не розщеплюють лактозу і сахарозу, за виключенням S. sonnei, які повільно (на 2-у добу) розщеплюють ці сахариди, наведено в таблиці 2.2 [3].
Таблиця 2.2 - Біохімічні властивості шигел (за [6])
|
Вид |
Ферментація вуглеводів |
Індол |
Каталаза |
|||||
|
лактоза |
глюкоза |
мальтоза |
маніт |
дульцит |
сахароза |
|||
|
S. dysenteriae S. flexneri S. boydii S. sonnei |
- - - + повільно |
+ + + + |
- + ± + |
- + + + |
- + + - |
- - - + повільно |
- - + - |
- - - + |
Клебсієли
Вони невимогливі до поживного субстрату. Добре ростуть на основних поживних середовищах, утворюючи куполоподібні блискучі слизові колонії. Викликають інтенсивне помутніння середовища при рості у бульйоні.
Більшість штамів ферментують глюкозу з утворенням кислоти і газу; використовують глюкозу і цитрат в якості єдиного джерела вуглецю, а аміак - як джерело азоту.
Диференціація різних видів клебсієл проводиться на основі їх різних здатностей ферментувати вуглеводи, утворювати уреазу і лізиндекарбоксилазу, утилізувати цитрат та інших властивостей наведено в таблиці 2.3.
Клебсієли є факультативними анаеробами. Оптимальна температура для їх росту 35-37°С [3].
Таблиця 2.3 - Диференціальні ознаки клебсієл (за [6])
|
Вид |
Ферментація |
Індол |
Реакція Фогеса-Проскауера |
Утилізація цитрату |
|||
|
Глюкоза |
Лактоза |
Сахароза |
Сечовина |
||||
|
K. pneumoniae K. oxytoca K. planticola K. terrigena K. ozaenae K.rhinoskleromatis |
+ + + + ± ± |
+ + + + ± − |
+ + + + + ± |
+ + + - - - |
- + ± - - - |
+ + + + - - |
+ + + ± ± - |
Протеї
Вони не вимогливі до поживного субстрату, добре ростуть на основних поживних середовища. На середовищах Ендо і Плоскирєва виростають прозорі блискучі безбарвні колонії. На вісмут-сульфатному агарі через 48 годин утворюються сірувато-коричневі колонії, під якими формується чорно-коричнева зона. Н-форма (джгутикова) на простому агарі дає характерний повзучий ріст або „феномен роїння”. Культура має неприємний гнильний запах. При посіві у стовпчик напіврідкого агару визначають рухливість. Повзучий характер росту протею використовують для виділення чистих культур посівом у конденсаційну воду скошеного агару за методом Шукевича. Протеї є факультативними анаеробами. Ростуть при температурі 20-37°С.
Ферментують багато вуглеводів з утворенням кислих продуктів, глюкозу розщеплюють з утворенням кислот і невеликої кількості газу. Різні види відрізняються один від одного за ферментацією вуглеводів, утворенню індолу, уреази, сірководню, орнітиндекарбоксилази та іншим ознакам, наведено в таблиці 2.4 [3,6].
Таблиця 2.4 - Диференціальні ознаки бактерій роду Proteus (за [6])
|
Вид |
Ферментація |
Утворення |
Орнітин-декарбокси- лаза |
Реакція Фогеса-Проскауера |
|||
|
мальтози |
сахарози |
ксилози |
індолу |
H |
|||
|
P. vulgaris P. mirabilis P. myxofaciens P. penneri |
+ - + + |
+ - + + |
+ + - + |
+ - - - |
+ + - ± |
- + - - |
- ± + - |
3. МЕТАБОЛІЗМ БАКТЕРІЙ РОДИНИ ENTEROBACTERIACEAE
Більшість ентеробактерій можуть використовувати в якості субстратів для дихального метаболізму велику кількість простих органічних сполук – органічні кислоти, амінокислоти, вуглеводи. В аеробних умовах, всі ці бактерії добре ростуть на звичайних складних бактеріологічних середовищах, азотисті компоненти яких (амінокислоти і пептиди) забезпечують їх окислюючими субстратами. В анаеробних умовах, однак, ріст стає строго залежним від наявності зброджує мого вуглеводу. Деякі моносахариди, дисахариди і поліспирти зброджуються всіма представниками даної групи. Використання полісахаридів менш розповсюджено.
Хоча ентеробактерії вирощують на складних середовищах, мінімальні харчові потреби цих організмів, зазвичай прості. Представники багатьох родів взагалі не потребують факторів росту (наприклад Escherichia coli, багато видів Salmonella), а у ауксотрофних організмів ці потреби зазвичай прості. Особливо розповсюджена потреба в нікотиновій кислоті, що характерна для багатьох видів роду Proteus. У Salmonella typhi є специфічна потреба в триптофані.
Дослідження механізму регуляції біосинтезу амінокислот у багатьох ентеробактерій виявило характерні особливості цього процесу, які відрізняють данні організми від всіх інших бактерій. Наприклад, перша стадія в біосинтезі амінокислот родини аспартату, перетворення аспарагінової кислоти в аспартилфосфат, у ентеробактерій завжди здійснюється трьома ізофункціональними аспартаткіназами: аспартаткіназою I, яка і інгібується, і репресується треоніном, аспартаткіназою II, яка репресується, але не інгібується метіоніном, і аспартаткіназою III, яка і інгібується, і репресується лізином. Цей тип регуляції аспарагінового шляху не був виявлений ні в однієї бактерії не кишкової групи.
ГЦ-склад ДНК ентеробактерій варіює в широких межах – від 37 до 63%. За виключенням роду Proteus, представники якого суттєво відрізняються за нуклеотидним складом. ГЦ-склад ДНК споріднених організмів трьох видів - Salmonella, Escherichia і Shigella – розрізняються зовсім мало. Діапазон змінення
ГЦ-складу для „класичних” бактерій родини Enterobacteriaceae (від 37 до 59%) дуже близький до діапазону для представників родини з полярним джгутикуванням (від 39 до 63%) [10].
Бродіння
Зброджування сахаридів у ентеробактерій відбувається за шляхом Ембдена-Мейєргофа. Продукти бродіння у різних представників групи розрізняються і якісно, і кількісно. Однак у цих процесів є одна характерна біохімічна властивість, яка рідко зустрічається при інших типах бактеріального бродіння. Мова йде про особливий спосіб розщеплення однієї з проміжних сполук – піровиноградної кислоти (ПВК), що веде до утворення мурашиної кислоти:
СН>3>СОСООН + КоАSН→СН>3>СОSКоА+ НСООН.
Таким чином, мурашина кислота часто є основним кінцевим продуктом бродіння. Вона накопичується, хоча не завжди, оскільки деяки з цих бактерій володіють формнатгідрогенліазою, яка розщеплює мурашину кислоту до СО>2> і Н>2>:
НСООН → СО>2>+Н>2>.
У таких організмів замість мурашиної кислоти як кінцевого продукту бродіння утворюються еквімолярні кількості Н>2> і СО>2>.
Найбільш розповсюдженим типом перетворення сахаридів при бродінні у ентеробактерій є так зване бродіння змішаного типу, яке приводить до утворення молочної, оцтової, бурштинової і мурашиної кислот (або СО>2> і Н>2>), а також етилового спирту. Такий тип бродіння властив представникам родів Salmonella, Escherichia, Shigella і Proteus. Відношення кількості різних кінцевих продуктів різниться як від штаму до штаму, так і для одного штаму при рості в різних умовах, наприклад при різних значеннях рН. Така різниця відображає ту обставину, що кінцеві продукти утворюються з піровиноградної кислоти трьома незалежними шляхами (рис. 3.1). Утворення газу в результаті зброджування сахаридів є дуже важливою ознакою для ентеробактерій – газоутворювальні бактерії роду Escherichia відрізняються за цим критерієм від патогенних видів групи Shigella і Salmonella typhi, які зброджують сахариди без виділення газу. В випадку простого бродіння змішаного типу газ може утворюватися тільки за рахунок розщеплення мурашиної кислоти; відповідно, утворення газу відображає наявність форміатгідрогенліази. Цей ферментний комплекс, звісно, не суттєвий для бродіння і може загубитися в результаті мутації без зміни здатності бактерії до бродіння. І дійсно, дослід показує, що в природі існують „неаерогенні”, тобто не утворюючі газоподібних продуктів штами такого типового продуценту їх, як Escherichia coli. Тому, хоча виділення газу являє собою корисну ознаку при класифікації ентеробактерій, даний критерій не є безпомилковим.
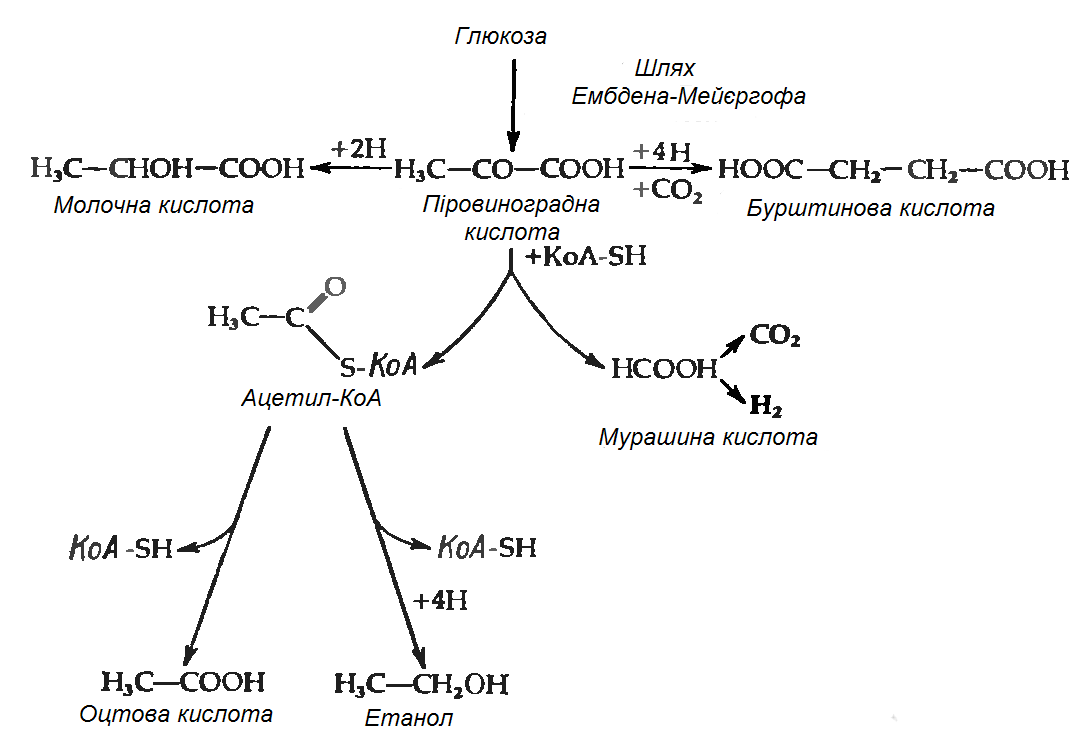
Рисунок 3.1 - Шляхи утворення із ПВК типових кінцевих продуктів кислотного бродіння змішаного типу (за [10])
Іншою ознакою, яка має велике значення для ідентифікації ентеробактерій, є здатність зброджувати дисахарид лактозу, що обумовлено наявністю у цих організмів β-галактозідази. Ефективність зброджування лактози залежить ще і від наявності специфічної пермеази галактозидів, яка сприяє проникненю лактози в клітину. Штами які мають β-галактозідазу, але позбавленні пермеази, не можуть поглинати лактозу зі швидкістю, яка достатня для інтенсивного бродіння, і зазвичай класифікуються як не здатні до зародження цього сахариду. Зброджування лактози властиво Escherichia, але відсутні у Salmonella, Shigella і Proteus. Деякі штами Shigella утворюють β-галактозідазу, але не можуть зброджувати лактозу із-за відсутності пермеази [10].
4. ГЕНЕТИКА БАКТЕРІЙ РОДИНИ ENTEROBACTERIACEAE
Відкриття явища переносу генів у ентеробактерій при кон’югації і трансдукції дозволило доволі детально дослідити генетичні родинні відносини між деякими її представниками. Утворення хромосомних гібридів між E. coli і бактеріями родів Salmonella і Shigella каже про дуже високу ступінь генетичної гомології цих бактерій, що підтверджується і порівнянням хромосомних карт двох найбільш вивчених в цьому відношенні видів − Salmonella typhimurium і Escherichia coli. Як видно на рис. 4.1 і 4.2, де представленні в спрощеному вигляді хромосомні карти цих двох організмів, багато маркерів розташовуються на обох хромосомах в однакових локусах.
Про близьку генетичну спорідненість груп Escherichia, Salmonella і Shigella свідчить також висока ступень ДНК-ДНК-гібридизації in vitro. Утворення ж хромосомних гібридів між бактеріями цієї підгрупи і представниками інших родів ентеробактерій (Proteus, Enterobacter) відбувається дуже рідко. Крім того, випробування по гібридизації ДНК-ДНК вказують, що ступень генетичної гомології між представниками підгрупи Escherichia− Salmonella− Shigella та іншими групами ентеробактерій доволі низька. Ентеробактерії, що належать до різних родів, можуть отримувати плазміни від донорних штамів E. coli при кон’югації і потім зберігати їх як позахромосомні елементи (ефективність кон’югації варіює в широких межах; таблиця 4.1). Таким чином серед бактерій кишкової групи можуть розповсюджуватися F-фактори, які містять додаткові гени (наприклад F-lac), і R-фактори, які визначають стійкість до різноманітних лікарських засобів [10].
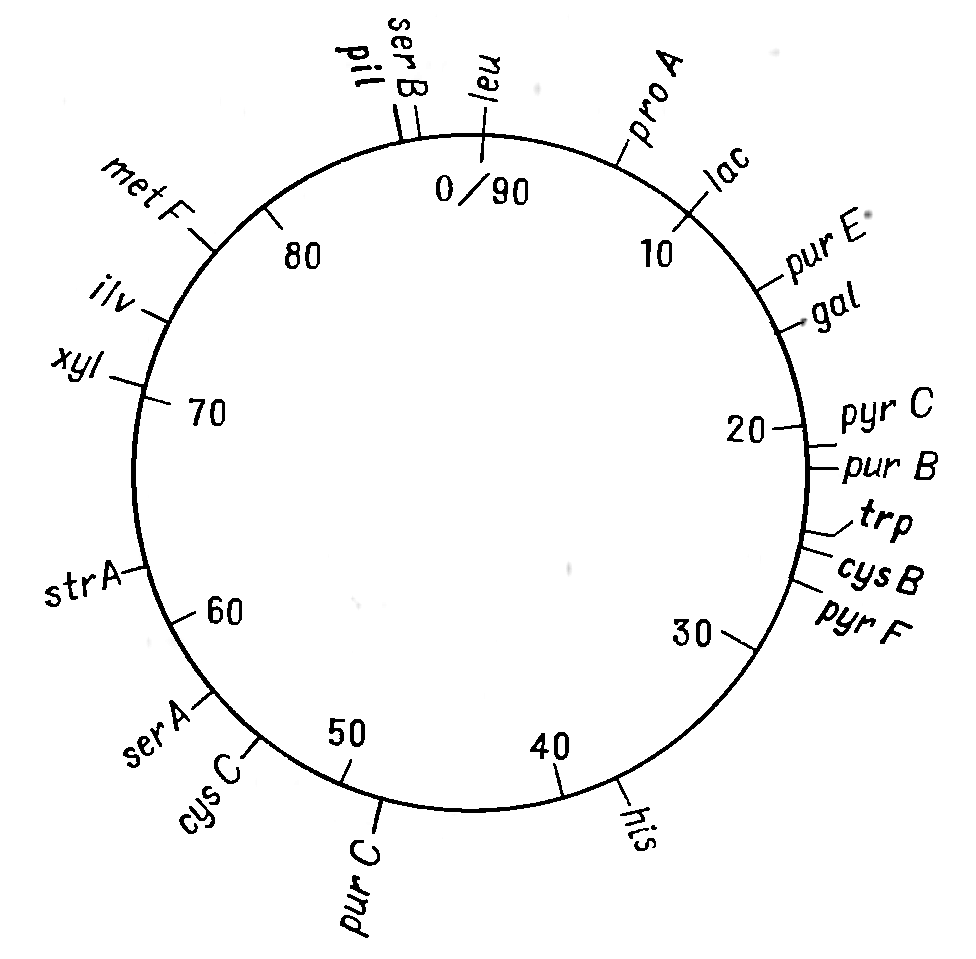
Рисунок 4.1 - Генетична карта E. coli К12 в спрощеному вигляді (за [10])
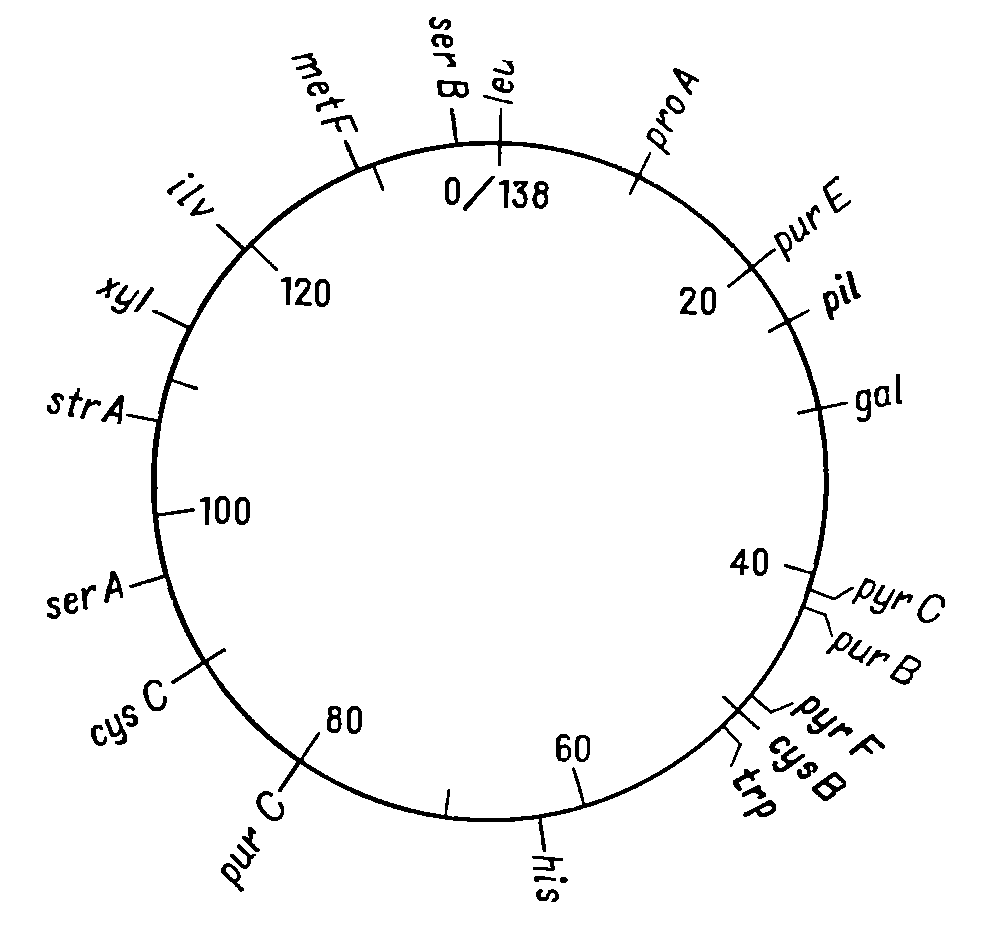
Рисунок 4.2 - Генетична карта S. typhimurium в спрощеному вигляді (за [10])
5. ЕКОЛОГІЯ БАКТЕРІЙ РОДИНИ ENTEROBACTERIACEAE
Середовищем проживання для більшості ентеробактерій є кишковий тракт хребетних тварин і людини.
В організмі людини багато ентеробактерій містяться в складі мікробних біоценозів тонкого і товстого кишечника. Патогенні види зустрічаються тільки у хворих і бактеріоносіїв.
З випорожненнями людей і тварин ентеробактерії потрапляють в навколишнє середовище. В ній вони можуть зберігатися на протязі різних строків в залежності від виду і умов. Деякі з них (E. coli) використовуються в санітарній мікробіології в якості показників фекального забруднення навколишнього середовища [3].
Кишкова паличка наприклад може зберігатися в воді і ґрунті декілька місяців [11].
Широке розповсюдження сальмонел в природі і різноманіття шляхів, якими вони проникають в організм людини, пояснюється їх генетичною пластичністю. Сальмонели легко знаходять екологічні ніші і адаптуються до найрізноманітніших умов [12].
Клебсієли – широко розповсюджені бактерії. Вони порівняно стійкі до факторів навколишнього середовища завдяки наявності капсули і можуть на протязі тривалого часу зберігатися в ґрунті, воді, приміщеннях. Разом з тим вони зустрічаються в складі мікробних біоценозів в організмі людини і тварин [3].
Протеї P. vulgaris і P. mirabilis є мешканцями кишечника багатьох тварин, виявляються також в стічних водах і ґрунті. Всі види протеїв можуть виділятися з хворих людей [3].
6. АНТИГЕНИ БАКТЕРІЙ РОДИНИ ENTEROBACTERIACEAE
Антигенна будова служить одним із суттєвих критеріїв, на яких основана класифікація, а також ідентифікація ентеробактерії. Розрізняють три основних типи антигенів: 1) О-соматичний антиген; 2) Н-джгутиковий антиген; 3)К-антигени. О-антиген є складовою частиною ліпополісахариду (ЛПС) зовнішнього шару клітинної стінки. Специфічність О-антигену визначається детермінантними сахаридами (гексозами і аміносахаридами), ковалентно зв’язаними з базисною частиною ЛПС. Н-антиген локалізується в джгутиках клітини. Він складається з білка флагеліну. Капсульні антигени К-антигени, так як і О-антигени, містяться в ЛПС клітинної стінки, але в більш поверхневому шарі. Вони маскують О-антигени.
В зв’язку з цим для визначення О-антигену у бактерій, які володіють К-антигеном, необхідно зруйнувати останній шляхом кип’ятіння або автоклавування культури. К-антигени за хімічними властивостями відносяться до кислих полісахаридів. До К-антигенів відносяться К-антигени E. coli, М-антигени Salmonella schottmuelleri, Vi-антиген збудника черевного тифу, який виявляється також у S. parathyphi C та деяких штамів E. coli. Всі ці антигени характеризуються імуннохімічною специфічністю, що дозволяє диференціювати роди і види, а також виділяти серед них серогрупи і серологічні варіанти (серовари). Антигенну будову ентеробактерій вивчають в реакціях аглютинації, непрямої (пасивної) гемаглютинації, преципітації, зокрема при імунно-електрофорезі та інших імунологічних реакціях з відповідними діагностичними сироватками.
Крім антигенів які перелічені, у ентеробактерії виявленні загальні антигени. Один з них (антиген Куніна) зв’язаний з ЛПС, інший – з білками клітинної стінки. Антиген Куніна подібне до антигену, який виявлений в товстому кишечнику людей, які страждають виразковим колітом [3].
Ешерихії Кишкова паличка має складну антигену структуру. Вона містить соматичний, або О-антиген, поверхневий (капсульний, оболонковий) К-антиген і джгутиковий Н-антиген. О-антигени у бактерій триби має схожу хімічну будову і зв’язані з ЛПС клітинної стінки. Н-антигени є тільки у джгутикових форм бактерій і складаються з білка флагеліну.
О-антигени є основними антигенами які визначають серологічну групу ешеріхій. В теперішній час описано близько 170 О-серогруп E. coli. Більшість ешеріхій, що належать до різних О-серогруп, зв’язані одна з одною перехресними антигенними зв’язками. Близько 100 серогруп мають антигенні зв’язки з шигелами, сальмонелами та іншими ентеробактеріям.
К-антигени представленні трьома антигенами, які позначаються літерами A,B і L. Вони відрізняються один від одного чутливістю до нагрівання і хімічним речовинам. К-антигени мають здатність маскувати О- антигени, які можна виявити тільки після руйнування перших кип’ятінням культури. Ешерихії містять близько 97 різних К-антигенів, переважно В-типу.
Н-антигени на відміну від О- і К-антигенів є типоспецифічними. У ешеріхій описано близько 50 різних Н-антигенів.
Антигени ешеріхій позначаються антигенними формулами, які вказують на серогруп, наприклад E. coli О126:К6О(В6), або серовар − E. coli О126:К6О(В6):Н2 [3,13].
Сальмонели
Сальмонели, так як і ешерихії, мають складну антигенну структуру. Вони містять О-антигени (соматичні) та Н-антигени (джгутикові). Деякі сальмонели мають К-антиген. За своєю хімічною структурою О-антигени сальмонел аналогічні тим же антигенам ешерихій. Вони відрізняються один від одного тільки структурою кінцевих ланок полісахаридного компоненту ЛПС, який визначає імуннохімічну специфічність грамнегативних бактерій. Н-антигени можуть існувати в двох різних фазах: специфічній 1-й фазі і менш специфічній, або груповій, 2-й фазі. Систематична розробка антигенної будови різних сальмонел була проведена Ф. Кауфманом, який в 1934 році запропонував серологічну класифікацію цих бактерій. Всі відомі на той час сальмонели (близько 700) він розділив на 44 серологічних типи в реакціях аглютинації з діагностичними моно рецепторними сироватками.
Ця схема в подальшому неодноразово розширювалася Ф. Кауфманом та П. Уайтом і зараз включає близько 2000 сероварів сальмонел, які входять в склад 4 підродів.
До підроду I − S. kauffmani − відноситься більша частина патогенних для людини сальмонел серологічних груп A, B, C, D, E.
Аналіз антигенної будови є обов’язковим елементом мікробіологічної діагностики сальмонельозів. В основу схеми Ф. Кауфмана і П. Уайта закладена общин О-антигенів сальмонел, які об’єднанні в серологічні групи, які позначаються літерами латинського алфавіту. Диференціація сальмонел всередині групи проводиться на основі особливостей їх Н-антигенів. Фаза 1 позначається прописними літерами латинського алфавіту: a, b, c тощо, фаза 2 − арабськими числами і, рідше, латинськими літерами. Кожному сировару привласнюють одна з видових назв з відповідною антигенною формулою, наприклад: S. enteritidis 1, 9, 12 [3].
Сальмонели − збудники черевного тифу і паратифів А і В (S. typhi, S. parathyphi A, S. schottmuelleri)
Сальмонели черевного тифу і паратифу разом з О- і Н-антигенами можуть містити і Vi-антиген. Він відноситься до К-антигенів і являє собою полімер N-ацетилгалактозаміноуронової кислоти. Наявність у сальмонел Vi-антигену, так як і ешерихій К-антигену, перешкоджає аглютинації бактерій О-сироватками. Vi-антиген міститься в вірулентних штамах сальмонел і є специфічним рецептором для деяких фагів, які називаються Vi-фагами. Це дало змогу розробити методику визначення фаговарів черевнотифозних і паратифозних сальмонел і розподілити їх по певним групам. Визначення фаговару згаданих сальмонел, так як і серовару, проводиться для епідеміологічного аналізу черевного тифу і паратифів з метою встановлення джерела інфекції, шляхів і засобів її розповсюдження [3].
Шигели так як і ешерихії і сальмонели мають складну антигенну структуру. В складі їх клітинних стінок є О-, а у деяких видів (шигели Флекснера) і К-антигени. За хімічною структурою вони аналогічні антигенам ешеріхій. Відмінності складаються головним чином в структурі кінцевих ланок полісахаридного компоненту ЛПС, які обумовлюють імуннохімічну специфічність, що дає змогу диференціювати їх від інших ентеробактерій і між собою. Крім того, шигели мають перехресні антигенні зв’язки з багатьма серогрупами ентеропатогенних ешерихій, які викликають головним чином дизентерієподібні захворювання, і з іншими бактеріями [3].
Клебсієли
Клебсієли містять О- і К-антигени. Всього відомо 11 О-антигенів і 80 К-антигенів. Останні зв’язані з капсулами. Серологічна класифікація клебсієл основана на їх антигенних відмінностях. У штамів однієї і тієї ж О-серогрупи можуть бути різні К-антигени і навпаки. Деякі О- і К-антигени клебсієл споріднені О-антигенам ешерихій і сальмонел. Найбільша кількість О- і К-антигенів знайдено у K. pneumoniae. За своїм хімічним складом ці антигени принципово не відрізняються від подібних антигенів інших ентеробактерій [3].
Протеї
Протеї володіють принаймні двома антигенами. Один з них представлений джгутиковим Н-антигеном, а другий – О-антиген − є ЛПС клітинної стінки. У трьох видів – P. morganii, P. rettgeri, P. inconstans ідентифіковано декілька десятків сероварів.
Деякі серовари протея
(ОХ-штами) мають антигени, які перехресно
реагують з антигенами рикетсій. Раніше
ця властивість використовувалась при
серодіагностиці сипного тифу – реакція
Вейля-Фелікса. В якості антигенів для
цієї реакції слугували культури Proteus
OX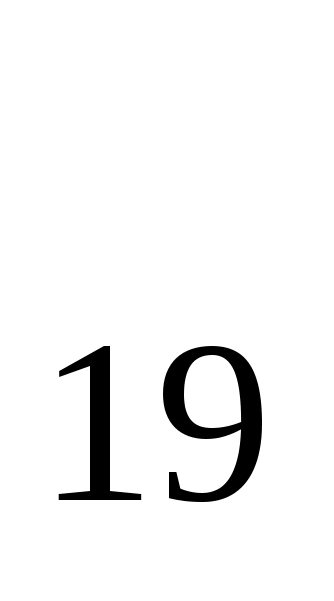 [3].
[3].
7. ПАТОГЕНІСТЬ І ТОКСИНОУТВОРЕННЯ БАКТЕРІЙ РОДИНИ ENTEROBACTERIACEAE
Патогенна дія ентеробактерій пов’язана з ліпополісахаридами клітинної стінки.
Вірулентність ентеробактерій визначається їх адгезивною здатністю, яка обумовлена позитивним хемотаксисом між поверхневими структурами мікробу і рецепторами епітеліальних клітин. Крім того, адгезивність пояснюється наявністю у багатьох ентеробактерій ворсинок, а також структурою ЛПС. Після „прилипання” до епітеліальних клітин ешерихії, які викликають дизентерієподібні колі-інфекції, і шигели проникають всередину цих клітин і там розмножуються: сальмонели розмножуються в макрофагах лімфоїдної тканини тонкого кишечника (в пейерових бляшках) і внутрішніх органах, а ешерихії – на поверхні клітин. Потім бактерії поступають у просвіт кишечника.
Токсигенність ентеробактерій обумовлена ендотоксином і екзотоксинами (ентеротоксинами і цитотоксинами). Перший являє собою ЛПС клітинної стінки. Він вивільняється тільки після руйнування бактеріальних клітин. Ентеротоксигенні ешерихії продукують два типи ентеротоксину білкової природи, які розрізняються чутливістю до температури. Їх утворення контролюється Ent-плазмідою. Термолабільний ентеротоксин, який утворюється ешерихіями і шигелами, за своїми властивостями схожий на холероген, який продукується холерним вібріоном. Багато ешерихій, які викликають колі-ентерити і дизентерієподібні захворювання, а також шигели Флекснера, Зонне, сальмонели тощо, здатні утворювати ентеротоксини [3].
До дійсного часу систематизовано біля ста патогенних серотипів кишкової палички, що викликають захворювання в людини, тварин, у тому числі і птахів.
З представників групи кишкової палички найбільш патогенної вважають підгрупу A. aerogenes (І.С.Загаєвський). Ці бактерії часто викликають колібактеріоз у телят і дітей, важкі мастити в корів, гостре запалення легких і сечостатевих шляхів у людини і тварин. Крім захворювання, деякі види бактерій кишкової палички викликають псування молока і молочних продуктів [14].
Сальмонели утворюють ендотоксин, який виявляє ентотропну, нейротропну і пірогенну дію. Білки зовнішньої мембрани обумовлюють адгезивні властивості. Стійкість до фагоцитозу пов’язана з наявністю мікрокапсули.
Вірулентні властивості шигел визначаються їх адгезією на епітельних клітинах товстого кишечника, проникненням і розмноженням в цих клітинах. Токсигенність шигел пов’язана з утворенням двох білкових токсинів: ентеротоксину і цитотоксину. За механізмом дії ентеротоксин нагадує холероген, хоча його токсичність виявляється набагато слабше. Цитотоксин шигел порушує синтез білка на рибосомах клітин кишечника. Разом з тим після руйнування бактеріальних клітин вивільняється ендотоксин, який діє на нервову і судинну системи.
Бактерії роду Proteus відносяться до умовно-патогенних мікроорганізмів. Їх патогенність визначається ЛПС клітинної стінки, з якими пов’язані вірулентні і токсичні властивості (ендотоксин) [3].
8. РЕЗИСТЕНТНІСТЬ БАКТЕРІЙ РОДИНИ ENTEROBACTERIACEAE
Ешерихії володіють більшою стійкістю до факторів навколишнього середовища, ніж шигели і сальмонели. Вони можуть на протязі декількох місяців зберігатися в воді і ґрунті, гинуть при нагріванні до 55°С на протязі години, а при 60°С − через 15 хв.; в 5% розчині фенолу, 3 % розчині хлораміну і в розчині сулеми 1:1000 – через декілька хвилин [3].
Сальмонели доволі стійкі до низької температури – в холодній воді можуть зберігатися до 1,5 років; дуже чутливі до дезінфікуючих засобів, високої температури, ультрафіолетовому випромінюванню. В харчових продуктах (м’ясі, молоці та ін.) сальмонели можуть не тільки довго зберігатися, але й розмножуватися [1].
Різні види шигел неоднаково стійкі до факторів навколишнього середовища. Найбільшою стійкістю володіють шигели Зонне, які виживають у воді відкритих водоймищ до 1½ міс., а в водопровідній воді при кімнатній температурі – до 2½ міс., в той час як шигели Флекснера – до 2 тижнів. Шигели групи А найменш стійкі до різних фізичних і хімічних дій [1].
Клебсієли гинуть при температурі 65°С через 60 хв., в розчинах звичайних дезінфікуючих засобів.
До багатьох антибіотиків (пеніциліни, макроліти тощо) клебсієли стійкі. Вони володіють чутливістю до ампіциліну, аміноглікозидам, тетрацикліну, левоміцетину. В останній час збільшилась кількість резистентних форм до перелічених антибіотиків.
Протеї порівняно стійкі до дії факторів навколишнього середовища. Переносять нагрівання при температурі 60°С на протязі години, а також зберігають життєздатність в слабких розчинах фенолу та інших дезінфікуючих речовин. Протеї володіють стійкістю до багатьох антибіотиків.
Найбільш чутливі до аміноглікозидів, особливо до гентаміцину. Відмічається також чутливість P. mirabilis до ампіциліну. Багато штамів P. morganii чутливі до тетрацикліну і нітрофуроновим препаратам [3].
9. ПАТОГЕНЕЗ БАКТЕРІЙ РОДИНИ ENTEROBACTERIACEAE
бактерія enterobacteriaceae патогенність токсиноутворення
Серед родини ентеробактерій є патогенні, умовно-патогенні та сапрофітні види. Патогенні представники викликають у людей, а також у тварин відмінні за патогенезом і клінічному виявленню інфекційні захворювання, які класифікуються як ешерихіози, шигельози, сальмонельози та ін.
Найбільш розповсюдженні на теперішній час захворювання людей, які викликані бактеріями які належать до родів Salmonella, Escherichia, Klebsiella, Proteus і Shigella. При цьому деякі види і біовари цих родів викликають внутрішньолікарняні інфекції, особливо серед дітей молодшого віку [3].
Ешерихії
Кишкові палички викликають декілька патогенних і клінічних форм інфекційного процесу. Їх патогенні особливості корелюють в певному ступіні з вмістом відповідного О-антигену, тобто з приналежністю до тієї чи іншої серогрупи. Так, наприклад, при інфекціях сечовивідних шляхів зустрічаються серогрупи О2, О6, О9 та ін., при холециститі – О1, О8, О11 та ін., при апендициті – О1, О2, О8 та ін. Крім того, кишкова паличка може викликати перитоніт, сепсис, ендотоксичний(септичний) шок, харчові токсикоінфекції.
Нерідкі випадки, коли кишкова паличка в асоціаціях з стафілококом або деякими грам негативними бактеріями (протеї, псевдомонас) є причиною внутрішньолікарняних інфекцій в хірургічних, гінекологічних лікарнях, в відділеннях для новонароджених тощо, викликаючи загниття ран і запальні процеси. Велика роль кишкової палички в виникнені гострих кишкових інфекцій, які обєднанні в групу ешерихіозів. В таких випадках встановленний ще більш тісний взаємозв’язок між певними серогрупами ешерихій і нозологічними формами інфекцій які викликані ними. До них відносяться колі-ентерити, переважно дітей молодшого віку, дизентерієподібні і холероподібні захворювання дітей і дорослих. При колі-ентеритах дітей молодшого віку частіше за все виділяють ентеропатогенні ешерихії серогруп О26, О55, О111 та ін., при дизентерієподібних захворюваннях – О25, О124, О144 та ін., при холероподібних інфекціях – ентеротоксигенні ешерихії О1, О15, О78, О148 та ін.
Збудники колі-ентеритів і холероподібних захворювань розмножуються на поверхні епітеліальних клітин кишечника, дизентерієподібних захворювань – всередині епітеліальних клітин, як і шигели. При руйнуванні бактерій вивільняється ендотоксин. Він володіє пірог енними властивостями, в великих дозах пригнічує фагоцитоз. Збудники холероподібних захворювань – ентеротоксигенні ешерихії утворюють секретуємі токсини білкової природи – ентеротоксини, які розрізняються за чутливістю до температури та іншим ознакам. Так, термолабільний ентеротоксин, близький до холеро гену стимулює активність аденілциклази і викликає накопичення цАМФ (циклічний аденозінмонофосфат), що призводить до порушення секреції і розвитку гострої діареї. Термостабільний ентеротоксин активує гуанілциклазу і викликає утворення цГМФ [3].
Сальмонели
Сальмонели − збудники черевного тифу і паратифів А і В (S. typhi, S. parathyphi A, S. schottmuelleri)
Через ротову порожнину і стравохід сальмонели разом з харчами потрапляють в шлунок, де багато з них гинуть. Бактерії які збереглись поступають в тонкий кишечник. Вони руйнують епітелій і проникають через нього в власний шар слизової оболонки, локалізуються і розмножуються в макрофагах лімфатичної тканини пейерових бляшок і солітарних фолікулів. В кінці інкубаційного періоду, який триває приблизно 10-14 днів, сальмонели поступають в лімфу і кров, викликаючи бактеріємію, яка триває на протязі всього гарячкового періоду. З кров’ю вони розносяться по організму і локалізуються в лімфатичній тканині внутрішніх органів і жовчному міхурі. Накопичуючись в жовчі, сальмонели разом з нею в великих кількостях вторинно потрапляють в тонкий кишечник і знову інфікують вже сенсибілізовану лімфатичну тканину, що супроводжується утворенням специфічних черевнотифозних гранулем і виразок.
В результаті фагоцитозу і лізису сальмонел вивільняється ендотоксин, надходження якого в кров і викликає інтоксикацію організму. З кінця 2-го і початку 3-го тижня хвороби, сальмонели, що надійшли в просвіт кишечника, виводяться із організму з фекаліями і сечею. Бактеріоносійство не закінчується в період клінічного видужування і може продовжуватися багато місяців, а іноді і роки. Це пояснюється розвитком хронічного процесу в жовчному міхурі. В ньому сальмонели знаходять сприятливі умови для свого розмноження. Має значення також недостатня напруженість постінфекційного імунітету [3].
Сальмонели – збудники харчових токсикоінфекцій
Вирішальне значення для виникнення харчових токсикоінфекцій має кількість сальмонел, які з харчами потрапляють в шлунково-кишковий тракт. При масовій загибелі цих бактерій в травному тракті відбувається вивільнення ендотоксину. Ендотоксин потрапляти в організм людини з м’ясом або іншими харчовими продуктами, які були заражені сальмонелами. Із шлунково-кишкового тракту ендотоксин потрапляє в кров. Він володіє пірогенними та іншими властивостями, які викликають клінічний синдром гострого гастроентериту і загального токсикозу, який розвивається через декілька годин після прийому заражених харчів. Разом з тим ендотоксин порушує захисний бар’єр слизової оболонки кишечника і його лімфатичного апарату, що сприяє тим самим проникнень бактерій в кров, де вони іноді можуть бути виявлені вже в перші години. Захворювання триває не більше 4-5 днів [3].
Сальмонели – збудники внутрішньолікарняних хвороб
Збудником внутрішньолікарняного сальмонельозу частіше за все є S. typhimurium. Однак не рідкі захворювання які викликані S. derby, S. heidelberg, S. wien, S. haifa та ін., які відносяться до групи В. Ці сальмонели за своїми морфологічними, фізіологічними, біохімічними і антигенними ознаками не відрізняються від бактерій – збудників харчових токсикоінфекцій. В теперішній час ідентифіковані біовари деяких з перелічених сальмонел, які, як правило, виділяють лише при внутрішньолікарняній інфекції.
Так, серед S. typhimurium ідентифіковано три біовари, однакових за своєю антигенною структурою, але вони відрізняються один від одного за патогенністю для білих мишей при ентеральному зараженні і за чутливістю до антибіотиків. Як правило, сальмонели, які виділяються при внутрішньолікарняній інфекції, резистентні до 15-20 антибіотиків. Це пов’язано з наявністю в них кон’югативних R-плазмід, що несуть стійкість до антибіотиків [3].
Шигели
Патогенез дизентерії визначається адгезивними властивостями, проникненням збудника в клітини епітелію товстого кишечника, внутрішньоклітинним розмноженням і токсигенністю. Зараження людей шигелами відбувається фекально-оральним шляхом. Велике значення при цьому має величина інфекційної дози. Для дизентерії характерні водний, харчовий, а також побутовий шляхи передачі інфекції, особливо серед дітей дошкільного віку. Дизентерія Зонне часто передається харчовим шляхом. Шигели Зонне здатні більш інтенсивно, ніж інші види шигел, розмножуватися в харчових продуктах і в кишечнику і утворювати ентеротоксин, що визначає можливість виникнення харчової токсикоінфекції [3].
Клебсієли
Клебсієли є збудниками різних захворювань людини. Вони викликають захворювання дихальних шляхів, а також вражають слизову оболонку сечостатевого тракту, повік, а в ряду випадків мозкової оболонки, суглобів. Крім того, клебсієли можуть викликати кишкові інфекції у дорослих та дітей і гнійні післяопераційні ускладнення, у новонароджених – важкі пневмонії, кишкові інфекції і токсикоз-септичні стани з летальним випадком. В останні десятиріччя антибіотикорезистентні штами клебсієл не рідко є збудниками внутрішньолікарняних інфекцій. Джерело інфекції – хворі та бактеріоносії. Зараження відбувається через респіраторні шляхи. Для пневмонії, яка викликана K. pneumoniae, характерні утворення багато чисельних осередків в дольках легеня з наступним їх злиттям і ослизненням враженої тканини, яка містить велику кількість клебсієл. Можливо утворення гнійних осередків в інших органах і розвиток сепсису.
При склеромі, яка викликана K.rhinoskleromatis, вражається слизова оболонка носу (риносклерома), носоглотки, гортані, трахеї, бронхів. В тканинах утворюються гранулеми з наступним склеротичними змінами. Хвороба протікає хронічно, з поступовим розповсюдженням хрящовидних інфільтратів з носової порожнини на гортань. Смерть може наступити внаслідок звуження просвіту дихальних шляхів.
При озені, яка викликається K. ozaenae, вражається слизова оболонка носу і придатних порожнин з наступною атрофією носових раковин і виділенням зловонного секрету. При підсиханні утворюються кірки, які перешкоджають диханню [3].
Протеї
Протеї є збудниками харчових токсикоінфекцій, в виникненні яких суттєве значення має кількість бактерій., що потрапляють аліментарним шляхом в шлунково-кишковий тракт. Руйнування бактеріальних клітин з вивільненням великої кількості ендотоксину грає головну роль в патогенезі цього захворювання, що протікає аналогічно харчовим токсикоінфекціям, які викликані ентеробактеріям. P. morganii викликає диспепсію у дітей, а також гнійні запальні процеси (цисти, кон’юнктивіти, отити). В останні десятиріччя відмічається збільшення числа септицемій, які викликані протеєм.
Протей в асоціаціях з іншими грам негативними бактеріями (E. coli, Pseudomonas), стафілококами або стрептококами, а також з анаеробними клострідіями ускладнює протікання гнійної і анаеробної інфекції.
P.rettgeri і P. morganii можуть бути причиною внутрішньолікарняних інфекцій [3]
10. ІМУНІТЕТ ДО БАКТЕРІЙ РОДИНИ ENTEROBACTERIACEAE
Ешерихії
При колі-бактеріозах, які проявляються гнійними запальними процесами, часто ослаблюється імунна система організму, пригнічується фагоцитарна реакція, що призводить до порушення природних бар’єрів і розповсюдженню збудника.
В імунітеті при ешерихіозах провідну роль відіграють:
1) транс плацентарна передача антитіл (IgG) і їх проникнення із крові дитини в кишечник;
2) пасивна ентеральна імунізація антитілами материнського молока;
3) продукція секреторних антитіл SIgA лімфатичними клітинами кишечника.
Імунітет дітей молодшого віку при дизентерієподібні колі-інфекції принципово відрізняється від такого при колі-ентериті. При цьому основне значення мають отриманні від матері IgG, які дифундують із крові в просвіт кишечника. В той же час ці антитіла не запобігають дітей від колі-ентериту, а антитіла, які містяться, в крові матері IgM не здатні подолати плацентарний бар’єр. Цим пояснюється те, що діти перших місяців життя не сприймають ешерихії, які викликають дизентерієподібну колі-інфекцію, і шигелам і висока чутливість до збудників колі-ентеритів.
Утворення місцевого імунітету в кишечнику немовлят, а в деяких випадках і в більш дорослих дітей пов’язано з секреторними антитілами SIgA, які містяться в грудному молоці. Пасивна ентеральна імунізація цими антитілами, а також продукція секреторних антитіл аналогічного класу лімфатичними клітинами кишечника сприяє покриттю ними слизової оболонки кишечника, що перешкоджає „прилипанню” до неї бактерій. Діти, що знаходяться на штучному годуванні і які не отримали достатньої кількості IgM і IgA, більш сприятливі до колі-ентеритів [3].
Сальмонели − збудники черевного тифу і паратифів А і В(S. typhi, S. parathyphi A, S. schottmuelleri)
Постінфекційний імунітет характеризується високою напруженістю і тривалим зберіганням. Повторні захворювання спостерігаються рідко.
До кінця 1-го тижня хвороби в крові з’являються аглютиніни, преципітини, комплементзв’язуючи антитіла, бактеріолізини. Під впливом цих антитіл відбувається загибель бактерій з вивільненням ендотоксинів і розвитком основних симптомів захворювання. Кількість антитіл поступово збільшується і досягає максимуму на 14-15-й день захворювання [3].
Сальмонели – збудники харчових токсикоінфекцій
Постінфекційний імунітет не тривалий і не володіє достатньою напруженістю. В сироватці крові хворих і реконвалесцетів виявляються аглютиніни, преципітини, бактеріолізини та інші антитіла. Захворювання, що викликані одними сироварами, не створюють не сприйнятливість до інших, а перенесена інфекція не виключає реінфекцію [3].
Шигели
При дизентерії розвивається місцевий ы загальний імунітет. При місцевому імунітеті суттєве значення має секреторний IgA (SIgA), прикріплення шигел до епітеліальних клітин і проникнення в них. SigF утворюється в 1-й тиждень захворювання в лімфатичних клітинах слизової оболонки кишечника. Покриваючи слизову оболонку кишечника, ці антитіла перешкоджають прикріпленню і перенетрації шигел в епітеліальні клітини.
Крім того, в процесі інфекції зростає титр сироваткових антитіл IgM, IgA, IgG, який досягає максимуму на 2-му тижні захворювання. Найбільша кількість IgM виявляється в 1-й тиждень хвороби.
Наявність специфічних сироваткових антитіл не є показником напруженості місцевого імунітету [3].
Клебсієли
В імунітеті при інфекціях, які викликані клебсієлам, має значення фагоцитоз опсонізованих специфічними антитілами клебсієл. Внутрішньоклітинна локалізація збудника сприяє розвитку хронічних форм інфекції. При захворюваннях, які викликані клебсієлам, розвивається гіперчутливість уповільненого типу.
В процесі інфекцій накопичуються антитіла, які не мають суттєвого значення в імунітеті [3].
11. ЛАБОРАТОРНА ДІАГНОСТИКА БАКТЕРІЙ РОДИНИ ENTEROBACTERIACEAE
Мікробіологічну діагностику кишкових інфекцій проводять так. Основою її є виділення чистих культур ідентифікація їх до виду за допомогою визначення біохімічних властивостей та реакції аглютинації з відповідними антисироватками. Матеріалом для дослідженим найчастіше служать випорожнення, блювотні маси, кров, гній, ліквор, сеча, жовч, дуоденальний вміст, секційний матеріал.
Посіви проводять на
середовище Ендо, Плоскирєва, Левіна,
вісмут-сульфітний агар з наступною
мікроскопією колоній, пересівом їх на
середовище Олькеницького та строкатий
ряд Гісса, постановкою реакції аглютинації
з О- і Н-антисироватками. В ряді випадків
застосовують тести, що дають можливість
надійно встановити вид виділеної
культури. Дуже важливо проводити й
кількісне дослідження, що дозволяє
точніше встановиш збудника захворювання.
При патологічних процесах, викликаних
умовно-патогенними бактеріями,
концентрація справжнього збудника, як
правило, становить 10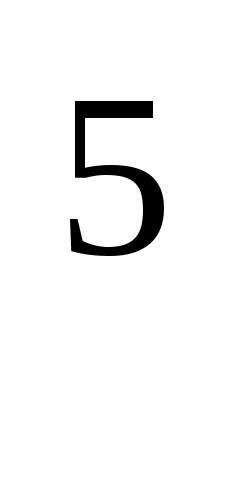 -10
-10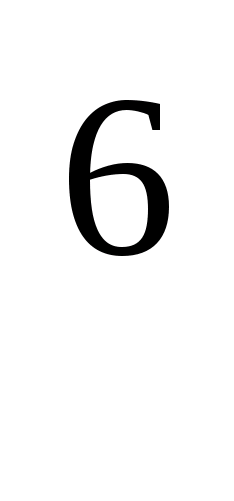 клітин в 1 мл досліджуваного матеріалу.
Це особливо важливо при виділенні
мікробних асоціацій [6].
клітин в 1 мл досліджуваного матеріалу.
Це особливо важливо при виділенні
мікробних асоціацій [6].
Ешерихії
Мікробіологічна діагностика ешерихіозів має особливо важливе значення, оскільки їх клінічні прояви характеризуються відсутністю патогномонічних симптомів відносно збудника. Провідним залишається бактеріологічний діагноз. Його особливість полягає в тому, що виділення й ідентифікація збудника базується на визначенні його антигенної структури, а не на вивченні біохімічних властивостей.
Взяття матеріалу для дослідження. До початку етіотропного лікування у хворих беруть випорожнення, блювотні маси, дуоденальний вміст, кров, сечу, гній, спинномозкову рідину, від трупів - кров із серця, вміст кишок, шматочки легень, печінки, селезінки, нирок. При необхідності досліджують промивні води шлунка, залишки їжі, змиви з рук обслуговуючого персоналу, повітря палат тощо.
Бактеріоскопічне дослідження. В окремих випадках роблять первинну мікроскопію крові, сечі, ліквору, гнійних виділень, секретів слизових оболонок у мазках, забарвлених за Грамом. Виявлення грамнегативних паличок допомагає бактеріологові вибрати відповідні живильні середовища і наступні етапи лабораторної діагностики. Початкова бактеріоскопія випорожнень не проводиться [5].
Бактеріологічне дослідження, супроводжується виділенням чистої культури E. coli. Виділення чистої культури супроводжується певними труднощами. Вони пов’язані з наявністю у досліджуваному матеріалі (фекалії) звичайних ешерихій, представників нормальної мікрофлори кишечника. Ці бактерії разом з ентеропатогенними штамами утворюють однотипні колонії на диференційно-діагностичних середовищах [3].
Серологічне дослідження. Виявлення антиешерихіозних аглютинінів у сироватці крові хворих з діагностичною метою в рутинній лабораторній практиці не знайшло широкого використання. Антитіла якщо й виявляються, то в низьких титрах (не вище 1:100). Для серологічної діагностики колі-інфекцій більш чутливою і специфічною є реакція непрямої гемаглютинації (РИГА). За допомогою РИГА можна виявити лише О-антитіла, що може бути використано для диференціації захворювань від бактеріоносійства, особливо, якщо взяти для реакції еритроцитарний діагностикум з автоштамом.
Достовірність серологічної діагностики ешерихіозів зростає при виявленні окремих класів імуноглобулінів. Зміна підвищеної концентрації ІgМ на ІgG обумовлена гострим інфекційним процесом. Виявлення в сироватці крові лише класу ІgG свідчать про бактеріоносійство.
Отже, лабораторна діагностика основана на виділенні збудника захворювання і наступній ідентифікації патогенних і непатогенних штамів кишкових паличок за допомогою діагностичних ОК-сироваток в орієнтованій і розгорнутій реакції аглютинації [6].
Сальмонели
Сальмонели − збудники черевного тифу і паратифів А і В(S. typhi, S. parathyphi A, S. schottmuelleri)
В основі лабораторної діагностики тифо-паратифозних захворювань лежать патогенетичні особливості цих інфекцій, що пов’язані з локалізацією збудника в лімфатичній тканині внутрішніх органів, крові, жовчі і виділенням його з випорожненням і сечею [3].
Взяття матеріалу для дослідження. Важливе значення для успішного проведення лабораторної діагностики черевного тифу і паратифів має правильний і своєчасний забір досліджуваного матеріалу залежно від фази патогенезу і строків черевнотифозного захворювання. Мікробіологічні дослідження при паратифах проводять так само, як і при черевному тифі. Досліджуваний матеріал для виділення чистої культури збудника, по можливості, слід брати до початку антибіотикотерапії. Найчастіше беруть кров, кістковий мозок, дуоденальний вміст (жовч), ексудат із розеол, випорожнення, сечу, гній, спинномозкову рідину, секційний матеріал при летальних випадках.
Бактеріологічні методи дослідження. Для ранньої діагностики черевного тифу і паратифів найефективнішим є виділення збудника з крові й кісткового мозку, в меншій мірі з жовчі, сечі, випорожнень та інших досліджуваних матеріалів. Висів паличок черевного тифу і паратифів із кров'яного русла чи кісткового мозку має абсолютну, 100 % діагностичну цінність.
Бактеріологічні методи поділяються на методи гемокультури, мієлокультури, білікультури, розеолокультури, уринокультури, копкокультури.
Більш надійною є серологічна ідентифікація виділених культур в реакції аглютинації з діагностичними сироватками. Спочатку реакцію ставлять на склі з адсорбованими аглютинуючими сироватками, які містять антитіла до антигенів 09 (S. typhi), 02 (S. parathyphi A) і 04 (S. schottmuelleri). Якщо виділена культура за біохімічними властивостями подібна до тифозної, але не аглютинується 09-сироваткою, її необхідно проаглютинувати з Vі-сироваткою.
Фаготипування виділених культур. Важливе епідеміологічне значення, особливо для встановлення джерела інфекції, має фаготипування тифо-паратифозних мікробів. Збудники черевного тифу з Vі-антигеном лізуються Vі-бактеріофагами. їх нараховують 86 типів. Всі вони високоспецифічні. Є набори фагів і для типування паратифозних сальмонел [6].
Сальмонели – збудники харчових токсикоінфекцій
Взяття досліджуваного матеріалу. Від хворих на сальмонельоз забирають блювотні маси, промивні води шлунка, випорожнення, кров (у перші години захворювання при підозрі на бактеріємію), кістковий мозок, жовч, сечу, спинномозкову рідину. Для виявлення бактеріоносіїв серед працівників підприємств громадського харчування, водопостачання та дитячих закладів досліджують фекалії після прийому проносного. При розтині трупів беруть вміст шлунка і кишок, кров із серця, шматочки паренхіматозних органів, лімфатичні вузли брижі.
При діагностиці харчових токсикоінфекцій обов'язково беруть також залишки підозрілої їжі, продукти, з яких її готу вали, змиви з поверхні столів, кухонних дощок, рук обслуговуючого персоналу тощо.
До лабораторії матеріали доставляють в упакованому та опечатаному вигляді. При неможливості швидкої доставки їх зберігають при 4-6 °С не більше доби [6].
Остаточний діагноз харчової токсикоінфекції ставлять тільки після виділення збудника із організму хворих людей і харчових продуктів. Для цього проводять бактеріологічне дослідження [3].
Також проводять серологічну діагностику. У тих випадках, коли при наявності клінічних симптомів, характерних для сальмонельозної токсикоінфекції, мікробіологічне дослідження не проводилось, або збудник не виділено, ставлять реакцію аглютинації з сироваткою крові перехворілих. Серологічні дослідження проводять також з метою ретроспективного аналізу масових захворювань на підприємствах громадського харчування і в організованих колективах [6].
Сальмонели – збудники внутрішньолікарняних хвороб
Основне значення має виділення чистої культури і визначення її серогрупи, серовару і біовару при інфекції S. schottmuelleri [3].
Шигели
Основним методом мікробіологічної діагностики дизентерії є бактеріологічний. Схема виділення збудника класична: посів матеріалу на середовище збагачення та агар Плоскирєва, одержання чистої культури, вивчення її біохімічних властивостей та ідентифікація за допомогою полівалентних і моновалентних аглютинуючих сироваток.
Взяття матеріалу для дослідження. Позитивний результат мікробіологічного аналізу значною мірою залежить від своєчасного і правильного забору досліджуваного матеріалу. У хворих і бактеріоносіїв найчастіше беруть випорожнення, значно рідше − блювотні маси і промивні води шлунка та кишок. Фекалії (1 − 2 г) беруть скляною паличкою із судна або пелюшок, включаючи шматочки слизу і гною (але не крові). Краще всього для дослідження взяти слиз (гній) із місць ураження слизової оболонки під час колоноскопії.
Збудники дизентерії дуже рідко проникають в кров і сечу, у зв'язку з чим ці об'єкти звичайно не сіють. Бактеріологічний аналіз секційного матеріалу необхідно проводити якомога скоріше після смерті (товстий кишечник, мезентеріальні лімфатичні вузли, шматочки паренхіматозних органів). При спалахах дизентерії досліджують також харчові продукти, особливо молоко, сир, сметану.
Серологічна ідентифікація виділених культур проводиться за допомогою реакції аглютинації на склі спочатку з сумішшю сироваток проти видів Флекснера і Зонне, які найчастіше зустрічаються, а потім із моновидовими та монорецепторними сироватками. Останнім часом випускають комерційні як полівалентні, так і моновалентні сироватки проти всіх видів збудників дизентерії.
З метою швидкої і надійної ідентифікації шигел ставлять також пряму й непряму реакції імунофлуоресценції та ензиммічених антитіл. Остання при дизентерії є високоспецифічною і все частіше використовується при лабораторній діагностиці захворювання.
Серологічну діагностику дизентерії проводять рідко. Інфекційний процес не супроводжується значним антигенним подразненням, тому титри антитіл у сироватці хворих і реконвалесцентів невисокі, їх виявляють на 5-8 добу захворювання. Найбільше антитіл утворюється на 2-3-му тижні [6].
Клебсієли
Основним методом лабораторної діагностики клебсієльозів є бактеріологічне дослідження. Серологічний метод застосовують рідко. Проводять також мікроскопію мазків.
Матеріалом для мікробіологічного аналізу може бути харкотиння, слиз із рото- і носоглотки, промивні води і блювотні маси, гній, кров, ліквор, сеча, жовч, випорожнення, секційний матеріал, інфільтрати слизової при риносклеромі, кірочки при озені, змиви з предметів тощо.
Бактеріологічне дослідження проводять за такою ж схемою, що й при подібних захворюваннях, викликаних іншими видами ентеробактерій. Визначають також чутливість виділених культур до антибіотиків за допомогою дискодифузійного методу.
Серологічні дослідження проводять шляхом постановки розгорнутої реакції аглютинації з сироваткою крові хворих і капсульним та безкапсульним антигенами, а також зв'язування комплементу (титр 1:40 і вище) та більш чутливої і специфічної реакції непрямої гемаглютинації з еритроцитарним клебсієльозним діаг-ностикумом [6].
Протеї
Матеріал для досліду піддається бактеріоскопії і засівається на поживні середовища з метою виділення чистої культури. Ідентифікацію отриманої культури проводять на основі морфологічних і біохімічних ознак.
При наявності
діагностичних сироваток види протею
ідентифікують в реакції аглютинації.
Найважливішою ознакою, яка відрізняє
протеїв від інших ентеробактерій є їх
здатність дезамінувати внесений до
агару фенілаланін. Розклад останнього
до фенілпіровиноградної кислоти в
присутності FeCl
призводить до забарвлення середовища
в зелений колір [3,6].
12. ПРОФІЛАКТИКА І ЛІКУВАННЯ ЗАХВОРЮВАНЬ ВИКЛИКАНИХ БАКТЕРІЯМИ РОДИНИ ENTEROBACTERIACEAE
Ешерихії
Основу хіміотерапії ешерихіозів складає призначення ефективних антимікробних засобів (ампіцилін, норфлоксацин та ін.). Для лікування інфекцій сечовивідних шляхів також використовують цефалоспоріни і аміноглікозиди. Для лікування колі-інфекцій користуються антибіотиками, наприклад поліміксин, ампіцилін, тетрацикліни [3,13].
Засобів специфічної імунопрофілактики немає. Профілактика колі-інфекцій направлена на дотримання санітарно-гігієнічних правил, попередження інфікування продуктів харчування і розмноження в харчах мікроорганізмів, знищення мікробів, які потрапили за допомогою термічної обробки [13].
Сальмонели
Основу лікування складає адекватна антимікробна терапія (препарати вибору – ампіцилін, аміноглікозиди, фторхінолони та ін.) [13].
Сальмонели − збудники черевного тифу і паратифів А і В (S. typhi, S. parathyphi A, S. schottmuelleri)
Профілактика основана на проведенні ветеринарно-санітарних, санітарно-гігієнічних і противоепідемічних заходів. В теперішній час застосовується хімічна, адсорбована на гелі окису алюмінію тифо-паратифозна-стовбнячна вакцина (TABte). Вона складається з повних антигенів сальмонел черевного тифу, паратифів А і В, і стовбнячого анатоксину. Хороші результати спостерігаються при використанні вакцин, які містять Vi-антиген S. typhi.
Етіотропну терапію проводять на протязі всього гарячкового періоду, а також 10 днів після його закінчення. Патогенетичне лікування включає інфузіонно-дезінтоксикаціону терапію, екстракорпоральну детоксикацію та ін.
Для лікування користуються також левоміцитином та іншими антибіотиками [3,13].
Сальмонели – збудники харчових токсикоінфекцій
У хворих з харчовими токсикоінфекціями основний метод лікування - патогенетична терапія, що направлена на дезінтоксикацію і відновлення водно-електролітного балансу і гемодинаміки. В першу чергу слід промити шлунок звичайною питною водою або розчином соди [13].
Сальмонели – збудники внутрішньолікарняних хвороб
З метою специфічної профілактики використовують полівалентний сальмонельозний фаг. Його вводять дітям в лікарняних стаціонарах, які контактували з хворими сальмонельозами і носіями. Отримують фаг також матері, які знаходять в тісному контакті з хворими дітьми [3].
Основу лікування складає антимікробна терапія (препарати вибору – ампіцилін, норфлоксацин та ін.). При виділенні резидентних штамів лікування слід проводити з врахуванням їх чутливості. В більшості випадків необхідні сімптоматична терапія, заповнення втрат рідини і електролітів. Отримання різних вакцин (формалінізовані, хімічні, грітих) не вирішило проблему специфічної профілактики дизентерії, оскільки всі вони володіють низькою ефективністю. Основні заходи направлені на дотримання санітарно-гігієнічних правил профілактики кишкових інфекцій [3,13].
Клебсієли
Враховуючи велику стійкість клебсієл, антимікробну терапію слід починати після встановлення чутливості до препаратів. Препарати вибору – аміноглікозиди і β-лактамні антибіотики широкого спектру дії. Антимікробну терапію слід поєднувати з призначенням сімптоматичного лікування. Засоби специфічної імунопрофілактики не розроблені. Для попередження клебсієльозів слід суворо
дотримуватися правил зберігання харчових продуктів, правил асептики і антисептики в лікарняних закладах і правил особистої гігієни [13].
Протеї
Протеї володіють природною стійкістю до багатьох антибіотиків. Препарати вибору – ампіцилін, цефалоспорини третього покоління, фторхінолони. При дисбактеріозах кишечника можна призначати інтестибактеріофаг (суміш фагів, що включає протейний фаг) внутрішньо. Протейний або колі-протейний фаг також застосовуються місцево (при гнійних процесах або ураженні сечовивідної системи) [13].
13. ДИНАМІКА ЗМІНИ РЕЗИСТЕНТНОСТІ УМОВНОГО-ПАТОГЕННИХ ЕНТЕРОБАКТЕРІЙ ШЛУНКОВО-КИШКОВОГО ТРАКТУ ДО АНТИМІКРОБНИХ ПРЕПАРАТІВ
Формування резистентності мікроорганізмів до антибактеріальних препаратів є серйозною медичною проблемою. З огляду на це, здійснення постійного моніторингу за поширенням резистентних штамів, особливо умовно-патогенних мікроорганізмів, в певному регіоні дозволяє встановити місцеві особливості чутливості виділених штамів до антибіотиків.
Мета роботи полягала у дослідженні антибіотикорезистентності умовно-патогенних мікроорганізмів шлунково-кишкового тракту. Обстежено 150 хворих різних вікових груп, у яких виділено 160 штамів умовно-патогенних мікроорганізмів. Ідентифікацію мікроорганізмів проводили за морфо-тинкторіальними, культуральними та біохімічними властивостями з використанням тестів Lachema (Чехія) - Enterotest I та Enterotest II.
Чутливість до антибіотиків визначали за допомогою диско-дифузійного метода з використанням стандартних дисків фірми ТОВ «Аспект»: хлорамфенікол (30 мкг/диск), цефалексин (30 мкг/диск), цефтріаксон (30 мкг/диск), гентаміцин (10 мкг/диск) та фуразолідон (300 мкг/диск).
Встановлено, що у E.colі (гемолітичної) зростає резистентність до фуразолідону (від 30% до 60%) та спостерігається зростання і коливання показників резистентності до цефтріаксону (25%-60%-50%) та гентаміцину (40%-60%-50%), відповідно за 2005-2006-2007 роки. У E.coli (атипової) відмічено зростання резистентності до хлорамфеніколу (з 20% до 67%) та гентаміцину (з 30% до 50%). У K. рneumoniae виявлено зростання резистентності до фуразолідону (з 20% до 36%). У K. mobilis – до хлорамфеніколу (з 20% до 67%) і цефалексіну (з 40% до 67%), гентаміцину (з 50% до 67%). У P. agglomeransis – зростання і коливання резистентності до усіх вивчених антибіотиків. Таким чином найбільшу резистентність до антибіотиків виявлено у різних видів капсульних бактерій (P. agglomeransis, K. рneumoniae, K. mobilis) та гемолітичних і атипових E.colі.
За динамікою зростання резистентності умовно-патогенних мікроорганізмів шлунково-кишкового тракту можна поставити в такий ряд: гентаміцин >фуразолідон > левоміцетин > цефтріаксон та цефалексин.
ПЕРЕЛІК ПОСИЛАНЬ
1. Воробьёв А.А., Буков А.С., Пашков Е.П., Рубакова А.М. Микробиология. – 2-е изд. – М.: Медицина, 2003. – 175с.
2. http://www.djerelo.com/index.php?option=com_content&task=view&id= 12402& Itemid=536
3. Тимаков В.Д., Левашев В.С., Борисов Л.Б. Микробиология: учебник. – 2-е изд., перераб. и доб. – М.: Медицина, 1983. – 512с.
4. Пирог Т.П. Загальна мікробіологія: Підручник. – К.: НУХТ, 2004. – 471 с.
5. http://uk.wikipedia.org/wiki/Файл:EscherichiaColi_NIAID.jpg
6. Климнюк С.І., Ситник І.О., Творко М.С. та ін. Практична мікробіологія: Навчальний посібник для студентів вищих мед. Закладів ІV рівня акредитації. − Тернопіль, 2004р.
7. http://uk.wikipedia.org/wiki/Файл:SalmonellaNIAID.jpg
8. http://uk.wikipedia.org/wiki/Файл:Shigella_stool.jpg
9. http://www.medkurs.ru/img-spub/23637_pub.jpg
10. Стейниер Р., Эдельберг Э., Ингрэм Дж. Мир микробов: В 3 т. − М.: Мир, 1979. – Т.2. – 464с.
11. http://www.water.ru/bz/likbez/escherichia.shtml
12. http://humbio.ru/humbio/infect_har/0002c000.htm
13. Поздеев О.К. Медицинская микробиология. − М.: ГЭОТАР-МЕД, 2001г.
14. Загаєвський І.С. Жмурко Т.В. Ветеринарно-санітарна експертиза з основами технології переробки продуктів тваринництва. − М.: Колосся, 1983.
15. http://medsocium.com/dinam-ka-zm-ni-rezistentnost-umovno-patogennikh-enterobakter-i-shlunkovo-kishkovogo-traktu-do-antim-