Математическое моделирование естествознания
Нижегородский Государственный университет
им. Н. И. Лобачевского
Реферат на тему
Математическое моделирование естествознания
выполнил:
ст. Шишкин Тимофей
731 группа
проверил:
Антаков Р.А.
Нижний Новгород
1999г.
Структура естественнонаучных знаний
Естественные и гуманитарные науки.
Наука занимается изучением объективно существующих ( т.е. существующих независимо от чьего-либо сознания) объектов и явлений природы. Вопрос о том, существует ли окружающий нас мир сам по себе или он является продуктом деятельности разума (принадлежащего некому высшему существу или каждому конкретному индивиду) составляет суть т.н. основного вопроса философии, классически формулируемом в виде дилеммы о первичности материи или сознания. В зависимости от ответа на основной вопрос философы подразделяются на материалистов (признают объективное существование окружающего нас мира, возникшего в результате саморазвития материи), объективных идеалистов (признают объективное существование мира, возникшего как результат деятельности высшего разума) и субъективных идеалистов (считают, что окружающий нас мир не существует реально, а есть плод воображения отдельного индивида) . По-видимому невозможно дать экспериментально обоснованного ответа на основной вопрос философии, хотя большинство естествоиспытателей являются приверженцами материалистических концепций.
Все существующие научные дисциплины условно (любая классификация носит приближенный характер и неполно отражает истинную суть вещей!) разделены на две основные группы: естественнонаучные (занимаются изучением объектов природы и явлений, не являющихся продуктом деятельности человека или человечества) и гуманитарные (изучают явления объекты, возникшие как результат деятельности человека).
Предмет естествознания - факты и явления, которые воспринимаются нашими органами чувств. Задача ученого- обобщить эти факты и создать теоретическую модель, включающую законы, управляющие явлениями природы. Следует различать факты опыта, эмпирические обобщения и теории, которые формулируют законы науки. Явления, например, тяготение, непосредственно даны в опыте; законы науки, например, закон всемирного тяготения - варианты объяснения явлений. Факты науки, будучи установленными, сохраняют свое постоянное значение; законы могут быть изменены в ходе развития науки, как, скажем, закон всемирного тяготения был скорректирован после создания теории относительности.
Значение чувств и разума в процессе нахождения истины - сложный философский вопрос. В науке признается истиной то положение, которое подтверждается воспроизводимым опытом. Основной принцип естествознания гласит: знания о природе должны допускать эмпирическую проверку. Не в том смысле, что каждое частное утверждение должно обязательно эмпирически проверяться, а в том, что опыт, в конечном счете, является решающим аргументом принятия данной теории.
Естествознание в полном смысле слова общезначимо и дает “родовую” истину, т.е. истину, пригодную и принимаемую всеми людьми. Поэтому оно традиционно рассматривалось в качестве эталона научной объективности. Другой крупный комплекс наук - обществознание - напротив, всегда был связан с групповыми ценностями и интересами, имеющимися как у самого ученого, так и в предмете исследования. Поэтому в методологии обществоведения наряду с объективными методами исследования приобретает большое значение переживание изучаемого события, субъективное отношение к нему и т.п.
От технических наук естествознание отличается нацеленностью на познание, а не на помощь в преобразовании мира, а от математики тем, что исследует природные, а не знаковые системы.
Следует учитывать различие между естественными и техническими науками, с одной стороны, и фундаментальными и прикладными - с другой. Фундаментальные науки - физика, химия, астрономия - изучают базисные структуры мира, а прикладные - занимаются применением результатов фундаментальных исследований для решения как познавательных, так и социально-практических задач. В этом смысле все технические науки являются прикладными, но далеко не все прикладные науки относятся к техническим. Такие науки, как физика металлов, физика полупроводников являются теоретическими прикладными дисциплинами, а металловедение, полупроводниковая технология - практическими прикладными науками.
Однако провести четкую грань между естественными, общественными и техническими науками в принципе нельзя, поскольку имеется целый ряд дисциплин, занимающих промежуточное положение или являющихся комплексными по своей сути. Так, на стыке естественных и общественных наук находится экономическая география, на стыке естественных и технических - бионика, а комплексной междисциплинарной дисциплиной, которая включает и естественные, и общественные, и технические разделы, является социальная экология.
Уровни организации материи и иерархия естественно научных знаний.
Окружающие нас объекты природы имеют внутреннюю структуру, т.е. в свою очередь сами состоят из других объектов (яблоко состоит из клеток растительной ткани, которая сложена из молекул, являющихся объединениями атомов и т.д.). При этом естественным образом возникают различные по сложности уровни организации материи: космический, планетарный, геологический, биологический, химический, физический. Представители естественных наук, занимающиеся изучением объектов какого-либо уровня могут достичь их полного описания лишь основываясь на знаниях более “низкого” (элементарного) уровня (невозможно понять законы жизнедеятельности клетки, не изучив химизм протекающих в ней реакций). Однако реальные возможности каждого отдельного исследователя весьма ограничены (человеческой жизни недостаточно не только для того, чтобы плодотворно заниматься изучением сразу нескольких уровней, но даже заведомо не хватает на сколько-нибудь полное освоение уже накопленных знаний о каком-то одном). Из-за этого возникло деление естественно научных знаний на отдельные дисциплины, примерно соответствующие вышеперечисленным уровням организации материи: астрономию, экологию, геологию, биологию, химию и физику. Специалисты, работающие на своем уровне, опираются на знания смежных наук, находящихся ниже по иерархической лестнице. Исключение составляет физика, находящаяся на “самом нижнем этаже” человеческих знаний (“составляющая их фундамент”): исторически сложилось так, что в ходе развития этой науки обнаруживались все более “элементарные” уровни организации материи (молекулярный, атомный, элементарных частиц...), изучением которых по-прежнему занимались физики.
Естественные науки различных уровней не обособлены друг от друга. При изучении высокоорганизованных систем возникает естественная потребность в информации о составляющих их элементах, предоставляемой дисциплинами “более низких” уровней. При изучении же “элементарных” объектов весьма полезны знания о их поведении в сложных системах, где при взаимодействиях с другими элементами проявляются свойства изучаемых. Примером взаимодействия наук разных уровней может служить разработка Ньютоном классической теории тяготения (физический уровень), возникшей на основе законов движения планет Кеплера (астрономический уровень), и современные концепции эволюции Вселенной, немыслимые без учета законов гравитации.
Естественные науки, находящиеся на нижних этажах иерархической лестницы, несомненно проще вышестоящих, поскольку занимаются более простыми объектами (строение электронного облака атома углерода, несомненно “проще пареной репы”, содержащей множество атомов с такими облаками!). Однако, именно из-за простоты изучаемых объектов науки нижних уровней сумели накопить гораздо больше фактической информации и создать более законченные теории.
Место математики среди естественных наук.
Обсуждавшаяся выше структура естествознания не содержит математики, без которой невозможна ни одна из современных точных наук. Это связано с тем, что сама математика не является естественной наукой в полном смысле этого понятия, поскольку не занимается изучением каких-либо объектов или явлений реального мира. В основе математики лежат аксиомы, придуманные человеком. Для математика не имеет решающего значения вопрос, выполняются ли эти аксиомы в реальности или нет (напр. в настоящее время благополучно сосуществует несколько геометрий, основанных на несовместных друг с другом системах аксиом).
Если математика заботит лишь логическая строгость его выводов, делаемых на основе аксиом и предшествующих теорем, естествоиспытателю важно, соответствует ли его теоретическое построение реальности. При этом в качестве критерия истинности естественнонаучных знаний выступает эксперимент, в ходе которого осуществляется проверка теоретических выводов.
В ходе изучения свойств реальных объектов часто оказывается так, что они приближенно соответствуют аксиоматике того или иного раздела математики (напр. положение небольшого тела можно приближенно описать, задав три его координаты, совокупность которых можно рассматривать как вектор в трехмерном пространстве). При этом ранее доказанные в математике утверждения (теоремы) оказываются применимыми к таким объектам.
Кроме сказанного, математика играет роль очень лаконичного, экономного и емкого языка, термины которого применимы к внешне совершенно разнородным объектам окружающего мира (вектором можно назвать и совокупность координат точки, и характеристику силового поля, и компонентный состав химической смести, и характеристику экономико-географического положения местности).
Очевидно, что более простые объекты нашего мира удовлетворяют более простым системам аксиом, следствия из которых математиками изучены более полно. Поэтому естественные науки “низших” уровней оказываются более математизированными.
Опыт развития современного естествознания показывает, что на определенном этапе развития естественно научных дисциплин неизбежно происходит их математизация, результатом которой является создание логически стройных формализованных теорий и дальнейшее ускоренное развитие дисциплины.
Приближенный характер естественнонаучных знаний.
Несмотря на то, что естественные науки часто называют точными, практически любое конкретное утверждение в них носит приближенный характер. Причиной этого является не только несовершенство измерительных приборов, но и ряд принципиальных ограничений на точность измерений, установленных современной физикой. Кроме того, практически все реально наблюдаемые явления столь сложны и содержат такое множество процессов между взаимодействующими объектами, что их исчерпывающее описание оказывается не только технически невозможным, но и практически бессмысленным (человеческое сознание способно воспринять лишь весьма ограниченный объем информации). На практике исследуемая система сознательно упрощается путем ее замены моделью, учитывающей только самые важные элементы и процессы. По мере развития теории модели усложняются, постепенно приближаясь к реальности.
Основные этапы развития естествознания могут быть выделены, исходя из различных соображений. По мнению автора, в качестве основного критерия следует рассматривать доминирующий среди естествоиспытателей подход к построению их теорий. При этом оказывается возможным выделение трех основных этапов.
Естествознание древнего мира. Завершенного деления на дисциплины не существовало, создаваемые концепции в своем большинстве носили мировоззренческий характер. Экспериментальный метод познания в принципе допускался, но роль решающего критерия истинности эксперименту не отводилась. Верные наблюдения и гениальные обобщающие догадки сосуществовали с умозрительными и часто ошибочными построениями.
Классический период развития естествознания берет свое начало с экспериментальных работ Галилея (18 век) и длится до начала нашего столетия. Характеризуется четким разделением наук на традиционные области и даже несколько гипертрофированной ролью эксперимента в их развитии (“понять- значит измерить”). Эксперимент рассматривается не только как критерий истинности, но и как основной инструмент познания. Вера в истинность экспериментально добытых результатов столь велика, что их начинают распространять на новые области и проблемы, где соответствующей проверки не производилось. При обнаружении расхождений так создаваемых концепций с реально наблюдаемыми явлениями неизбежно возникало недоумение, граничащее с попытками отрицания самой возможности познания окружающего мира.
Современное естествознание характеризуется лавинообразным накоплением нового фактического материала и возникновением множества новых дисциплин на стыках традиционных. Резкое удорожание науки, особенно экспериментальной. Как следствие - возрастание роли теоретических исследований, направляющих работу экспериментаторов в области, где обнаружение новых явлений более вероятно. формулировка новых эвристических требований к создаваемым теориям: красоты, простоты, внутренней непротиворечивости, экспериментальной проверяемости, соответствия (преемственности). Роль эксперимента, как критерия истинности знания, сохраняется, но признается , что само понятие истинности не имеет абсолютного характера: утверждения, истинные при определенных условиях, при выходе за границы, в рамках которых проводилась экспериментальная проверка, могут оказаться приближенными и даже ложными. Современное естествознание утратило присущую классическим знаниям простоту и наглядность. Это произошло главным образом из-за того, что интересы современных исследователей из традиционных для классической науки областей переместились туда, где обычный “житейский” опыт и знания об объектах и происходящих с ними явлениях в большинстве случаев отсутствуют.
После триумфа классической механики Ньютона химия в лице Лавуазье, положившего начало систематическому применению весов, встала на количественный путь, а вслед за ней и другие естественные науки. “Таково первое основание, по которому физик не может обойтись без математики; она дает ему единственный язык, на котором он в состоянии изъясняться (А.Пуанкаре.Цит.соч. - С.220)”.
Дифференциальное и интегральное исчисление хорошо подходит для описания изменения скоростей движений, а вероятностные методы - для необратимости и создания нового. Все можно описать количественно, и тем не менее, остается проблемой отношение математики к реальности. По мнению одних методологов, чистая математика и логика используют доказательства, но не дают нам никакой информации о мире ( почему Пуанкаре считал, что законы природы конвенциональны), а только разрабатывают средства его описания. Однако, еще Аристотель писал, что число есть промежуточное между частным предметом и идеей, а Галилей полагал, что Книга Природы написана языком математики.
Не имея непосредственного отношения к реальности, математика не только описывает эту реальность, но и позволяет, как в уравнениях Максвелла, делать новые интересные и неожиданные выводы о реальности из теории, которая представлена в математической форме. Как же объяснить непостижимую истинность математики и ее пригодность для естествознания? Может все дело в том, что “ механизм математического творчества, например, не отличается существенно от механизма какого бы то ни было иного творчества”( А.Пуанкаре.Цит.соч.-С.285)? Или более пригодны более сложные, системные объяснения?
По мнению некоторых методологов, законы природы не сводятся к написанным на бумаге математическим соотношениям. Их надо понимать как любой вид организованности идеальных прообразов вещей, или пси-функций. Есть три вида организованности: простейший - числовые соотношения; более сложный - ритмика 1-го порядка, изучаемая математической теорией групп; самый сложный - ритмика 2-го порядка - “слово”. Два первых вида организованности наполняют Вселенную мерой и гармонией, третий - смыслом. В рамках этого объяснения математика занимает свое особое место в познании.
Работа над любой математической моделью начинается со сбора и анализа фактического материала. Определяются цели моделирования. Выделяются главные черты изучаемого объекта или явления. Вводятся формализованные характеристики. Принимаются правила работы с ними. В результате возникает математический объект, который и называется математической моделью. Разрабатываются методы математического анализа модели, которыми она исследуется. Полученные результаты математического моделирования интерпретируются в рамках исходного фактического материала, что позволяет оценить степень адекватности модели. Результаты моделирования не должны противоречить выделенным ранее ключевым экспериментальным фактам. Одновременно, модель не может объяснить все стороны изучаемого объекта или явления.
Хорошая модель, кроме объяснения известных, должна давать возможность предсказывать новые свойства. Математическое моделирование широко используется там, где экспериментальные исследования трудоемки и дорогостоящи, или вообще невозможны (например, в изучении социальных явлений).
Кроме задачи о прогнозе, математическое моделирование помогает классифицировать и систематизировать фактический материал, увидеть существующие связи в мозаике фактов. Это вытекает из того, что модель является специфическим -ярким и выразительным языком, предназначенным для описания для описания изучаемого объекта или явления.
Мир математических моделей разнообразен. Существуют различные схемы их классификации. Однако, каждая модель конкретна и предназначена для описания достаточно узкого круга объектов и явлений. Модели сильно отличаются друг от друга не только предметными областями, но математической терминологией, а также математическими методами их исследования.
Предпочтение отдается более простым моделям. Отметим, что “простота” (иногда в ущерб точности)-один из принципов, о котором всегда нужно помнить при разработке математической модели.
Математические модели нейронных систем
Изучение нейронных систем - одно из самых романтических направлений научных исследований, поскольку нейронные системы присущи как человеку, так и животным. Самая совершенная интеллектуальная система - человеческий мозг. Никакой компьютер в настоящее время не может воспроизвести ее феномен. Более того, даже поведение таких относительно простых организмов, как кальмары, в настоящее время в полной мере невозможно смоделировать на компьютере. Законы функционирования отдельных элементов нервной системы в целом не плохо изучены. Однако, законы функционирования ассоциаций нельзя свести законам поведения отдельных элементов. На самом деле об эффектах, обусловленных коллективным поведением нейронных популяций, известно мало. Понятны некоторые самые общие принципы. Например, нейронные системы способны адаптироваться к меняющимся условиям, т.е. им не нужны жесткие программы. Одновременно, последние, хотя бы в форме рефлексов, присутствуют в нервной системе. Экспериментальное изучение эффектов коллективного поведения нейронных систем затруднено. Эти системы слишком сложно устроены. Так в мозге человека и животных каждый нейрон находится под воздействием тысяч других нейронов и, соответственно, влияет на тысячи нейронов. Всего же по современным оценкам в мозге порядка миллиарда нейронов. Огромное значение имеет математическое моделирование, как метод косвенного исследования. Оно помогает понять, какие процессы могут происходить в нейронных популяциях. Затем уже можно пытаться обнаружить соответствующие явления экспериментально. Модели различаются в зависимости от целей моделирования. Некоторые модели достаточно адекватно в деталях описывают поведение отдельных нейронов и помогают понять закономерности их функционирования. Они же являются базовыми для моделей малых нейронных популяций. Для описания больших популяций используют упрощенные модели нейронов. Упор делается на изучение эффектов коллективного поведения. Результаты моделирования используются как в нейрофизиологии, так и в технике. Уже сейчас выпускаются нейронные платы. Пока их возможности невелики. Они используются, например, в обработке изображений, а также при решении некоторых экономических задач. Следует отметить, что сейчас все задачи, которые можно решить с помощью нейронных плат, в принципе можно решить и с помощью обычного компьютера. Однако, нейронные платы увеличивают быстродействие. Перспективным считается направление, связанное с использованием нейронной техники для проведения вычислений. Ряд вычислений на нейроподобных системах может проводиться нетрадиционным способом -путем имитации явлений.
Возбудимые системы
Нервная клетка, или нейрон является структурной единицей нервной системы живых организмов. Индивидуальные границы нейрона, как и любой другой клетки, определяются клеточной мембраной. В любой клетке существует разность потенциалов между содержимым клетки -цитоплазмой и внеклеточной средой. Мембрана поляризована. Ее внутренняя поверхность заряжена отрицательно по отношению к внешней. Разность потенциалов называют мембранным потенциалом. Его величина составляет несколько десятков милливольт. Причина возникновения мембранного потенциала -неравенство концентраций ионов в цитоплазме и в тканевой жидкости. Это обусловлено тем, что клеточные мембраны обладают избирательной проницаемостью для различных ионов.
Живые организмы, все клетки и, в частности, нейроны обладают раздражимостью -способностью реагировать на различного рода воздействия. Реакции носят сложный, комплексный характер, но всегда сопровождаются изменением электрического заряда мембраны. Воздействия, вызывающие реакции, называются раздражителями, или стимулами. Хотя живые организмы состоят из клеток, реакции организма в целом не сводятся к реакциям отдельных клеток. Напомним универсальное правило: закономерности системы не могут быть сведены к закономерностям отдельных элементов.
Нервные клетки и некоторые ткани (мышечная, железистая) специально приспособлены к осуществлению быстрых реакций на раздражение. Такие клетки и ткани называются возбудимыми, а их реакция -возбудимостью. Клетки при возбуждении переходят из состояния физиологического покоя в состояние активности. В частности, нервная клетка генерирует один, или несколько электрических импульсов. Это связано с тем, что при возбуждении проницаемость мембраны для ионов резко увеличивается. Возникают ионные токи, что естественно приводит к изменению разности потенциалов на поверхности мембраны. Это изменение мембранного потенциала называется потенциалом действия. Генерируемые нервными клетками импульсы также называются спайками. Группы, тесно следующих друг за другом спайков, называются пачками или берстами. Амплитуды спайков относительно стабильны для каждого типа нейронов, поэтому часто говорят, что нейрон реагирует на раздражение по закону “все, или ничего”.
Нервное возбуждение - распространяющийся процесс. Возникнув в одном, или нескольких нейронах, оно передается другим клеткам. Проведение возбуждения обусловлено тем, что потенциал действия, возникнув в одной клетке, становится раздражителем для связанных с ней клеток. То же самое относится к другим возбудимым тканям, например, к сердечной мышце. Согласно наиболее распространенной в настоящее время точки зрения считается, что “сообщения” в нервной системе передаются в виде импульсов “все, или ничего” в форме частотного кода.
Обсудим природу мембранного потенциала. Мембрана, отделяющая цитоплазму нейрона от межклеточной среды, состоит в основном из липидов -жироподобных веществ. В нее встроены белковые молекулы. Некоторые молекулы пронизывают липидный слой насквозь, а другие погружены лишь до сердцевины (своеобразные ворсинки). Некоторые белки выполняют роль каналов, осуществляющих транспорт ионов через мембрану. Другие -служат рецепторами, позволяющими нейрону реагировать на поступающие от других нейронов потенциалы действия.
Механизмы переноса ионов через мембрану делятся на два класса: пассивный и активный транспорт. Пассивный транспорт не требует затрат энергии. Он определяется двумя факторами. Диффузия способствует выравниванию концентрации растворов. Носители зарядов -ионы перемещаются под действием электрического поля. При активном транспорте за счет химической энергии, освобождающейся при окислении аминокислот, может осуществляться перенос ионов как в области с более высокой их концентрацией, так и против электрических сил.
Закономерности пассивного
транспорта опираются на некоторые
положения электрохимии. Как уже
говорилось, мембраны биологического
происхождения обладают избирательной
проницаемостью. Одни ионы достаточно
легко диффундируют через них. Для других
ионов мембраны практически непроницаемы.
Рассмотрим на примере положение об
электрохимическом равновесии. Пусть в
сосуд >
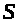 >
с раствором >
>
с раствором >
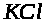 >
помещена полость >
>
помещена полость >
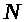 >,
ограниченная мембраной, где также
находится раствор >
>.
Обозначим через >
>,
ограниченная мембраной, где также
находится раствор >
>.
Обозначим через >
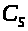 >и
>
>и
>
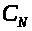 >
концентрации раствора в сосуде и в
полости. Предположим, что мембрана
проницаема для ионов >
>
концентрации раствора в сосуде и в
полости. Предположим, что мембрана
проницаема для ионов >
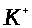 >
и непроницаема для ионов >
>
и непроницаема для ионов >
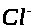 >.
Пусть концентрация раствора в сосуде
ниже, чем в полости, т.е. >
>.
Пусть концентрация раствора в сосуде
ниже, чем в полости, т.е. >
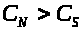 >.
Тогда ионы калия будут переходить из
полости в сосуд, поскольку диффузия
стремится выровнять концентрации. В
полости образуется относительный
дефицит положительно заряженных ионов
и она заряжается отрицательно. Точнее,
отрицательный заряд сосредотачивается
на внутренней поверхности мембраны.
Наоборот, внешняя поверхность мембраны
заряжается положительно. Электрическое
поле направлено так, что диффузия
положительных ионов >
>.
Тогда ионы калия будут переходить из
полости в сосуд, поскольку диффузия
стремится выровнять концентрации. В
полости образуется относительный
дефицит положительно заряженных ионов
и она заряжается отрицательно. Точнее,
отрицательный заряд сосредотачивается
на внутренней поверхности мембраны.
Наоборот, внешняя поверхность мембраны
заряжается положительно. Электрическое
поле направлено так, что диффузия
положительных ионов >
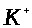 >
будет затруднена. в результате при
некоторой разности потенциалов >
>
будет затруднена. в результате при
некоторой разности потенциалов >
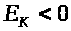 >между
внутренней и внешней поверхностью
мембраны диффузия ионов вообще
прекратится. Величина >
>между
внутренней и внешней поверхностью
мембраны диффузия ионов вообще
прекратится. Величина >
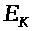 >
называется равновесным электрохимическим
потенциалом. Она вычисляется по формуле
Нернста:
>
называется равновесным электрохимическим
потенциалом. Она вычисляется по формуле
Нернста:
>
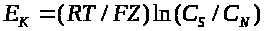 >,
(1)
>,
(1)
где >
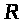 >
-универсальная газовая постоянная, >
>
-универсальная газовая постоянная, >
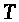 >
-абсолютная температура, >
>
-абсолютная температура, >
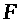 >
-число Фарадея, >
>
-число Фарадея, >
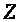 >
-заряд иона.
>
-заряд иона.
Пусть >
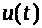 >
текущее значение мембранного потенциала.
привлечем некоторые положения теории
электрических цепей. Мембрана является
диэлектриком (изолятором) и поэтому
обладает емкостью. Ток >
>
текущее значение мембранного потенциала.
привлечем некоторые положения теории
электрических цепей. Мембрана является
диэлектриком (изолятором) и поэтому
обладает емкостью. Ток >
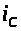 >,
текущий через емкость и напряжение >
>,
текущий через емкость и напряжение >
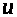 >
(оно же мембранный потенциал) связаны
соотношением: >
>
(оно же мембранный потенциал) связаны
соотношением: >
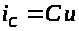 >,
где коэффициент пропорциональности >
>,
где коэффициент пропорциональности >
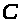 >
называется емкостью. Отметим, что в
некотором смысле ток, текущий через
мембрану, является фиктивным. Он не
сопровождается переносом зарядов.
Согласно закону Нернста мембрана
является источником напряжения >
>.
Ток >
>
называется емкостью. Отметим, что в
некотором смысле ток, текущий через
мембрану, является фиктивным. Он не
сопровождается переносом зарядов.
Согласно закону Нернста мембрана
является источником напряжения >
>.
Ток >
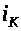 >,
текущий через источник напряжения >
>,
связан с напряжением >
>
на его полюсах формулой: >
>,
текущий через источник напряжения >
>,
связан с напряжением >
>
на его полюсах формулой: >
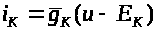 >.
По закону Кирхгофа сумма токов равна
нулю: >
>.
По закону Кирхгофа сумма токов равна
нулю: >
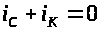 >.
Таким образом, получаем дифференциальное
уравнение:
>.
Таким образом, получаем дифференциальное
уравнение:
 .
Cu¢
+
g>K>(u
-
E>K>)
(2)
.
Cu¢
+
g>K>(u
-
E>K>)
(2)
Проведем тривиальный анализ уравнения (2). Перепишем его в виде:
>
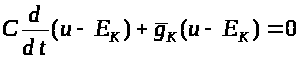 >.
>.
Отсюда получаем:
>
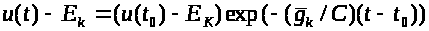 >.
>.
Таким образом, по экспоненте
мембранный потенциал >
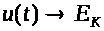 >
при >
>
при >
 >.
>.
Сделаем ряд замечаний.
При анализе уравнения (2) мы предполагали,
что коэффициенты >
>
и >
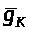 >
не зависят от >
>.
Действительно для мембраны емкость
практически постоянна. Однако, проводимость
>
>
есть функция от >
>.
Проведенный анализ справедлив лишь при
малых отклонениях >
>
не зависят от >
>.
Действительно для мембраны емкость
практически постоянна. Однако, проводимость
>
>
есть функция от >
>.
Проведенный анализ справедлив лишь при
малых отклонениях >
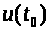 >
от равновесного значения >
>.
>
от равновесного значения >
>.
Пусть теперь дополнительно
к >
>
в полости и в сосуде присутствует >
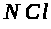 >,
концентрации которого в полости и в
сосуде различны. Ионам >
>,
концентрации которого в полости и в
сосуде различны. Ионам >
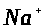 >
отвечает свое равновесное значение >
>
отвечает свое равновесное значение >
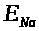 >.
Натриевый ток >
>.
Натриевый ток >
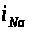 >
связан с мембранным потенциалом >
>
соотношением: >
>
связан с мембранным потенциалом >
>
соотношением: >
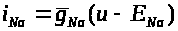 >.
Привлекая снова закон Кирхгофа, получаем:
>.
Привлекая снова закон Кирхгофа, получаем:
 .
. Cu¢
+
g>Na>(u
-
E>Na>)
+g>K>(u
-E>K>
)
(3)
.
. Cu¢
+
g>Na>(u
-
E>Na>)
+g>K>(u
-E>K>
)
(3)
Теперь в силу (3) равновесное значение мембранного потенциала суть:
>
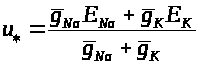 >.
>.
Одна из первых гипотез
о природе мембранного потенциала была
высказана в 1902 г. Бернштейном, который
использовал положения электрохимии.
Он предположил, что предположил, что
концентрация ионов >
>
в клетке выше, чем во внеклеточной среде.
Вследствие этого ионы >
>
диффундируют из клетки. Образуется их
относительный дефицит положительных
ионов внутри клетки и внутренняя
поверхность мембраны заряжается
отрицательно. Тем самым, потенциал покоя
определяется равновесным электрохимическим
потенциалом для ионов >
>.
Далее, Бернштейн предположил, что в
жизни нейрона иногда наступает особый
момент -состояние возбуждения, когда
на короткое время мембрана становится
проницаемой и для других ионов, в
частности, для натрия и хлора. В результате
притока внутрь клетки положительных
ионов величина мембранного потенциала
падает, а затем, после восстановления
избирательных свойств мембраны, снова
входит в норму. Так впервые была объяснена
генерация спайка.
Натриево - калиевый цикл
Гипотеза Бернштейна хорошо объясняла многие известные физиологам того времени факты. В частности, зависимость потенциала покоя от концентрации калия в межклеточной среде, а также от температуры. Однако в то время гипотезу нельзя было подвергнуть экспериментальной проверке. В распоряжении исследователей не было методики непосредственного изменения мембранного потенциала и концентраций ионов. Гипотеза Бернштейна вызывала массу споров.
Ситуация изменилась после сообщения английского зоолога Дж. Юнга, сделанного в 1936 г. Он обнаружил, что длинные тяжи у кальмаров и каракатиц являются не кровеносными сосудами, как это считалось ранее, а необычайно толстыми аксонами (отростками нервных клеток). Они получили название гигантских аксонов и стали естественным объектом для изучения мембран. Диаметр аксонов достигает 1 мм и дает возможность вводить в них электроды и капилляры. В результате можно измерить как мембранный потенциал, так и концентрацию внутриклеточного раствора.
Выяснилось, что мембранная
теория Бернштейна в целом верна, хотя
и нуждается во внесении ряда существенных
дополнений и изменений. В частности
верным оказалось утверждение, что
потенциал покоя обусловлен в
основном электрохимическим
потенциалом ионов >
>.
Наоборот, согласно теории Бернштейна
в точке максимума потенциала действия
(спайка) мембранный потенциал должен
быть равен нулю (ток ионов прекращается,
когда напряжение равно нулю). Однако,
английские физиологи А. Ходжкин и А.
Хаксли в 1939 г. показали, что на максимуме
потенциала действия мембранный потенциал
не уничтожается, а меняет знак. Внешняя
поверхность мембраны заряжается
положительно по отношению к внутренней.
Явление получило название “овершут”
и не имело объяснений в рамках теории
Бернштейна. Оно связано с неравномерным
распределением ионов натрия. Оказалось,
что концентрация ионов >
>
внутри клетки примерно в десять раз
ниже чем в межклеточной жидкости. В
начале спайка ионы натрия под действием
электрического поля и концентрационного
градиента устремляются из межклеточной
среды в клетку (в клетке их концентрация
ниже, а внутренняя поверхность мембраны
заряжена отрицательно по отношению к
внешней). В этом гипотеза Бернштейна
оказалась верной. После смены знака
мембранного потенциала ионы натрия
продолжают перемещаться в клетку только
под действием концентрационного
градиента. Мембранный потенциал на
максимуме овершута приближается к
равновесному электрохимическому
потенциалу для ионов >
>.
В дальнейшем проницаемость мембраны
для ионов натрия падает и активируется
проводимость для ионов калия, которые
движутся из клетки, где их концентрация
выше. Ионы калия уносят из клетки
положительный заряд. Внутренняя
поверхность мембраны заряжается
отрицательно. Этот натриево-калиевый
цикл ниже будет рассмотрен подробнее.
В течение цикла ионы натрия заходят в клетку, а ионы калия ее покидают. Такое замещение должно выравнивать концентрации ионов. Казалось бы, что после ряда циклов нервная клетка не должна генерировать спайков (концентрации ионов внутри и вне клетки уравняются). В действительности это не так. Неравномерное распределение ионов внутри и вне клетки поддерживается активным транспортом ионов. Механизмы, осуществляющие транспорт ионов через мембрану в сторону повышенной концентрации, называются мембранными насосами (помпами). Они потребляют энергию, которая освобождается в результате окисления аминокислот.
Наиболее изучен натриевый
насос. Он выводит ионы >
>
из клетки и одновременно транспортирует
ионы >
>
внутрь клетки. Этот процесс включает в
себя обязательный обмен двух ионов >
>
во внешней среде на три иона >
>
в клетке (если удалить из среды ионы >
>,
то ионы >
>
из клетки выводиться уже не будут). Если
мембранный потенциал отрицателен
(внутренняя поверхность мембраны
заряжена отрицательно), то натриевый
насос выполняет электрическую работу,
выводя из клетки суммарно один
положительный заряд. Натриевый насос
действует избирательно. Например, он
не способен переносить ионы лития.
Молекулярный механизм активного
транспорта в настоящее время до конца
не ясен.
Натриевый насос - один
из главных механизмов, благодаря которому
внутри клетки поддерживается избыточная
по сравнению с внешней средой концентрация
ионов >
>
и недостаточная, примерно в десять раз
меньшая, чем во внеклеточной жидкости,
концентрация ионов >
>.
Для исследования процесса генерации спайка А.Ходжкиным и А.Хаксли был разработан оригинальный прием, который называется методом фиксации потенциала. Внутриклеточно вводятся электроды и используется система с обратной связью, позволяющая поддерживать мембранный потенциал на любом заданном уровне. Тем самым, можно имитировать различные фазы потенциала действия, как бы “замораживая” его. Система позволяет измерять ионные токи. Изменяя ионный состав среды (так же через микрокапилляры меняют состав внутриклеточной жидкости) можно идентифицировать ионы, которые обуславливают ток. Исследования обнаружили, что даже пассивный транспорт ионов (в соответствии с градиентом концентраций и направлением электрического поля) подчиняется специфическим закономерностям. Для их объяснения А.Ходжкиным и А.Хаксли была высказана гипотеза о наличии в мембране специальных ионных каналов, служащих для их транспортировки.
При положительном
отклонении мембранного потенциала от
потенциала в состоянии покоя (значение
потенциала уменьшается по абсолютной
величине) активируются, т.е. открываются
каналы для >
>,
пропускающие эти ионы внутрь клетки.
Ионы >
>
движутся в соответствии с направлением
электрического поля, причем в область
с меньшей концентрацией. Каналы играют
роль селективных (избирательных)
фильтров, пропускающих ионы >
>и
непроницаемых для других ионов. Механизмы,
приводящие к открытию и закрытию каналов,
называются воротами.
При деполяризации (уменьшении абсолютной
величины мембранного потенциала)
открываются активационные так называемые
>
 >
-ворота для натриевых каналов. Чем
сильнее деполяризована мембрана, тем
больше >
>
-ворот открыто, что в свою очередь
увеличивает деполяризацию. Процесс
открытия ворот протекает лавинообразно.
>
-ворота для натриевых каналов. Чем
сильнее деполяризована мембрана, тем
больше >
>
-ворот открыто, что в свою очередь
увеличивает деполяризацию. Процесс
открытия ворот протекает лавинообразно.
При относительно длительной
деполяризации натриевые каналы
инактивируются. Срабатывают так
называемые >
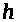 >
-ворота, которые блокируют каналы через
время порядка 1 мсек после открытия >
>
-ворот. В 1980 г. Ф. Сигворс и Э. Неер
зарегистрировали токи через единичные
натриевые каналы. Оказалось, что они
подчиняются закону “все или ничего”,
Открываются практически мгновенно, а
через время около 1 мсек. так же практически
мгновенно закрываются. Ионные каналы
представляют собой молекулы белка,
вкрапленные в липидный слой мембраны.
>
-ворота, которые блокируют каналы через
время порядка 1 мсек после открытия >
>
-ворот. В 1980 г. Ф. Сигворс и Э. Неер
зарегистрировали токи через единичные
натриевые каналы. Оказалось, что они
подчиняются закону “все или ничего”,
Открываются практически мгновенно, а
через время около 1 мсек. так же практически
мгновенно закрываются. Ионные каналы
представляют собой молекулы белка,
вкрапленные в липидный слой мембраны.
В районы максимума пика
потенциала действия большинство
натриевых каналов инактивируется, т.е.
закрываются >
>
-ворота. Встает вопрос: каким образом
мембранный потенциал возвращается к
исходному уровню -состоянию покоя?
Экспериментально А. Ходжкин и А. Хаксли
обнаружили, что с некоторым запаздыванием
по отношению к началу натриевого тока
возникает калиевый ток. Ионы калия
начинают покидать клетку, поскольку
активируются калиевые каналы. Гипотеза
связывает это с открытием так называемых
>
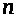 >
-ворот. Ионы калия движутся в область с
меньшей концентрацией (пассивный
транспорт). При этом в районе овершута
(мембранный потенциал положителен)
движение осуществляется в соответствии
с направлением электрического поля.
Калиевый ток уносит из клетки (точнее
с внутренней поверхности мембраны)
отрицательный заряд. Мембрана снова
поляризуется.
>
-ворот. Ионы калия движутся в область с
меньшей концентрацией (пассивный
транспорт). При этом в районе овершута
(мембранный потенциал положителен)
движение осуществляется в соответствии
с направлением электрического поля.
Калиевый ток уносит из клетки (точнее
с внутренней поверхности мембраны)
отрицательный заряд. Мембрана снова
поляризуется.
Считается, что в отличие
от натриевых каналов, для калиевых
каналов нет инактивационных ворот.
Активационные >
>
-ворота сами закрываются, когда мембрана
возвращается к уровню потенциала покоя
(реполяризуется).
Важна отметить, что
количество ионов, которое проходит
через мембрану при одиночном потенциале
действия, практически не вызывает
изменения внутриклеточных концентраций
ионов. Исключение могут составлять лишь
мельчайшие нервные клетки. По некоторым
оценкам при одном потенциале действия
содержание >
>
внутри гигантского аксона кальмара
изменяется всего лишь на >
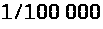 >
от исходного. Если блокировать активные
натриевые насосы каким-нибудь ядом, то
аксон все равно сможет сгенерировать
несколько тысяч импульсов. Непосредственно
активный транспорт не участвует в
генерации спайков. Однако, он поддерживает
на должном уровне концентрационные
градиенты, наличие которых приводит к
возникновению потенциалов действия.
>
от исходного. Если блокировать активные
натриевые насосы каким-нибудь ядом, то
аксон все равно сможет сгенерировать
несколько тысяч импульсов. Непосредственно
активный транспорт не участвует в
генерации спайков. Однако, он поддерживает
на должном уровне концентрационные
градиенты, наличие которых приводит к
возникновению потенциалов действия.
После описания механизма генерации спайка, обратимся к системе уравнений, описывающей это процесс. Она предложена А. Ходжкиным и А. Хаксли.
Аксон Ходжкина - Хаксли
Модель базируется на
электрохимическом уравнении (3). Мембранный
потенциал будем отсчитывать от его
уровня >
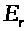 >
в состоянии покоя, т.е. положим >
>
в состоянии покоя, т.е. положим >
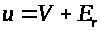 >,
или же >
>,
или же >
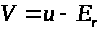 >.
Аналогично введем обозначения для
отсчитываемых от >
>
равновесных электрохимических потенциалов
>
>.
Аналогично введем обозначения для
отсчитываемых от >
>
равновесных электрохимических потенциалов
>
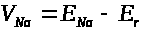 >
и >
>
и >
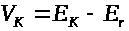 >.
Ясно, что сохраняются разности: >
>.
Ясно, что сохраняются разности: >
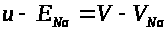 >
и >
>
и >
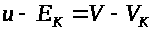 >.
В создании мембранного потенциала кроме
ионов >
>
и >
>
принимают участие и другие ионы, в
частности, ионы >
>.
В создании мембранного потенциала кроме
ионов >
>
и >
>
принимают участие и другие ионы, в
частности, ионы >
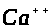 >.
Обозначим их суммарный электрохимический
потенциал через >
>.
Обозначим их суммарный электрохимический
потенциал через >
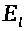 >,
соответственно >
>,
соответственно >
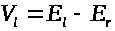 >.
В уравнение (3) для баланса мембранных
токов следует добавить слагаемое >
>.
В уравнение (3) для баланса мембранных
токов следует добавить слагаемое >
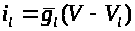 >.
Данный ток называется током утечки. Для
гигантского аксона кальмара А. Ходжкиным
и А. Хаксли были экспериментально найдены
значения: >
>.
Данный ток называется током утечки. Для
гигантского аксона кальмара А. Ходжкиным
и А. Хаксли были экспериментально найдены
значения: >
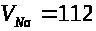 >
мв, >
>
мв, >
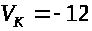 >
мв, >
>
мв, >
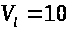 >мв.
>мв.
Электрохимическое
уравнение (3) суть обыкновенное
дифференциальное уравнение первого
порядка. Как показано выше, его коэффициенты
следует считать функциями от мембранного
потенциала >
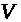 >.
По биологическому смыслу решения
уравнения ограничены на положительной
полуоси >
>.
По биологическому смыслу решения
уравнения ограничены на положительной
полуоси >
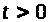 >.
Можно доказать, что все решения такого
уравнения монотонно стремятся к
состояниям равновесия. Объяснить в
рамках одного электрохимического
уравнения генерацию потенциала действия
невозможно.
>.
Можно доказать, что все решения такого
уравнения монотонно стремятся к
состояниям равновесия. Объяснить в
рамках одного электрохимического
уравнения генерацию потенциала действия
невозможно.
Из приведенных выше
построений следует, проводимости >
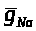 >
и >
>
следует считать функциями не только
мембранного потенциала >
>,
но и времени. Нижеследующие построения
А. Ходжкина и А. Хаксли во многом носят
феноменологический характер, т.е. не
имеют под собой прочной теоретической
основы и опираются на ряд допущений и
догадок. Рассмотрим натриевую проводимость.
Для описания ее зависимости от времени
и мембранного потенциала А. Ходжкин и
А. Хаксли ввели две новые переменные >
>
и >
>
следует считать функциями не только
мембранного потенциала >
>,
но и времени. Нижеследующие построения
А. Ходжкина и А. Хаксли во многом носят
феноменологический характер, т.е. не
имеют под собой прочной теоретической
основы и опираются на ряд допущений и
догадок. Рассмотрим натриевую проводимость.
Для описания ее зависимости от времени
и мембранного потенциала А. Ходжкин и
А. Хаксли ввели две новые переменные >
 >
и >
>
и >
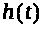 >,
которые характеризуют процессы активации
и инактивации натриевых каналов. В
интерпретации авторов >
>
-вероятность открытия >
>
-ворот, >
>
-вероятность закрытия >
>
-ворот, или же -отношение числа открытых
и закрытых ворот соответствующего типа
к числу каналов на поверхности мембраны
единичной площади. Авторы модели
положили: >
>,
которые характеризуют процессы активации
и инактивации натриевых каналов. В
интерпретации авторов >
>
-вероятность открытия >
>
-ворот, >
>
-вероятность закрытия >
>
-ворот, или же -отношение числа открытых
и закрытых ворот соответствующего типа
к числу каналов на поверхности мембраны
единичной площади. Авторы модели
положили: >
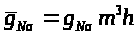 >,
где коэффициент >
>,
где коэффициент >
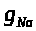 >
был назван максимальной натриевой
проводимостью. Предложенное А. Ходжкиным
и А. Хаксли обоснование вида зависимости
в настоящее время выглядит искусственным.
Методом подбора были выписаны линейные
обыкновенные уравнения первого порядка
для переменных >
>
и >
>.
При этом в уравнение для >
>
не входит переменная >
>,
а в уравнение для >
>
не входит >
>.
Коэффициенты уравнений зависят только
от мембранного потенциала >
>.
Более менее убедительное обоснование
необходимости именно такой зависимости
отсутствует.
>
был назван максимальной натриевой
проводимостью. Предложенное А. Ходжкиным
и А. Хаксли обоснование вида зависимости
в настоящее время выглядит искусственным.
Методом подбора были выписаны линейные
обыкновенные уравнения первого порядка
для переменных >
>
и >
>.
При этом в уравнение для >
>
не входит переменная >
>,
а в уравнение для >
>
не входит >
>.
Коэффициенты уравнений зависят только
от мембранного потенциала >
>.
Более менее убедительное обоснование
необходимости именно такой зависимости
отсутствует.
Для описания изменения
калиевой проводимости >
>
А. Ходжкиным и А. Хаксли была введена
функция >
 >.
Она интерпретировалась как вероятность
открытия >
>
-ворот для калиевых каналов, т.е. как
вероятность активации последних.
Напомним, что по гипотезе А. Ходжкина и
А. Хаксли инактивационные ворота для
калиевых каналов отсутствуют. Авторы
опытным путем подобрали линейное
обыкновенное дифференциальное уравнение
для >
>
и положили >
>.
Она интерпретировалась как вероятность
открытия >
>
-ворот для калиевых каналов, т.е. как
вероятность активации последних.
Напомним, что по гипотезе А. Ходжкина и
А. Хаксли инактивационные ворота для
калиевых каналов отсутствуют. Авторы
опытным путем подобрали линейное
обыкновенное дифференциальное уравнение
для >
>
и положили >
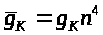 >.
Коэффициенты уравнения зависят только
от мембранного потенциала >
>.
Коэффициент >
>.
Коэффициенты уравнения зависят только
от мембранного потенциала >
>.
Коэффициент >
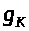 >
называется максимальной калиевой
проводимостью.
>
называется максимальной калиевой
проводимостью.
Относительно проводимости
>
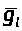 >
для тока утечки А. Ходжкин и А. Хаксли
предложили считать его постоянным: >
>
для тока утечки А. Ходжкин и А. Хаксли
предложили считать его постоянным: >
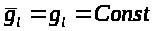 >.
В результате для описания процесса
генерации потенциала действия была
предложена система четырех уравнений,
Первое из них -электрохимическое
уравнение для баланса мембранных токов
: >
>.
В результате для описания процесса
генерации потенциала действия была
предложена система четырех уравнений,
Первое из них -электрохимическое
уравнение для баланса мембранных токов
: >
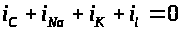 >.
Напомним, что
>.
Напомним, что
>
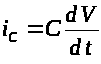 >,
>
>,
>
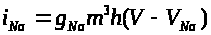 >,
(4)
>,
(4)
>
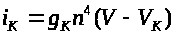 >,
>
>,
>
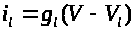 >.
(5)
>.
(5)
Второе и третье уравнения
описывают соответственно процессы
открытия >
>
и закрытия >
>
-ворот для натриевых каналов. Наконец,
четвертое уравнение отражает процесс
открытия >
>
-ворот для калиевых каналов. Система
уравнений Ходжкина -Хаксли имеет
следующий вид:
>
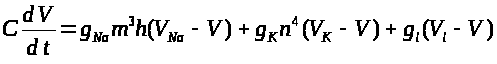 >,
(6)
>,
(6)
>
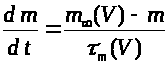 >,
(7)
>,
(7)
>
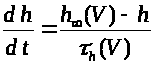 >,
(8)
>,
(8)
>
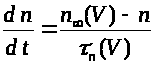 >,
(9)
>,
(9)
где, напомним, >
>,
>
>,
>
>.
В (7) -(9) функции >
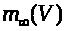 >,
>
>,
>
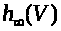 >,
>
>,
>
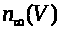 >,
>
>,
>
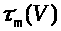 >,
>
>,
>
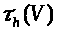 >,
>
>,
>
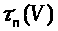 >
положительны и непрерывны. Функции >
>,
>
>,
>
>
относятся к классу так называемых
сигмоидных функций, т.е. они монотонны
обладают следующими свойствами: >
>
положительны и непрерывны. Функции >
>,
>
>,
>
>
относятся к классу так называемых
сигмоидных функций, т.е. они монотонны
обладают следующими свойствами: >
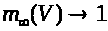 >,
>
>,
>
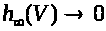 >,
>
>,
>
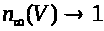 >
при >
>
при >
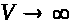 >
и >
>
и >
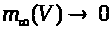 >,
>
>,
>
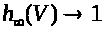 >,
>
>,
>
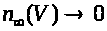 >
при >
>
при >
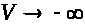 >.
В работах А. Ходжкина и А. Хаксли указан
их конкретный вид. Функции весьма сложно
устроены. Реально система уравнений
допускает только численное исследование.
Ограничимся рядом замечаний на
качественном уровне, иллюстрирующих
впечатления от численного исследования.
Входящие в систему (6) -(9) функции подобраны
так, что выполнен ряд свойств. Она имеет
устойчивое состояние равновесия >
>.
В работах А. Ходжкина и А. Хаксли указан
их конкретный вид. Функции весьма сложно
устроены. Реально система уравнений
допускает только численное исследование.
Ограничимся рядом замечаний на
качественном уровне, иллюстрирующих
впечатления от численного исследования.
Входящие в систему (6) -(9) функции подобраны
так, что выполнен ряд свойств. Она имеет
устойчивое состояние равновесия >
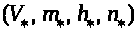 >,
где >
>,
где >
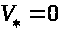 >,
>
>,
>
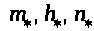 >
-положительны. Пусть >
>
-положительны. Пусть >
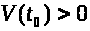 >,
>
>,
>
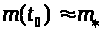 >,
>
>,
>
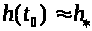 >,
>
>,
>
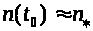 >.
Если значение >
>.
Если значение >
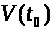 >
не слишком велико, то в силу устойчивости
состояния равновесия с течением времени
>
>
не слишком велико, то в силу устойчивости
состояния равновесия с течением времени
>
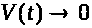 >.
Существует такое пороговое значение
>
>.
Существует такое пороговое значение
>
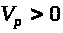 >,
что при >
>,
что при >
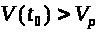 >
решения системы ведут себя по другому.
Первоначально >
>
решения системы ведут себя по другому.
Первоначально >
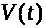 >
убывает. Однако начинает расти величина
>
>
убывает. Однако начинает расти величина
>
 >.
Слагаемое >
>.
Слагаемое >
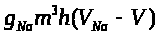 >
уравнении (6) положительно, т.к. >
>
уравнении (6) положительно, т.к. >
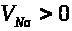 >,
а >
>
близко к нулю. В результате производная
V¢(t)
становится положительной. Мембранный
потенциал начинает расти (момент начала
генерации спайка). Вместе с ним растет
и величина >
>,
что приводит к дальнейшему увеличению
V¢(t)
и ускорению роста >
>.
Однако, чуть позже начинает уменьшаться
величина >
>,
а >
>
близко к нулю. В результате производная
V¢(t)
становится положительной. Мембранный
потенциал начинает расти (момент начала
генерации спайка). Вместе с ним растет
и величина >
>,
что приводит к дальнейшему увеличению
V¢(t)
и ускорению роста >
>.
Однако, чуть позже начинает уменьшаться
величина >
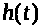 >.
Это замедляет рост >
>,
тем более, что уменьшается разность >
>.
Это замедляет рост >
>,
тем более, что уменьшается разность >
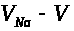 >.
Одновременно растет величина >
>.
Одновременно растет величина >
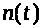 >.
Соответствующее слагаемое >
>.
Соответствующее слагаемое >
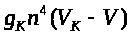 >
в уравнении (6) отрицательно, т.к. >
>
в уравнении (6) отрицательно, т.к. >
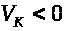 >
и >
>
и >
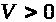 >.
Существует момент времени >
>.
Существует момент времени >
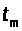 >,
где V¢(t>m>)=0.
Это точка максимума потенциала действия.
Значение мембранного потенциала >
>,
где V¢(t>m>)=0.
Это точка максимума потенциала действия.
Значение мембранного потенциала >
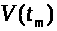 >
относительно близко к >
>
относительно близко к >
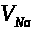 >.
После прохождения точки максимума
значение производной V¢(t)
определяется значением слагаемого >
>
и становится отрицательным. Скорость
убывания мембранного потенциала на
нисходящем участке спайка меньше по
абсолютной величине, чем скорость роста
на восходящем участке. Нисходящий
участок продолжительнее восходящего.
В дальнейшем значение >
>
становится отрицательным, а после этого
>
>.
Величины >
>.
После прохождения точки максимума
значение производной V¢(t)
определяется значением слагаемого >
>
и становится отрицательным. Скорость
убывания мембранного потенциала на
нисходящем участке спайка меньше по
абсолютной величине, чем скорость роста
на восходящем участке. Нисходящий
участок продолжительнее восходящего.
В дальнейшем значение >
>
становится отрицательным, а после этого
>
>.
Величины >
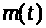 >,
>
>,
>
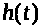 >,
>
>
также стремятся к своим равновесным
значениям. Процесс генерации спайка
закончен.
>,
>
>
также стремятся к своим равновесным
значениям. Процесс генерации спайка
закончен.
Особо подчеркнем значимость
факта, что система (6) -(7) обладает
пороговыми свойствами. Существует такое
критическое значение >
>
мембранного потенциала, что при начальном
отклонении мембранного потенциала >
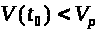 >
с течением времени значение >
>.
Если же >
>,
то через некоторое время (его называют
скрытым, или латентным периодом)
генерируется спайк. Модель адекватно
отражает пороговые свойства реального
аксона. Если воздействовать на мембрану
кратковременным толчком деполяризующего
тока (отклоняющего мембранный потенциал
в положительном направлении), то после
слабого воздействия мембранный потенциал
возвращается в равновесное состояние.
Вследствие достаточно сильного
воздействия спустя латентный период
генерируется спайк.
>
с течением времени значение >
>.
Если же >
>,
то через некоторое время (его называют
скрытым, или латентным периодом)
генерируется спайк. Модель адекватно
отражает пороговые свойства реального
аксона. Если воздействовать на мембрану
кратковременным толчком деполяризующего
тока (отклоняющего мембранный потенциал
в положительном направлении), то после
слабого воздействия мембранный потенциал
возвращается в равновесное состояние.
Вследствие достаточно сильного
воздействия спустя латентный период
генерируется спайк.
Обсудим еще одно свойство аксона. Гигантские аксоны достигают длины нескольких десятков сантиметров. Мембранный потенциал на разных участках аксона может различаться. Аксон представляет собой так называемую распределенную систему. Если подействовать на некоторый участок аксона допороговым деполяризующим толчком тока, то мембранный потенциал на этом и некотором примыкающем к нему участке отклонится от равновесного состояния в положительном направлении. Такое отклонение называется локальным ответом. Возмущение с течением времени затухает, одновременно, как капля, растекаясь по аксону. В результате локальные ответы, возникшие при стимулировании различных участков, могут накладываться друг на друга, т.е., как говорят, суммироваться по пространству. Если же на участок аксона повторно подается деполяризирующий ток, то новый локальный ответ накладывается на предыдущий. Говорят, что происходит временное суммирование. Локальные ответы градуальны. Их амплитуда зависит от силы стимула (точнее, силы -длительности). В случае, когда локальный ответ в результате суммации, или же после достаточно сильного одноразового локального воздействия достигает порогового уровня, запускаются механизмы генерации потенциала действия. Зародившись локально, потенциал действия распространяется по аксону.
Явление распространения объясняется следующим образом. Цитоплазма - электролит, а, следовательно, хороший проводник. Если между точками существуют ненулевые разности потенциалов, то вдоль мембраны возникают токи (вблизи поверхности). Они направлены от точек с более высоким потенциалом к точкам, где он меньше, и подчиняются закону Ома. Вследствие этого потенциал на участках, прилегающих, к области генерации спайка, растет и достигает порогового значения. В результате эти участки сами генерируют потенциалы действия. Процесс распространяется вдоль аксона.
Проведение нервных импульсов обладает важной особенностью. При “столкновении” они аннигилируют, т.е. уничтожаются. С помощью локального воздействия возбудим аксон в двух точках. По нему будут распространяться четыре импульса, два из которых движутся во встречных направлениях. Данные импульсы встретятся и не пропустят друг друга. Это связано с тем, что мембрана обладает следующим свойством. Во время потенциала действия и некоторое время после его завершения она не реагирует на воздействие. В частности, после завершения спайка некоторое время не способна сгенерировать нового потенциала действия. Данное состояние называется абсолютной рефрактерностью. Отметим, что постепенно абсолютная рефрактерность сменяется относительной. Для того, чтобы инициировать потенциал действия в состоянии относительной рефрактерности, нужно применить более мощный стимул, нежели в состоянии покоя.
Для описания процесса
распространения потенциала действия
вдоль аксона А. Ходжкин и А. Хаксли
модернизировали уравнение (6). Пусть >
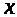 >
-координата точки вдоль аксона, а >
>
-координата точки вдоль аксона, а >
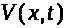 >
-значение мембранного потенциала в этой
точке. Используется закон Ома, согласно
которому ток через сечение пропорционален
градиенту напряжения (в одномерном
случае -производной). Ток через сечение
>
>
равен >
>
-значение мембранного потенциала в этой
точке. Используется закон Ома, согласно
которому ток через сечение пропорционален
градиенту напряжения (в одномерном
случае -производной). Ток через сечение
>
>
равен >
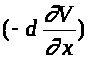 >,
где направление выбрано в сторону
возрастания >
>,
коэффициент >
>,
где направление выбрано в сторону
возрастания >
>,
коэффициент >
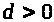 >
-проводимость. Выделим участок аксона,
координаты точек которого заключены в
интервале >
>
-проводимость. Выделим участок аксона,
координаты точек которого заключены в
интервале >
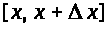 >.
Суммарный ток через сечения >
>
и >
>.
Суммарный ток через сечения >
>
и >
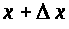 >
равен >
>
равен >
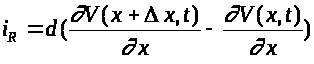 >.
Используя формулу Лагранжа получим: >
>.
Используя формулу Лагранжа получим: >
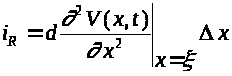 >,
где >
>,
где >
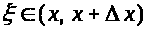 >.
Через поверхность мембраны выбранного
участка аксона проходят емкостной >
>.
Через поверхность мембраны выбранного
участка аксона проходят емкостной >
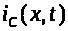 >,
натриевый >
>,
натриевый >
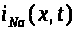 >,
калиевый >
>,
калиевый >
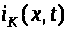 >
токи, а также ток утечки >
>
токи, а также ток утечки >
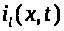 >.
Пусть >
>.
Пусть >
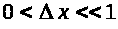 >.
Суммарный мембранный ток приближенно
равен >
>.
Суммарный мембранный ток приближенно
равен >
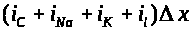 >.
Привлекая закон Кирхгофа, получим: >
>.
Привлекая закон Кирхгофа, получим: >
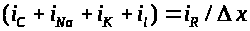 >.
Используя приведенные ранее формулы
(4), (5) для мембранных токов и устремляя
>
>.
Используя приведенные ранее формулы
(4), (5) для мембранных токов и устремляя
>
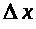 >
к нулю, приходим к уравнению:
>
к нулю, приходим к уравнению:
>
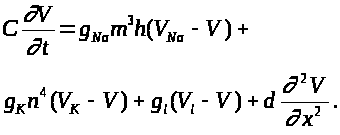 >
(10)
>
(10)
Его часто называют
уравнением аксона Ходжкина -Хаксли. В
(10) функции >
>,
>
>,
>
>
как и выше подчиняются уравнениям (7)
-(9).
А. Ходжкин и А. Хаксли
исследовали так называемые автоволновые
решения системы уравнений, т. е. решения
типа бегущих волн: >
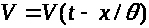 >
(аналогично для >
>,
>
>,
>
>).
Здесь параметр >
>
(аналогично для >
>,
>
>,
>
>).
Здесь параметр >
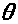 >
-скорость распространения волны. После
подстановки в (11) для >
>
получается обыкновенное дифференциальное
уравнение:
>
-скорость распространения волны. После
подстановки в (11) для >
>
получается обыкновенное дифференциальное
уравнение:
>
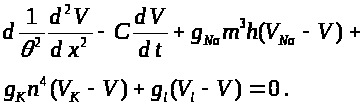 >
(11)
>
(11)
Система (7) -(9), (11)
исследовалась численно. Методом подбора
было найдено значение скорости >
>,
при которой система уравнений имела
приемлемое по мнению авторов решение.
Параметры полученного при исследовании
системы уравнений распространяющегося
потенциала действия оказались близкими
к реально наблюдаемым в эксперименте.
Скорость распространения импульса по гигантскому аксону относительно не велика (по сравнению с аналогичными по назначению проводниками нервных импульсов у человека). Она растет с увеличением диаметра аксона. Увеличение диаметров аксонов избрала природа для высокоскоростного проведения импульсов у беспозвоночных. Однако, когда организму нужна не только быстрота реакции, но и огромное количество связей, гигантские аксона становятся не приемлемыми. Например, в зрительном нерве, где больше миллиона связей, для больших аксонов не хватило бы места. Длинные аксоны у человека и позвоночных животных покрыты достаточно толстым липидным слоем -миелиновой оболочкой (точнее, они обвиты клетками содержащими и вырабатывающими миелин). В миелиновой оболочке присутствуют регулярно расположенные разрывы, где мембрана аксона оголена. Разрывы называются перехватами Ранвье. Миелинизированные участки имеют длину порядка 1 -2 мм. Протяженность перехватов Ранвье около 1 мкм. Перехваты Ранвье способны генерировать спайки. Миелинизированные участки обладают емкостью и омическим сопротивлением, но они не генерируют спайков. Импульсы по миелинизированным участкам распространяются пассивно, т.е. с затратами энергии на омическое сопротивление и на перезаряд участка, представляющего собой конденсатор (обратимые потери). Суммарно затраты энергии не велики.
Рассмотрим миелинизированный участок, ограниченный двумя перехватами. Пусть левый перехват генерирует спайк. Возмущение (положительное) по миелинизированному участку передается правому перехвату. Когда отклонение мембранного потенциала правого участка достигает порогового значения, он генерирует спайк. Этот импульс не может вызвать генерацию нового спайка левым перехватом Ранвье, поскольку тот находится в рефрактерном состоянии (в состоянии невосприимчивости). Однако, данный перехват по той же схеме возбуждает перехват, следующий за ним справа. По миелинизированному аксону, перескакивая от перехвата к перехвату, будет распространяться импульс. Процесс называется сальтаторным проведением возбуждения (saltare - прыгать). Оно обеспечивает скорость распространения импульсов в 20- 25 раз более высокую, чем в гигантских аксонах того же диаметра.
Описанный процесс
сальтаторного проведения импульсов
легко формализуется. В простейшем случае
это выглядит следующим образом. Пусть
>
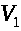 >
и >
>
и >
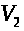 >
- мембранные потенциалы левого и правого
перехватов Ранвье, а >
>
-потенциал миелинизированного участка
(считаем его во всех точках участка
одинаковым). Тогда между участком и
соответствующими перехватами проходят
токи: >
>
- мембранные потенциалы левого и правого
перехватов Ранвье, а >
>
-потенциал миелинизированного участка
(считаем его во всех точках участка
одинаковым). Тогда между участком и
соответствующими перехватами проходят
токи: >
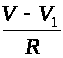 >
и >
>
и >
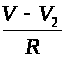 >,
где >
>
-сопротивление перехода от перехвата
к миелинизированному участку. Через
миелиновую оболочку проходят емкостной
ток>
>,
где >
>
-сопротивление перехода от перехвата
к миелинизированному участку. Через
миелиновую оболочку проходят емкостной
ток>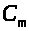 >V¢
и ток утечки >
>V¢
и ток утечки >
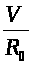 >,
где >
>
- суммарная емкость, а >
>,
где >
>
- суммарная емкость, а >
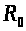 >
суммарное сопротивление миелиновой
оболочки. В силу первого закона Кирхгофа
получаем:
>
суммарное сопротивление миелиновой
оболочки. В силу первого закона Кирхгофа
получаем:
>
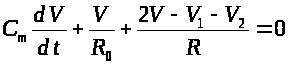 >.
>.
Для описания мембранных
потенциалов >
>
и >
>
перехватов служит уравнение (6), в правую
часть которого согласно закону Кирхгофа
следует добавить для левого перехвата
слагаемое >
>,
а для правого -слагаемое >
>.
Это токи, проходящие между соответствующими
перехватами и миелинизированным
участком. В результате получим уравнения:
>
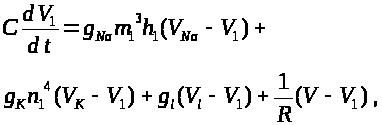 >
>
>
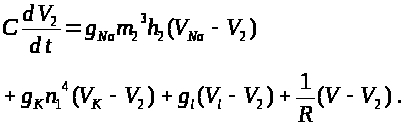 >
>
Величины >
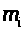 >,
>
>,
>
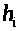 >
и >
>
и >
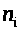 >
>
>
>
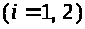 >
удовлетворяют уравнениям (7) -(9), в которых
для левого и правого перехватов следует
положить >
>
удовлетворяют уравнениям (7) -(9), в которых
для левого и правого перехватов следует
положить >
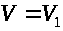 >
и >
>
и >
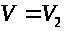 >
соответственно.
>
соответственно.
Ключевые источники естествознания
Имея в виду рациональность перехода от натурфилософии к математически точному естествознанию, В.И. Вернадский отмечал одинаковую существенность и взаимную дополнительность двух основных и воистину универсальных математических методов - количественного (арифметического или алгебраического) и качественного (геометрического), т. е. интегрального (внешнего) и дифференциального (внутреннего): “Одно и то же природное явление может быть независимо охвачено обоими этими направлениями творческой математической мысли”.
Отдавая должное философии и сознавая “огромное значение математики для естествознания”, он все-таки полагал, что “в основе естествознания лежат только научные эмпирические факты и научные эмпирические обобщения”:
“Все основные научные эмпирические понятия при логическом анализе приводят к иррациональному остатку...Никогда ни одно научно изучаемое явление, ни один научный эмпирический факт и ни одно научное эмпирическое обобщение не может быть выражено до конца, без остатка, в словесных образах, в логических построениях - в понятиях - в тех формах, в пределах которых только и идет работа философской мысли, их синтезирующая, их анализирующая. В предметах исследования науки всегда остается неразлагаемый рационалистически остаток, иногда большой, который влияет на эмпирическое научное изучение, остаток, исчезающий нацело из идеальных построений философии, космогонии или математики и математической физики. Глубокая мысль, в яркой красивой форме выраженная Ф. И. Тютчевым- “Мысль изреченная есть ложь”, всегда сознательно или бессознательно чувствуется испытателем природы и всяким научным исследователем, когда он в своей научной работе сталкивается с противоречиями между эмпирическими научными обобщениями и отвлеченными построениями философии или когда факты заставляют его менять и уточнять (обычно осложнять, а часто резко упрощать) свои гипотезы, особенно часто - неизбежно ограниченные математические выражения природных явлений”.
Поэтому Вернадский считал необходимым исходить прежде всего или, в конечном счете, именно из ключевых научных эмпирических фактов или соответствующих ключевых научных эмпирических обобщений (типа открытой Менделеевым Периодической системы химических элементов—“одного из величайших эмпирических обобщений”), т. е. обращаться непосредственно к этим естественным ключевым источникам, как бы вспоминая красноречивое тютчевское “Молчание”:
Как сердцу высказать себя?
Другому как понять тебя?
Поймет ли он, чем ты живешь?
Мысль изреченная есть ложь.
Взрывая, возмутишь ключи,
Питайся ими—и молчи.
Всегда остающийся в предметах исследования науки неразлагаемый рационалистически остаток, т. е. иррациональный остаток, к которому приводят все основные научные эмпирические понятия при логическом анализе, означает, что мы должны принимать во внимание наряду с безусловно необходимыми - достоверными - фактами, характерными для вполне детерминистической классической механики, и факты вероятностные, лежащие в основе надлежащей квантовой механики (с ее соответствующей вероятностной интерпретацией и с характерным для нее принципом неопределенности), а также факты веры, с которыми имеют дело не только все религии, но и атеизм, поскольку “основанные на философских заключениях” “атеистические представления,—как справедливо заметил Вернадский, - по существу тоже предмет веры”.
Чуть ли не все подвергая сомнению, Вернадский замечает: “Но это не касается эмпирических обобщений, которые в основе своей существенно отличны от научных теорий и научных гипотез, с которыми они обычно смешиваются”.
При этом он считал принципиально необходимым и возможным стремиться к предельно полному охвату природных явлений и самой Природы в целом.
Однако в пределе, охватывая в целом Природу, Вселенную, материю (со всеми присущими ей атрибутами, вплоть до Жизни и Разума, в том числе Высшего Разума - с бесконечными потенциальными возможностями), мы по крайней мере в принципе можем и должны получить - и действительно получаем! - не только искомое воистину универсальное (предельно полное) ключевое научное эмпирическое обобщение в виде вполне детерминированных взаимосвязанных периодических систем всевозможных (так называемых эталонных и производных) фундаментальных структурных элементов материи на всех четырех возможных последовательных основных уровнях ее естественной самоорганизации - физическом, химическом, биологическом и психологическом (т. е. на самом деле величайшее атомистическое научное эмпирическое обобщение менделеевского типа), но и адекватное ему столь же универсальное ключевое научное теоретическое обобщение в виде совершенно однотипных по своей симметрии и, соответственно, непосредственно однозначно дедуктивно определяемых по надлежащей математической индукции вполне детерминированных взаимосвязанных периодических систем всевозможных равномерно квантованных собственных значений всех возможных универсальных характеристик рассматриваемых элементов.
Список используемой литературы
1. Пуанкаре А. “О науке”.М., 1983
2. Аристотель “Сочинения”. М., “Мысль”, 1975 г.
3. Кант И. “Сочинения”, М., “Мысль”, 1964 г.
Н.К.Вахтомин “Теория научного знания Иммануила Канта”. М., “Наука”, 1986 г.
Кедров Б.М. “Предмет и взаимосвязь естественных наук”.М., “Наука”,1967.436с.
“Природа и общество” М, “Наука,1968.348с.
“Физика ХХ века. Развитие и перспективы.” Сборник. М,“Наука,1984.334с.
8.Алексеев И.С., Овчинников Н.Ф., Печенкин А.А. “Методология обоснования квантовой теории”. М.: “Наука”,1984.334с.
9.В.И.Кузнецов, Г.М.Идлис, В.Н.Гутина “Естествознание” М, “Агар”,1996,384с.