Розвиток кісткової тканини в онтогенезі безхвостих земноводних
План
Вступ 2
Розділ 1. Загальна характеристика кісткової тканини 4
Розділ 2. Особливості будови окістя в безхвостих амфібій 10
Розділ 3. Проліферативні властивості клітин окістя в амфібій 17
Висновок 24
Література 25
Вступ
За будовою та функціональним значенням кісткова тканина є унікальним різновидом сполучної тканини. Вона містить велику кількість мінеральних солей, серед яких найбільше сполук кальцію у вигляді гідроксиапатитів (Са>10>(РО>4>)>6>)(ОН)>2 >та фосфатів (Са>3>(РО>4>)>2>), що становить 50—70% сухої маси кістки.
Особливості хімічного складу, орієнтація колагенових волокон вздовж довгої осі кістки забезпечує її величезну механічну міцність та фізіологічну активність. Висока чутливість кісткової тканини до дії факторів зовнішнього середовища зумовлена інтенсивно перебігаючими в ній обмінними процесами. Кісткова тканина постійно знаходиться у стані оновлення і ремоделювання. Кістки скелета, черепа, грудної клітки забезпечують механічний захист органів центральної нервової системи, грудної та тазової порожнин. У губчастій речовині кісток скелету знаходиться червоний кістковий мозок, у якому відбуваються процеси кровотворення та диференціювання клітин імунного захисту організму. Частина мінеральних солей, що звільняються при перебудові скелету, надходить в кров, їх мобілізація інтенсивна в період вагітності та лактації, коли кальцій необхідний для розвитку зародка або новонародженого.
Земноводні тварини ведуть як водний так і наземний спосіб життя. Для їх організму властиве в зимовий період замерзання тіла, а навесні вони відновлюють всі наявні для них функції життєдіяльності. Ці тварини відносяться до холоднокровних, вони мають маленькі розміри порівняно з більшістю інших хребетних тварин. На безхвостих земноводних досить часто проводяться лабораторні досліди. Для кісткової тканини даних тварин характерна більш крихка структура, ніж у птахів та ссавців.
Метою нашої роботи є з`ясування особливостей процесу розвитку кісткової тканини у безхвостих земноводних.
Об`єктом дослідження є: безхвості земноводні.
Предмет дослідження – особливості будови окістя у безхвостих земноводних
Завдання роботи:
1 – сформувати загальні уявлення про кісткову тканину
2 – охарактеризувати особливості будови окістя у амфібій
3 – розглянути проліферативні властивості клітин окістя у амфібій.
В процесі вивчення та аналізу даної теми ми використовували наступні методи наукового дослідження: аналіз літературних даних, порівняння, узагальнення.
Розділ 1. Загальна характеристика кісткової тканини
Будова кісткової тканини. До складу кісткової тканини входять клітини та матрикс (міжклітинна речовина). Серед клітин значне місце належить остеобластам, їх основною функцією є синтез та секреція органічної частини матриксу кістки навколо своїх відростків, що сприяє утворенню канальців. Відростки остеобластів анастомозують між собою. Розмір остеобластів 12—20 мкм, за формою вони кубічні, призматичні, полігональні з базофільною цитоплазмою. Мають добре розвинену гранулярну ендоплазматичну сітку, вільні рибосоми, комплекс Гольджі, мітохондрії. Розміщуються рядами на поверхні новоутворюваної тканини, їх цитоплазма характеризується позитивною реакцією на лужну фосфатозу, ядро розміщується якнайдальше від кісткової речовини.
Остеоцити знаходяться у своєрідних порожнинах міжклітинної речовини — лакунах, з'єднаних між собою численними кістковими канальцями. Остеоцит являє собою більш диференційовану стадію розвитку остеобласта. За формою це переважно овальні клітини, розміром 18—40 мкм, містять добре помітне ядро та слабо базофільну цитоплазму, бідну на органели. Велика кількість відростків остеоцитів знаходиться у кісткових канальцях та порожнинах, де циркулює тканинна рідина, яка бере участь у метаболізмі кісткової тканини.
Остеокласти. На поверхні кісткової тканини в місцях знаходження остеобластів відбувається активна резорбція, яка є невід'ємним процесом її перебудови. Остеокласти — великі багатоядерні клітини (до 100 і більше), їх розмір 20— 100 мкм, цитоплазма оксифільна або базофільна, містить багато лізосом, мітохондрії, кілька структур комплексу Гольджі. Поверхня остеокластів, що межує з поверхнею кістки в місцях її резорбції, має гофровану облямівку, що розгалужується. Тут утворюються і нагромаджуються гідролітичні ферменти, які беруть участь у процесах резорбції кістки. Дія остеокластів гальмується під впливом гормона тіреокальцитоніну та посилюється дією паратгормона.
Матрикс кісткової тканини складається з осеїнових (колагенових) волокон та аморфної речовини — осеомукоїду. До її складу входять глікопротеїаи, сульфатовані глікозаміноглікани, білки, неорганічні сполуки — фосфат кальцію, гідроксиапатіт, різні мікроелементи (магній, мідь, цинк, барій та інш.) (рис. 1).
Залежно від структурної організації міжклітинної речовини, головним чи-ном осеїнових волокон, серед кісткової тканини розрізняють грубоволокнисту та пластинчасту.
Грубоволокниста кісткова тканина типова для кісток зародка, а у дорослих тварин вона є у ділянці швів черепа, місцях прикріплення сухожиль до кісток, зубних альвеол, кісткового лабіринту внутрішнього вуха. Вона характеризується значною товщиною пучків осеїнових волокон, різноманітністю їх орієнтації, оточена звапнованим осеомукоїдом; у лунках осеомукоїду знаходяться остеоцити. Пластинчаста кісткова тканина характеризується утворенням кісткових пластинок товщиною 4—16 мкм, у них осеїнові волокна розміщені паралельно. Суміжні пластинки завжди мають іншу орієнтацію осеїнових волокон. Кісткові пластинки утворюють цілі системи, які тісно пов'язані з кровоносними судинами та нервами. Залежно від розміщення кісткових пластинок розрізняють губчасту та компактну речовини. В губчастій речовині, особливо в епіфізах трубчастих кісток кісткові пластинки знаходяться під різними кутами відповідно напрямку дії механічного навантаження даної кістки. Завдяки цьому у трубчастій речовині утворюються комірки, в яких міститься кістковий мозок (рис. 2, 3).
Губчаста речовина інтенсивно забезпечується кров'ю. Вона характеризується активним мінеральним обміном. Компактна кісткова речовина складається з пластинок, що утворюють діафізи трубчастих кісток. Зовні діафіз оточений окістям (періостом), воно забезпечує ріст, живлення, регенерацію. Окістя складається з зовнішнього та внутрішнього шарів. Зовнішній шар утворюється щільною колагеновою сполучною тканиною, тісно пов'язаною з сухожиллям м'язів та колагеновими волокнами зв'язок. Від окістя у кістку проходять проривні (шарпеєві) пучки колагенових волокон, що забезпечують міцність зв'язку окістя з кісткою. Внутрішній шар окістя містить колагенові та еластичні волокна, остеоцити та остеобласти. В окісті знаходяться нерви та кровоносні судини.
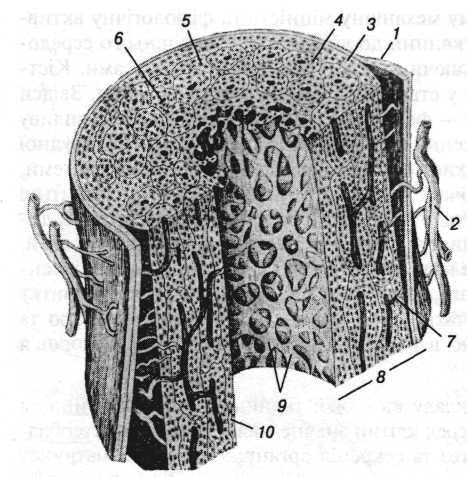
Рис. 1. Схема будови трубчастої кістки:
1 — окістя; 2 — кровоносні судини; 3 — зовнішня загальна система кісткових пластинок; 4 — остеон; 5 — вставна система; 6 — канал остеона; 7 — фолькманівський канал; 8 — компактна кістка; 9 — губчаста кістка; 10 — внутрішня загальна система кісткових пластинок
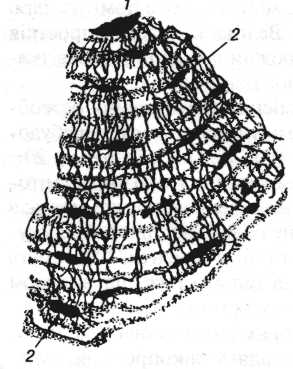
Рис. 2. Поперечний розріз трубчастої кістки:
1 — канал остеона; 2 — остеоцити
Останні через відповідні канали проникають у кісткову тканину та кістковий мозок. Кісткові пластинки компактної речовини діафізів трубчастих кісток утворюють зовнішню загальну систему пластинок, остеонний шар, вставні системи і внутрішню загальну систему кісткових пластинок. Кісткові пластинки зовнішньої загальної системи розміщені паралельно до поверхні кістки і тісно зв'язані з окістям. Остеонний шар займає більшу площу кістки.
Остеон — сукупність 4— 20 трубчастих пластинок, вставлених одна в другу; за своєю формою вони мають вигляд спиляного дерева. У копитних тварин остеони побудовані з пластинок з майже горизонтальним або вертикальним спрямуванням колагенових волокон, що забезпечує міцність та пружність. їх центральні канали містять судини, остеобласти та остеокласти. Між остеонами знаходяться вставні пластинки, що являють собою залишки остеонів у стані моделювання.
Внутрішні загальні пластинки межують з ендоостом, останній відмежовує кістковомозкову порожнину і складається з тонковолокнистої сполучної тканини і містить остеобласти та остеокласти. В пластинках зовнішньої загальної системи проходять проривні пучки і канали з кровоносними судинами та нервами, розгалужуються, з'єднуються з центральними каналами остеонів і анастомозують між собою. Остеоцити лежать між кістковими пластинками, а кісткові канальці пронизують системи пластинок.
Розвиток кісткової тканини — остеогенез. Існує два різновиди остеогенезу — безпосередньо із мезенхіми та заміни кісткою гіалінового хряща. Ділянки мезенхіми, в яких почався остеогенез за зовнішнім виглядом являють собою мембрани, осифікація у них називається інтрамембранною. Цей остеогенез характерний для розвитку кісток черепа та нижньої щелепи, на місці, де утворюється кістка, спочатку знаходиться шар рихлої мезенхіми, вона має вигляд зірчастих клітин з відростками, які вступають в контакт з відростками сусідніх клітин мезенхіми, утворюючи сітку, остання занурена в аморфну міжклітинну речовину, що містить окремі пучки колагенових волокон і кровоносні судини. Окремі мезенхімні клітини відмежовані міжклітинною речовиною. На поверхні таких остеогенних острівців, швидко і непомітно переходять через стадію остеогенних клітин, диференціюються в остеобласти, що беруть участь в остеогенезі. Окремі остеобласти втрачають здатність синтезувати міжклітинну речовину, замуровуються в неї і перетворюються в остеоцити, міжклітинна речовина насичується фосфатом кальцію, який нагромаджується в кістковій тканині у результаті розпаду гліцерофосфату крові під впливом лужної фосфатази. Фосфат кальцію та вуглекислий кальцій, що при цьому утворюються, насичують основну речовину кісткової тканини. В зоні мінералізації остеобластами інтенсивно секретуються глікозаміноглікани, які потім розщеплюються лізосомними ферментами, утворюючи при цьому високоактивні аніони. В цьому процесі значне місце належить АТФ, яка може бути одночасно і джерелом фосфору, необхідного для кальцифікації й джерелом енергії. Цей тип остеогенезу характерний для розвитку первинної грубоволокнистої кісткової тканини. В подальшому первинна грубоволокниста тканина заміщується пластинчастою кістковою тканиною.
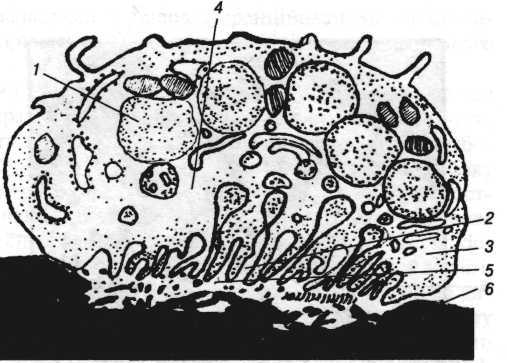
Рис. 3. Схема будови остеокласта:
1 — ядро; 2 — гофрований край остеокласта; 3 — світла зона; А — лізосоми; 5 — зона резорбції міжклітинної речовини; 6 — мінералізована міжклітинна речовина
Остеогенез на місці хряща. Інший різновид остеогенезу характерний для кісток тулуба, кінцівок, основи черепа. Він також відбувається в мезенхімі, однак йому передує формування моделі з гіалінового хряща, який руйнується і замінюється кістковою тканиною. При цьому в ділянці навколо хряща виникає волокниста кісткова манжетка, що сприяє появі перихондрального окостеніння. Воно відбувається за рахунок міграції із судин охрястя остеогенних клітин, які продукують осеїнові волокна й хондромукоїд, відбувається їх звапнування. Поступово манжетка потовщується і росте в напрямку епіфізів. Вона відмежовує ріст хряща в товщину і зумовлює формування у ньому клітинних колонок. Клітини хряща збільшуються в розмірі, набувають пузирчастого вигляду, їх ядра зморщуються, розпадаються. Міжклітинна речовина хряща між колонками клітин ущільнюється, заповнюється, утворює хрящові балки. Настає енхондральне окостеніння, яке супроводжується утворенням діафізарного центру окостеніння, що починається з проростання кровоносних судин кісткової манжетки всередину діафізу хрящової моделі; з судинами мігрують різнодиференційовані мехенхімні клітини гемопоетичного ряду й остеогенні. За рахунок остеокластів у хрящі виникають порожнини резорбції. Останні з'єднуються і утворюють кістково-мозкову порожнину. Малодиференційована сполучна тканина утворює первинний кіст-ковий мозок, збагачений остеобластами та малодиференційованими елементами, які в подальшому перетворюються у елементи дефінітивного кісткового мозку.
Навколо кровоносних судин утворюються кісткові пластинки, формуються остеони. В подальшому енхондральна кісткова тканина повністю розсмоктується і на її місці утворюється кістково-мозковий канал, зникає примітивна грубоволокниста кісткова тканина. Одночасно з енхондральним остеогенезом з боку окістя відбувається активний перихондральний остеогенез. Періостальна кісткова тканина являє собою компактну речовину кісток скелету, її будова типова пластинчастій з характерними системами кісткових пластинок. В епіфізах кісток центри окостеніння виникають пізніше. Окостеніння, яке виникає у діафізі, поширюється в напрямку епіфізів. У них з'являються місця окостеніння, за винятком хряща, що покриває суглобову поверхню та хрящову пластинку росту (епіфізарну лінію), яка відмежовує епіфіз від діафізу. Хрящ суглобової поверхні зберігається протягом усього життя тварини; у дорослих тварин хрящ епіфізарної лінії костеніє. За рахунок хряща епіфізарної лінії кістка росте в довжину.
Регенерує кісткова тканина за рахунок камбіальних елементів окістя. Перебудова кісток відбувається протягом усього життя тварини.
Розділ 2. Особливості будови окістя в безхвостих амфібій
На стегнових кістах амфібій (личинки Rana temporarіa, Pelobates fuskus, Hyla arborea на стадії 46, 47, 49 по Терентьєву, 1950; на стадії метаморфоза личинки, сеголіток і статевозрілих амфібій) у різні періоди онтогенезу вивчалися початкові стадії появи періосту і його наступний розвиток у процесі заміщення хрящової закладки кістою зі звертанням особливої уваги на клітинний склад і загальну архітектоніку за морфологічними і гістохімічними ознаками.
Шматочки стегнової кісти разом з періостом або тільки відщиплений періост вирізували із середньої частини діафізу, зони проксимального і дистального метафізів.
Відібраний матеріал фіксували в рідині Карнуа, 10 %-м нейтральному формаліні, 6 %-м формаліні, на насиченому розчині бензойної кислоти, у 80° і 96°-м етанолі при кімнатній температурі і при +4°С. У тих випадках, коли періост вирізували з підлягаючою кістою, відібрані шматочки поміщали в 10 %-й розчин динатриєвої солі етилендіаментетрауксусной кислоти при 18 °С (рН 7,4). Підготовлені зразки заливали в парафін. Гістологічні зрізи (5-6 мкм) офарблювали гематоксилин-еозином по Майеру і по Делафильду, а також селективними методами для виявлення визначених структур; у клітках визначали мітотичний індекс площі кліток, їхній ядерно-плазменне відношення. Числові показники оброблені методами варіаційної статистики.
У личинок на стадії 46 періост починає диференціюватися в діафізарній зоні майбутньої кісти. Перші ознаки периостальних структур з'являються на основі перихондра ще до виникнення тут кісткової пластинки. У цей час у найглибшому шарі перихондра клітини набувають характерного вигляду остеобластів і починають продукувати фермент лужну фосфатазу. Ці ознаки свідчать про початок розгортання периостального остеопластичного процесу. Очевидно, поява перших остеобластів зв'язано з проникненням кровоносних судин у перихондр і остеогенного диференціювання клітин перихондра в локусах, підготовлених до периостальному остеогенезі. Діяльністю цих клітин по окружності діафізарної хрящової закладки у вигляді такого пояса починає формуватися периостальна кісткова манжетка.
Клітини, що мають властивості остеобластів, морфологічно виділяються в новому періості більш великими розмірами, округлим ядром із сітчастою структурою і масивною цитоплазмою зі слабкими оксифільними властивостями. Для них характерна також визначена топографія: вони розташовуються безпосередньо в поверхні закладки і повернені до неї цитоплазматичной масою, тоді як ядро в кожній клітині знаходиться на протилежному кінці. Такий полярний розподіл ядра і цитоплазми додає зрілому функціонуючому остеобластові своєрідний вид комети, поверненої "хвостом" до споруджуваної кісти (мал. 4). Остеобласти, що дозрівають, (преостеобласти) мають більш симетричні контури з центральною локалізацією ядра. Маси цитоплазми в них не перевищує 1/3 її обсягу в зрілих формах. По фронті остеогенеза остеобласти в періості розташовуються в один ряд. Тут майже немає волокнистих структур, фіброзний каркас розташовується трохи вище і зв'язаний безпосередньо з преостеобластичним диференціюванням. Загальна товщина періосту в діафізарній зоні не перевищує 52 мкм.
У метафізарній зоні трубчастої кісти вже на 46-й стадії остеогенезу жаби мається типове для земноводних грибоподібне розростання епіфізарного хряща (мал. 5). У круговій щілині між диафизарной областю кісти, що розвивається, і хрящовим дивертикулом її епіфіза з моменту появи діафізарної кісткової манжетки сусідять на невеликій відстані кіста і хрящ. У цій області зберігається також своєрідна структура, що свідчить не тільки про можливу функціональну сумісність перихондра і періосту, але і про їхню єдність по генезу.
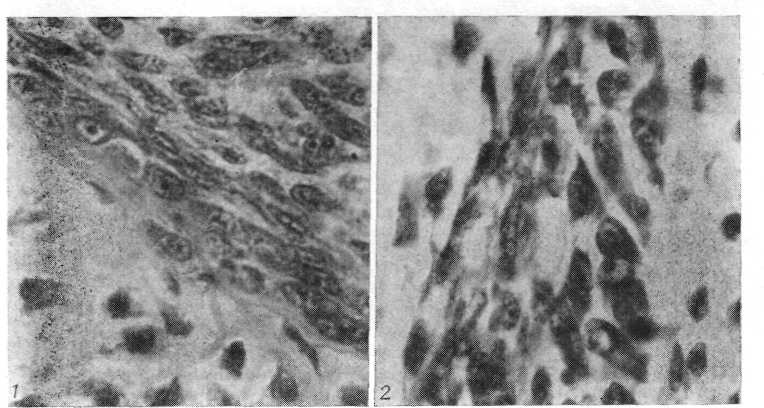
Рис. 4. Перихондр закладки стегнової кістки Rana temporarіa (в області диафиза). Видні остеобласти, що диференціюються, у внутрішньому шарі перихондра (гематоксилін Делафильда - эозин; про. 40, ок. 7).
Рис. 5. Періост області завороту хрящового епіфіза і метафиза стегнової кістки жаби. Між хрящовим епіфізом і кісткою розташовуються скелетогенные клітини (гематоксилін Делафильда - еозин; про. 60; ок. 7).
Перихондрально-періостальна ділянка включає всього 2-3 ряду клітин, з яких внутрішній шар складається з остеобластів, а зовнішній - з хондробластів (мал. 5). У кінцевому верхівковому відділі щілини, куди край кісткової трубки не досягає, утримуються тільки хондробласти.
У личинок 47-й стадії розвитку в трубчастих кістах кінцівок зберігається та ж ситуація, що описана для стадії 46 з тією лише різницею, що перихондральна кісткова трубка в області діафізу має трохи більші розміри по довжині і масі. Якщо на попередній стадії зачаток кісткового діафізу мав вигляд на гістологічному зрізі як тонка (10-12 мкм) пластинка, то на стадії 47 вона досягає в товщину 24 мкм і вже містить у своєму складі одиничні остеоцити (включені в кісту остеобласти) (мал. 6). У центральній частині діафізу остеобласти періосту здобувають подовжену форму і подовжнє розташування без вираженого полярного орієнтування до фронту остеогенеза (мал. 6). Зміна форми в остеобластах, як правило, сполучено з їхньою функціональною активністю (Мажуга, 1978; Вечерська, 1984).
Можна думати тому, що в серединній зоні діафіза кістки, що розвивається, остеопластичний процес здобуває поступово меншу інтенсивність, чим у метафізах.
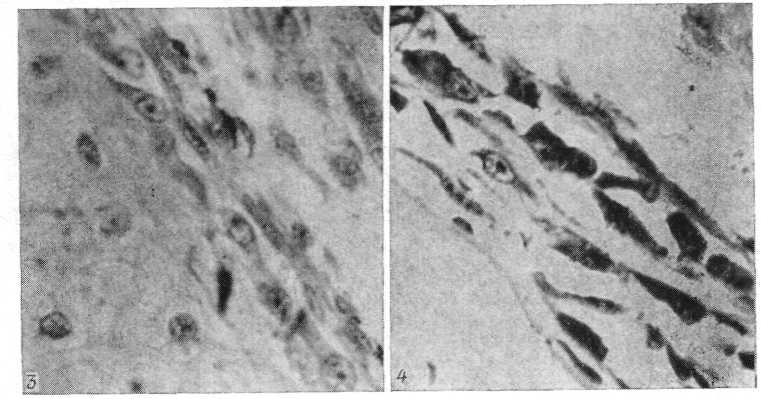
Рис. 6. Періост центра диафиза стегнової кісти R. temporarіa на 47-й стадії розвитку. Відзначаються одиничні остеоцити в периостальній кістці (гематоксилін Майера - еозин; про. 40, ок. 7).
Рис. 7. Періост зони діафіза кісти жаби на 49-й стадії розвитку. Відзначається вогнище енхондральної резорбції, у якому вже помітні зачатки миєлогенного ретикулума (гематоксилін - еозин; про. 40, ок. 7).
Клітини другого (зовнішнього) ряду морфологічно більш різнорідні. Серед них зустрічаються преостеобластические стадії з порівняно великим ядром. І ті й інші зв'язані з коллагеновым каркасом, утвореним, очевидно, їхньою участю.
Кількісний розподіл кліток у різних зонах періосту характеризується наступними показниками: у діафізарної області протягом 340 мкм утримується 11 остеобластів і 10 фібробластоподібних кліток.
У личинок трав'яної жаби 49-й стадії розвитку в стегновій кісті помітне насамперед збільшення в її розмірах по довжині й у поперечнику. Перихондральна кісткова трубка добре сформована. У центрі діафіза в хрящовому стрижні до цього часу з'явилося вогнище енхондральної резорбції, у якому вже помітні зачатки мієлогенного ретикулума (мал. 7). У проксимальному і дистальному відділах хрящового стрижня чітко виступає зональна диференціювання хондроцитів, у зв'язку з чим по довжині хрящового стрижня виділяються території: епіфізів, пролиферируючих хондроцитів, гіпертрофованих хондроцитів, що відрізняються своєї цитоархітектонікой.
Періост також здобуває в масі переважно за рахунок збільшення кількості остеобластів, що розташовуються, як правило, у два ряди, і фібробластоподобних клітин. Цей показник відповідає помітної інтенсифікації остеопластичного процесу в зв'язку з прискоренням темпів росту кісткової ланки і кістяка в цілому. Товщина періосту зберігається більш-менш рівномірної на всьому протязі і складає приблизно 50 мкм.
На початковій стадії метаморфоза личинки трав'яної жаби мають уже сформовані трубчасті кісти в тазовій кінцівці. Періост у них зберігає вид порівняно тонкої по масі структури, що включає 3 (не більш 4) ряди клітин, укріплених фібриллярним каркасом. Помітною ознакою, що з'явилася до цього часу в періості, виступає не стільки його загальна структура, скільки стан клітин. На відміну від періосту в личинок жаб на цій стадії онтогенезу остеобласти в періості здобувають форму подовжених кліток із щільним штриховидним ядром (мал. 8). Фактично клітки, що обмежують фронт периостального остеогенезу, за формою мало відрізняються від більш поверхово розташованих клітин періосту. Такий метаморфоз остеогенних клітин зв'язаний, очевидно, з переходом періосту в стан зрілого дифінітивного тихорєцького диференціювання, для якої подовжена форма остеогенних клітин є звичайною. Якби зміни у формі остеобластів були сполучені зі зниженням інтенсивності остеопластичного процесу, то вони проявилися б у різних ділянках періосту нерівномірно, оскільки периостальний остеопластичний процес відбувається в різних зонах кістки з неоднаковою інтенсивністю. Про це ж свідчить і чисто кількісний показник розподілу остеобластів. Незважаючи на зміну форми, їхня кількість на еквівалентній довжині періосту залишається в межах 9-10 одиниць на 34 мкм.
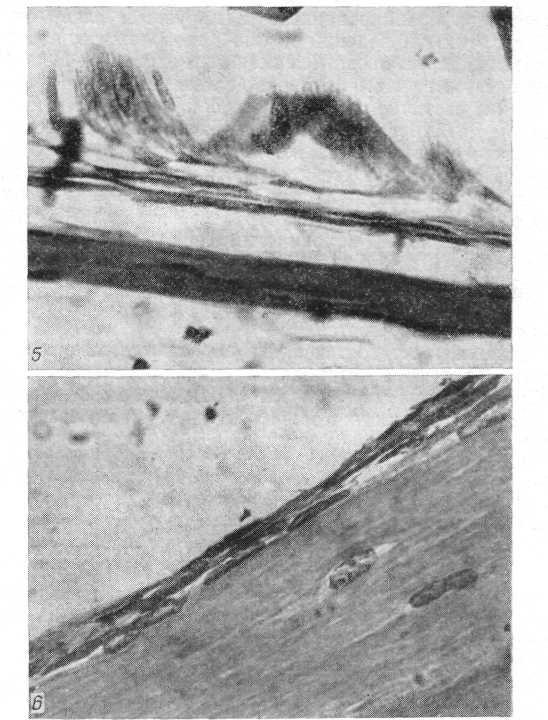
Рис. 8. Періост проксимального метафиза стегнової кісти жаби після завершення метаморфоза. Видні подовжені за формою остеобласти (гематоксилін Майера - эозин; про. 40, ок. 7).
Рис. 9. У сформованій стегновій кісті статевозрілої R. temporarіa окістя представлене одношаровою структурою (гематоксилін Майера - эозин; про. 90, ок. 7).
Природно, виникає питання: яким шляхом поповнюється склад остеогенних клітин у періості? На гістологічних зрізах довгих кіст трав'яної жаби личинокових стадій розвитку в періості ми завжди знаходили клітки в стані мітозу. Частка цих кліток невелика: серед остеобластів мітотичний індекс не перевищує 0,2 %, а для кліток менш диференційованих (у зовнішньому шарі) він досягає 1,0-1,5 %. З цих спостережень можна припустити, що періост самопідтримується тканинними системам. Поповнення кліток у ньому відбувається переважно за рахунок менш диференційованих станів і частково за рахунок функціонально зрілих остеобластів.
З початком метаморфоза, коли стегнова кіста в зростаючій кінцівці вже майже сформована, періост у діафізарної області переходить у зрілий стан і його клітки, включаючи остеобласти, здобувають компактну-подовжену форму з порівняно дрібним щільним ядром також подовженої форми. У структурі періосту до цього часу підсилюється колагеновий волокнистий каркас, і весь періост стає більш щільним і міцним. Зрілий періост можна тому, додаючи визначене зусилля, відокремити від кісти у виді тонкої плівки, чого не можна зробити з періостом тільки формується, при такій спробі він звичайно руйнується.
У кістках, що закінчили ріст, або в періоди сезонного припинення росту клітки періосту стають морфологічно однотипними (мал. 9). Серед них не зустрічаються з ознаками функціонуючих остеобластів, хоча всі клітки такого періосту є потенційно остеогенними. При зміні умов у ньому можуть активізуватися остеопластичні функції з появою морфологічно ідентифікованих остеобластів у зонах поновлення процесів ремоделювання кістки або її росту.
З цієї причини в дорослих жаб у період літнього активного стану завжди виявляються зональні особливості в структурі періосту, кількості в ньому клітинних рядів, а також формі і розмірах самих клітин. При цьому в діафізарної області структура періосту істотно не змінюється, оскільки периостальний ріст кістки в діафізі в дорослих жаб практично припиняється.
Таким чином, у ростучих кістках скелету амфібій окістя представлене складною структурою, що складається з остеогенних клітин, що знаходяться на різних стадіях диференціювання. У кістках сформованого і ріст кістяка, що закінчився, вона перетворюється в одношарову структуру, що складається з зовні однотипових фібробластоподобних клітин.
Розділ 3. Пролиферативні властивості клітин окістя в амфібій
У спеціальній літературі є нечисленні загальні відомості про розвиток кістяка довгих трубчастих кісток в амфібій (Кащенко, 1896; Eggelіng, 1911; Lubosch, 1928; Румянцев, 1958; Житников, 1980; Домашевская, 1984, 1989). При цьому фактично відсутня інформація про проліферативних потенції кліток окістя, що відіграє велику роль в апозиційному рості кістки. Для більш повного з'ясування особливостей росту і розвитку кістяка в амфібій представляється важливим вивчення не тільки структури окістя, але і проліферативних потенцій її клітин.
Дослідження проводили на личинках, маса яких складала 4-5 м, і на однорічних часночницах (Pelobates fuscus). 3Н-тимидин уводили внутрішньочеревно однократно в дозі 74 кбк/м живої маси з наступною фіксацією періосту з підлягаючим шаром компакт стегнової кістки в 10 %-м нейтральному формаліні через 1; 24 і 48 год. (личинки) і через 1; 24 і 72 год. (жабенята).
На гістоавтографах з 3Н-тимидином визначали (у %) відношення мічених кліток до загального числа кліток на умовну одиницю площі гістологічного зрізу, а також інтенсивність синтезу дезоксирибонуклеїнової кислоти по концентрації мітки над ядром клітки (інтенсивність мічення). Мітотичний індекс ядер обчислювався на 1000 клітин. Отримані дані обробляли методом варіаційної статистики (Терентьєв, Ростова, 1977).
У період розвитку личинки (стадія 46, 47, 49 по Н. В. Дабагяну) окістя довгих трубчастих кіст у зонах закладки кісти діафіз, метафіз і "перехідна зона" різна по товщині (кількості клітинних рядів) і функціональним проявам кліток. Так, у личинок (на стадії 46) окістя починає диференціюватися в діафізарній зоні майбутньої кісти в період, коли хрящова закладка підготовлена до заміщення. У найглибшому шарі перехондра ще до появи тут кісткової манжетки клітки здобувають характерний вид остеобластів і починають продукувати фермент - лужну фосфатазу. У цей час у земноводних відзначається типове для амфібій грибоподібне розростання хрящового епіфіза, краю якого далеко заходять на метафізи. Клітки окістя в "перехідній зоні" (на границі перихондр -і окістя) розташовані в 2-3 ряду, з яких внутрішній шар складається з остеобластів, а поверхневий - хондробластів. У кінцевому верхівковому відділі щілини, куди край кісткової трубки не доходить, утримуються тільки хондробласти (малюнок, а).
Уже через годину після введення 3Н-тимидина індекс мічення клітин, як по шарах перихондра, так і по зонах хрящової закладки кісти, неоднаковий (малюнок, б): він, по-перше, значно вище в зоні діафізу закладки кістки і, по-друге, великий відсоток мічених ядер відзначається в його поверхневому шарі. Варто звернути увагу на те, що і показник мітотичного розподілу цих клітин вищий.
Очевидно, сумарний показник концентрації всіх клітин окістя, як і фракції в кожній зоні закладки кістки, безпосередньо зв'язаний з інтенсивністю протекаючого тут перихондрально-періостального остеопластичного процесу. Тому що в метафізарній зоні приріст йде повільно, те тут і зосереджено на еквівалентній площі мінімальна кількість проліферуючих клітин.
Приблизно така ж картина розподілу кліток у зоні метафиза зберігається і через 24 год. після введення 3Н-тимидина (личинки досягають 47-й стадії розвитку), однак до цього часу вагомо (майже в 2 рази) зменшується інтенсивність мічення кліток поверхневого шару - свідчення розподілу, що пройшли за цей час їх, (від 13 до 27 %).
У цей час перихондральна кісткова трубка в області діафаза здобуває трохи великі розміри по довжині і масі. Якщо на попередній стадії зачаток кісткового діафізу виглядав на гістологічному зрізі як тонка (10-12 мкм) пластинка, то на стадії 47 товщина її досягає 24 мкм, і в ній уже є одиничні остеоцити.
Паралельно з цим слід зазначити кількісне збільшення мічених остеобластів (з 17 до 38 %) і незначна кількість мічених фібробластоподобних клітин. Однак якщо збільшення мічених остеобластів відбулося за рахунок дозрівання як позначених преостеобластів, так і розподілу самих остеобластів, ту утримуючу мітку фібробластоподібні клітки розподіляються в межах своєї ж популяції поверхневого шару.
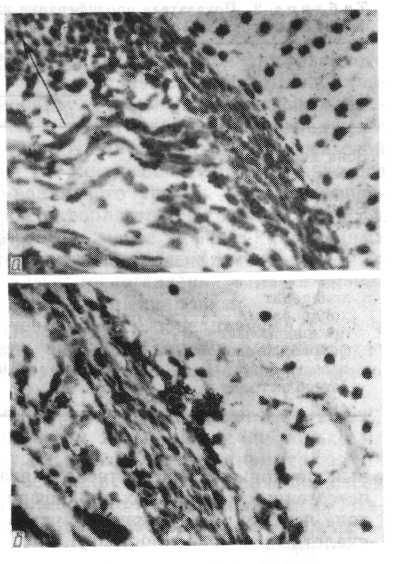
Гістоавтографи окістя стегнової кісти личинки P. fuscus на стадії 47 (по Н. В. Дабагяну). Видні численні мічені клітки окістя (а) і окістя зони діафізу (в) через 1 год. після внутріочеревного введення 3Н-тимідину (Гемат.-еозин Делафильда; про. 40. ок. 7).
Через 48 год. після введення 3Н-тимидина можна відзначити наступне: 1) помітно прогресує фракція мічених кліток у глибокому шарі (зона діафізу) при одночасному значимому зниженні індексу мічення кліток поверхневого шару, тобто поповнення остеобластів, очевидно, відбувається за рахунок кліток поверхневого шару періоду; 2) частина мічених кліток метафіза розділилася повторно. Мітка над ядрами деяких кліток стала на рівні тла, і вони не враховувалися при підрахунку.
Отже, сам собою напрошується висновок про те, що інтенсивність остеопластичного процесу в зростаючій закладці кістки в даний період онтогенезу тісно зв'язана з кількісною концентрацією остеогенних клітин і їх проліферуючих одиниць у кожній конкретній зоні періхондра - періосту, що в кінцевому рахунку і позначиться на росту кістякових ланок у цілому.
Для того щоб упевнитися в можливості цього, ми поставили додатковий досвід на однорічних чесночницях при тих же режимах введення 3Н-тимидина в організмі. Перед початком викладу результатів даного дослідження відзначимо, що в річної жаби діафізарна область стегнової кістки вже цілком сформовані, ростові процеси зберігаються ще в метафізах і значно активно протікають у перехідній зоні.
На відміну від окістя більш ранніх стадій розвитку личинок у жаб на цій стадії онтогенезу остеобласті в періості діафіза вже здобувають форму подовжених кліток із щільним штриховидним ядром. Такий метаморфоз остеогенних клітин, очевидно, зв'язаний з переходом окістя діафізарної зони кістки в стан більш зрілої тихорєцької диференціювання, для якої подовжена форма остеогенних клітин є звичайною. Незважаючи на зміну форми, їхня кількість на рівній довжині періосту зберігається в межах 17-20 одиниць на 250 мкм.
Природно, виникає питання: яким шляхом поповнюється склад остеогенних клітин в окісті, за рахунок яких джерел?
При аналізі отриманих гістоавтографів виявилося, що через годину після введення 3Н-тимидина в окісті діафізу відзначаються усього одиничні (до 6%) мічені клітини серед преостеобластів. В окісті зони метафізів реєструються мічені клітини усіх видів зрілості (остеобласти, преостеобласти і фібробластоподібні клітини), однак їхній відсоток невисокий (від 5 до 7 %). У той же час у "перехідній зоні" індекси мічення ядер над цими видами кліток трохи вище, а над преостеобластами і фібробластоподобними клітками індекси мічення ядер майже в 3 рази вище, ніж в окісті зони метафіза кістки.
Через 24 год. після початку досліду в зоні діафіза як і раніше реєструються невисокі відсотки мічених ядер над клітинами - 2-4 %, у той час як над ядрами клітин метафіза і перехідної зони - значно вище. Так, над прехондробластами індекс мічення ядер - 18 %, а над преостеобластами - 53%. При цьому відзначається зниження на половину інтенсивності мітки (з 60 до 22 зерен відновленого срібла), через 24 год. ми врахували фактично популяцію поділившихся раніше позначених клітин.
У метаепіфізарних зонах під грибоподібною муфтою епіфізарного хряща продовжує існувати періостально-перихондральна ділянка, у якому на границі з гіаліновим хрящем клітки такого тихорєцького утворення диференціюються в хондробласти, а на границі з кістою і кровоносними судинами - в остеобласти. Поверхневий шар окістя, як і на попередніх стадіях, складається з волокнистого каркаса з включеними клітками різного рівня диференціювання - від фібробластоподобних до преостеобластів. Поповнення кліток у ній відбувається переважно за рахунок менш диференційованих станів, і частково за рахунок функціонально зрілих остеобластів. Тобто , окістя в земноводних тут представлена єдиним шаром: входять до складу клітини різних стадій диференціювання.
При аналізі гистоавтографов через 72 год. у зоні діафізу кістки в окісті відзначається як і раніше невисокий індекс мічення ядер клітин. Серед ядер клітин окістя зони метафізів індекси мічення ядер також невисокі.
При цьому над фібробластоподобними клітинами відзначається і розведення мітки, тобто має місце, очевидно, не тільки їхній розподіл, але і трансформація в інші ряди клітин, наприклад, за їхній рахунок йде поповнення популяції преостеобластів. Зниження індексів мічення ядер спостерігається і над ядрами клітин перехідної зони, з одного боку, за рахунок зменшення прехондробластів ми маємо збільшення відсотка хондробластів, а з іншого боку - за рахунок раніше мічених преостеобластів - значний ще відсоток мічених остеобластів (19 %).
Преостеобластичні стани, зважаючи на все, являють собою основний клітинний фонд зростаючого окістя, тому що фібробластоподібні форми пролиферируют (по радіоактивному індексі) у межах 16-19%. Картина гісторадіографії цікава не стільки цими даними, скільки можливістю вірогідно переконатися в гістогенетичній наступності різно диференційованих клітин окістя. Справді, протягом доби можна відзначити достовірне поповнення фракції мічених остеобластів за рахунок диференціювання раніше мічених преостеобластів, що помітно по чисельному збільшенню остеобластів з половинною інтенсивністю мітки, характерної для преостеобластів, що розділилися. Однак навіть в умовах короткочасного досліду помітна тенденція переходу невеликої частки позначених клітин фібробластоподобних у шар преостеобластів. Тенденція домінуючого проліферування менш диференційованих клітин для поповнення складу більш диференційованих кліток самого внутрішнього шару добре помітна ще в перихондрі до початку периостального остеогенезу. На наступних етапах спостереження (через 24, 48 год.), коли вільно циркулює в організмі 3Н-тимидина вже нема, помітно прогресує фракція мічених клітин у глибокому шарі при одночасному значимому зниженні індексу мічення клітин поверхневого шару. При цьому слід зазначити, що ключовий етап периостального остеогенезу завжди починається з налагодженням остеогенної функції в клітках перихондра, тому структуру перихондра є підстави вважати як вихідну для диференціювання в періост. Але зовсім ясним варто визнати і те, що навіть у сформованій зоні кісткового кістяка (діафізі) клітини окістя якоюсь мірою зберігають проліферативні властивості, можливо, необхідні не стільки для росту кісти, скільки для її адаптивного ремоделировання і фізіологічної регенерації.
У спеціальній літературі немає даних про проліферативні властивості клітин окістя в земноводних. Такі відомості отримані методом авторадіографії з міченим попередником ДНК - 3Н-тимідином для ссавців (Owen, 1963; Tonna, 1965; Мажуга, Хрисан-фова, 1985; Родіонова, 1989). У цих роботах відзначається низька проліферативна активність клітин поверхневого шару окістя в пацюка.
Отримані нами дані дають підстави погодитись з припущенням інших авторів (Левикова, 1950; Prіchard, 1952; Burger et al. 1986), які допускають можливу участь клітин поверхневого шару окістя в поповненні його остеогенних клітин і якоюсь мірою доповнюють наявні в спеціальній літературі дані про властивості проліферації клітин окістя в інших хребетних.
Висновок
Для безхвостих земноводних, як і для всіх хребетних тварин характерний процес утворення кісткової тканини. В організмі безхребетних існує два різновиди остеогенезу — безпосередньо із мезенхіми та заміни кісткою гіалінового хряща. Ділянки мезенхіми, в яких почався остеогенез за зовнішнім виглядом являють собою мембрани, осифікація у них називається інтрамембранною. Цей остеогенез характерний для розвитку кісток черепа та нижньої щелепи, на місці, де утворюється кістка, спочатку знаходиться шар рихлої мезенхіми, вона має вигляд зірчастих клітин з відростками, які вступають в контакт з відростками сусідніх клітин мезенхіми, утворюючи сітку, остання занурена в аморфну міжклітинну речовину, що містить окремі пучки колагенових волокон і кровоносні судини.
У ростучих кістках скелету амфібій окістя представлене складною структурою, що складається з остеогенних клітин, що знаходяться на різних стадіях диференціювання. У кістках сформованого і ріст кістяка, що закінчився, вона перетворюється в одношарову структуру, що складається з зовні однотипових фібробластоподобних клітин.
Вивчення проліферативних властивостей клітин окістя у безхвостих проводять за допомогою спеціальних приладів – гіспоавтографів. При цьому використовують мічення клітин.
Література
Бродский В.Я., Урываева И.В. Клеточная полиплоидия. Пролиферация и дифференцировка. — М.: «Наука», 1981. — 259 с.
Вечерская Т. П. Пролиферативные свойства остеогенных клеток в растущем скелете птиц и млекопитающих: Тез. докл. Респ. научн. конф., поев. 150-летию со дня рождения В. А. Беца. - Киев, 1984. - 95 с.
Дабагян Н. В., Слепцова Л. А. Таблица нормального развития травяной лягушки Rana temporaria // Объекты биологии развития.— М.: Наука, 1975.— С. 442 — 463.
Домашевская Е. И. Особенности строения надкостницы у некоторых позвоночных // Вестн. зоологии.— 1984,—№ 4 —С. 21—34.
Домашевская Е. И. Особенности строения надкостницы у безхвостых амфибий // Вестн. зоологии. — 1982.—№ 2.—С. 42—46.
Житников А. Я. Морфология хондрогенеза и кинетика репродукции хондроцитов при развитии хрящевого скелета//Докл. АН УССР. Сер. Б. - 1980.—№ 7, — С. 69 — 74.
Кащенко Н. Ф. Что такое мезенхима?// Изв. Томск, ун-та.— 1896.— № 10. — С. 1 — 24.
Левикова В. П. Наблюдения над экспериментальным остеогенезом у кролика // Докл. АН СССР.—1950.—71, № 1.—С. 149—152.
Мажуга П.М., Домашевская Е.И. Развитие и структура надкостницы у наземных позвоночных. — К.: «Наукова думка», 1990. — 117 с.
Мажуга П. М. Кровеносные капилляры и ретикулоэндотелиальная система костного мозга.— Киев: Наук, думка, 1978.— 190 с.
Мажура П. М., Хрисанфова Е. Н. Очерки эволюции человека.— Киев: Наук, думка, 1985.— 136 с.
Родионова Н. В. Функциональная морфология клеток в остеогенезе.— Киев: Наук, думка, 1989.— 185 с.
Румянцев А. В. Опыт исследования эволюции хрящевой и костной тканей.— М.: Медицина, 1958.—439 с.
Терентьев П. В. Лягушка.— М.: Сов. наука.— 1950.— 345 с.