Регуляция биосинтеза белков на этапе транскрипции
Регуляция биосинтеза белков на этапе транскрипции
Введение
Проблемы, связанные с регуляцией метаболических процессов – важнейшие в системе биохимических знаний, и делятся на два больших класса:
Представления о молекулярных механизмах процессов регуляции, которые являются, скорее, предметом молекулярной биологии и, частично, молекулярной генетики.
Феноменологическое выражение последствий регуляторных событий, определяющее направление протекания биохимических процессов, что, собственно, и является предметом биохимии. Именно последний аспект проблемы будет лежать в основе представленного материала, но, учитывая небольшой объем пособия, ограничимся лишь некоторыми ключевыми вопросами. Для более детального ознакомления с проблемой следует обратиться к фундаментальным трудам.
1. Основные определения
Регуляцией метаболизма называется управление скоростью биохимических процессов путем обратимого изменения количества белковых посредников, участвующих в этих процессах, или их активности.
Белковый посредник – более общий и точный термин, чем термин фермент. Хотя во многих биохимических процессах белковые посредники представляют собой именно ферменты – катализаторы химических превращений субстратов, но, например, в транспорте субстратов через биологические мембраны перенос опосредуется белками, не являющимися ферментами, так как они не катализируют каких-либо химических реакций, а обеспечивают узнавание и транслокацию субстратов.
В отдельных случаях роль посредника выполняют не белки, а рибонуклеопротеиды или сама РНК, но это, скорее, исключение.
В соответствии с приведенным определением следует выделять два основных уровня регуляции: биосинтеза белковых посредников; их активности.
Оба уровня жизненно необходимы для организма, и мутанты с нарушением хотя бы одного, как правило, вытесняются из популяции.
2. Регуляция биосинтеза белков
Биосинтез белков складывается из процессов непосредственного построения и модификации белковой молекулы, а также из «подготовительных» процессов: репликации генетического материала и его транскрипции.
Репликация ДНК подробно рассматривается в курсах молекулярной биологии и фундаментальных учебниках биохимии. Мы остановимся лишь на некоторых принципиальных вопросах, имеющих значение для регуляции биосинтеза белка.
Необходимо отметить, что термин хромосома как место локализации ДНК применим только к клеткам эукариот, тогда как часто используемый термин бактериальная хромосома неточен и лучше говорить о генофоре или нуклеоиде, подразумевая под этими терминами ДНК-РНК-белковый комплекс.
Еще точнее термин геномный эквивалент, так как часть ДНК в бактериальной клетке присутствует в нескольких копиях. Если в бактериальную клетку поступает дополнительный фрагмент ДНК, несущий новые аллели тех же локусов, то возникает так называемый меродиплоид.
3. Особенности процесса репликации
Как и в случае биосинтеза других биополимеров, процесс репликации ДНК включает три этапа: инициацию, элонгацию и тер-минацию. Для репликации характерны следующие особенности:
1. Она осуществляется по полуконсервативному механизму, причем цепи ДНК антипараллельны (З'конец содержит остаток ортофосфата — Р, а 5' конец — ОН) и последующей транскрипции подвергается только одна цепь:
Синтез, по-видимому, протекает прерывисто и в направлении 5' 3'.
Каждая цепь начинается с РНК-затравки, фрагменты объединяются ДНК-лигазой с выщеплением РНК.
Система репликации является мультиферментной, в нее входят 2–3 ДНК-полимеразы, ДНК-лигаза, топоизомеразы, необходимые для расплетания цепей ДНК и последующей их сверхспирализации. Всего порядка 15 генетических локусов кодируют тот или иной полипептид, необходимый для репликации.
Существуют четыре основных типа: синтез фрагментов, репликация плазмид, рекомбинационный синтез ДНК, репарационный синтез ДНК. В каждом из них участвуют как общие, так и специфические компоненты.
Нативная система репликации ДНК является мембранной и инактивируется при разрушении мембран. Поэтому для моделирования процесса репликации используются, как правило, упрощенные системы, полученные из бактериальных «условных» мутантов или из «маленьких» фагов.
Регуляция процесса репликации ДНК наиболее строго осуществляется на этапе инициации. Репликация находится под положительным и отрицательным контролем, причем оба они скоординированы с клеточным делением.
В общих чертах принцип такой регуляции можно сформулировать следующим образом. При положительном контроле происходит накопление активатора репликации до порогового, достаточного для инициации нового цикла репликации уровня. Пороговый уровень достигается при удвоении клеточной массы так, чтобы во вновь образовавшихся клетках процесс репликации был бы уже запущен и успел завершиться к моменту нового деления. При отрицательном контроле происходит накопление ингибитора инициации репликации, который должен синтезироваться лишь в ограниченном количестве вскоре после начала предыдущего цикла репликации. Такой ингибитор может быть продуктом гена, локализованного вблизи от точки начала репликации, транскрипция которого осушествляется только в период репликации данного участка ДНК. В процессе роста клетки ингибитор «разбавляется», а к моменту удвоения массы клетки уровень его падает ниже критического, что позволяет клетке инициировать новый цикл репликации. Взаимодействие этих двух механизмов и должно координировать процессы репликации ДНК и деления клетки.
Реальные механизмы регуляции репликации еще не расшифрованы. У эукариот определенную роль в регуляции репликации, по-видимому, играют гистоны.
4. Транскрипция генетической информации
Этот этап биосинтеза белков состоит в «переписывании» информации, закодированной в ДНК, на олигонуклеотидную последовательность информационной РНК и осуществляется РНК-полимеразой. Строение этого фермента лучше всего изучено у бактерий Escherichia. Это сложный белок с молекулярной массой 450 кДа. Его сердцевина включает две идентичные а-субъединицы и две различные р-субъединицы. В состав «полного» фермента входит также а-субъединица, ответственная за процесс узнавания промотора и инициацию транскрипции. Наконец, существует еще один белковый компонент, обозначаемый р-фактор, который ответственен за правильную терми-нацию транскрипции. Таким образом, РНКП Escherichia coli и ряда других грамотрицательных бактерий выглядит следующим образом:
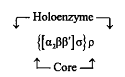
У грамположительных бактерий РНКП устроена еще сложнее. Например, в случае Bacillus sub>tilis она может содержать несколько а-факторов, а у ряда архебактерий РНКП состоит из 9–10 компонентов, приближаясь по сложности строения к РНКП эукариот.
В клетках эукариот обнаружено по крайней мере три типа РНКП. Полимераза I находится в ядрышке и транскрибирует гены большинства рибосомных РНК; полимераза II – в нуклеоплазме и транскрибирует большую часть других генов; полимераза III – гены транспортных РНК и одной из рРНК. Кроме того, в митохондриях и хлоропластах эукариот присутствуют собственные РНКП.
Между тем собственно полимеразная реакция может осуществляться гораздо более простыми ферментами. Так, РНКП «нечетных» ТЗ и Т7 фагов Escherichia coli состоит из единственного полипептида с молекулярной массой ПО кДа, а митохондриальная РНКП представляет собой полипептид с молекулярной массой 64 кДа.
По-видимому, сложное устройство бактериальной и, особенно, эукариотических РНКП, с одной стороны, обусловлено необходимостью «узнавать» большое число промоторов, а с другой – позволяет осуществлять многообразную регуляцию транскрипции в процессе функционирования этого фермента.
5. Регуляция процесса транскрипции
Исходя из возможности управления синтезом белковых посредников на этапе транскрипции, их можно разделить на три основные группы:
конститутивные белки, синтез которых не зависит от наличия субстратов и продуктов;
индуцибельные белки – их синтез ускоряется в присутствии субстратов;
репрессибельные белки, синтез которых подавляется избытком конечного продукта данного метаболического пути.
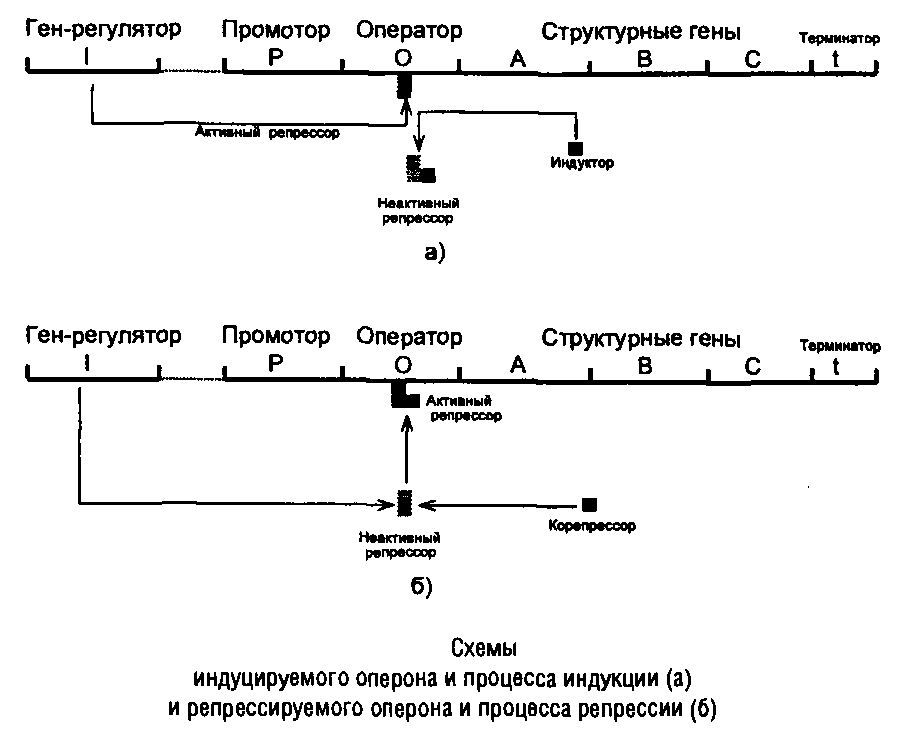
Регуляция на этапе инициации транскрипции. В 1960-е годы Ф. Жакоб и Ж. Моно установили, что в явлениях индукции и репрессии принимают участие белковые факторы – репрессоры, продукты специальных генетических элементов – генов-регуляторов, способные в определенных условиях тормозить процесс транскрипции на этапе инициации, поэтому оба этих типа регуляции относят к негативным.
В индуцируемом опероне ген-регулятор кодирует активный реп-рессор, который блокирует транскрипцию, взаимодействуя с операторным участком ДНК и препятствуя продвижению РНКП. Индуктор, представляющий собой исходный субстрат данного метаболического пути или близкое к нему соединение, способен взаимодействовать с репрессором и инактивировать его, освобождая таким образом операторный участок ДНК. В результате РНКП начинает транскрипцию данного оперона. Эти события отражены в схеме.
В репрессируемом опероне ген-регулятор кодирует неактивный репрессор, который может переходить в активное состояние и блокировать транскрипцию, соединяясь с оператором, только после взаимодействия с избытком конечного продукта данного метаболического пути. Процесс отражен в схеме.
Большую роль в регуляции транскрипции играет так называемая катаболитная репрессия, которая проявляется в диауксии в процессе роста бактерий. Феномен диауксии обнаруживается, когда в среде присутствуют два субстрата, причем ферменты, осуществляющие катаболизм одного из них, индуцибельны, а ферменты, осуществляющие катаболизм другого, конститутивны. В этом случае сначала потребляется только глюкоза, тогда как индукция лактозных ферментов не происходит до тех пор, пока не будет потреблена основная часть глюкозы. Это отражается во временном замедлении роста культуры на тот период, который необходим для индукции и синтеза р-галактозидазы. Таким образом, несмотря на присутствие в среде индуктора, альтернативный субстрат препятствует индукции.
Механизм явления катаболитной репрессии состоит в следующем. Для индукции некоторых «слабых» оперонов, в том числе Дзс-оперона, недостаточно инактивации отрицательного регулятора – репрессора. Необходимо и участие положительного регулятора, представляющего собой комплекс специального активирующего белка с циклической AMP. Этот белок, активирующий транскрипцию, получил название БАК-белка или «белка, активирующего катаболитные гены». БАК-белок представляет собой димер с молекулярной массой 45 кДа. Под действием сАМР он подвергается конформационным изменениям и приобретает повышенную способность связываться с промотором. Полагают, что присоединение комплекса сАМР-БАК к ДНК ослабляет спаривание Г-Ц-оснований, способствует частичному разделению спиралей ДНК и облегчает формирование инициирующего транскрипцию комплекса РНКП с ДНК. Уровень сАМР в клетке обратно пропорционален уровню АТР, и в присутствии легко метаболизируемых субстратов, способствующих повышению уровня АТР, сАМР «не хватает» для образования комплекса с БАК.
Тонкие механизмы регуляции уровня сАМР связаны с функционированием фосфотрансферазной системы транспорта Сахаров и будут рассмотрены в главе, посвященной регуляции процессов мембранного транспорта.
Необходимо отметить, что у ряда бактерий роль глюкозы в катаболитной репрессии могут выполнять другие источники энергии, которые в этом случае тормозят катаболизм глюкозы.
В индуцибельных оперонах возможны и другие типы положительной регуляции, независимой от сАМР. Например, в арабинозном опероне Escherichia coti арабиноза не просто инактивирует репрессор, но превращает его в положительный регулятор. Аналогичное явление обнаружено в случае оперонов галактозы и рамнозы.
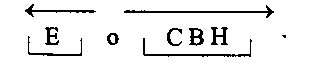
В дивергентных регулонах транскрипция протекает в разных направлениях и может быть некоординированной, т.е. осуществляться с разной скоростью. При этом возможно считывание с разных цепей ДНК. Примером служит аргининовый оперон: в его части, включающей 4 гена из 9, транскрипция трех генов осуществляется в одном направлении, а транскрипция другого гена – в противоположном:
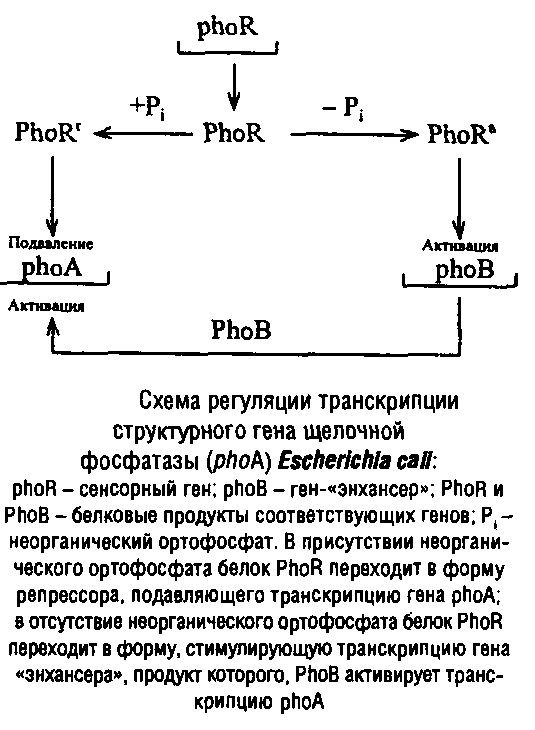
Еще одним примером положительной регуляции процесса транскрипции является регуляция с участием генов – «энхансеров». Ранее считали, что этот тип регуляции характерен только для эукариот. Но в последнее время формально сходные механизмы обнаружены и у прокариот.
Особенность генов – «энхансеров» в том, что они проявляют свою стимулирующую активность независимо от ориентации и расположения относительно активируемого гена: могут находиться перед геном, за ним и даже внутри него.
Продуктами генов – «энхансеров» являются белки с молекулярной массой 25–30 кДа, способные связываться с промоторной областью. Как правило, такая система двухкомпонентна и включает «сигнальный» белок, способный «чувствовать» изменение условий окружающей среды и стимулировать синтез другого белка – «активатора», который и запускает транскрипцию искомого белкового посредника.
Перечисленные механизмы регуляции транскрипции на стадии инициации достаточно быстро реагируют на изменение внешних условий, однако управляют работой одного или небольшого числа оперонов в каждый момент времени.
Наряду с ними существуют механизмы системной регуляции, связанные с изменением функционирования одновременно большого числа оперонов.
В клетках эукариот это достигается путем конформационных перестроек хроматина, процессинга и РНК, а также за счет управления трансляцией путем формирования так называемых информосом.
В клетках прокариот системная регуляция осуществляется путем модификации специфичности работы РНКП посредством изменения ее компонентного состава. Эти механизмы вступают в действие, когда нужно активировать одновременно большое число новых промоторов или сменить матрицу. Последний случай изучен наиболее подробно на примере фагов Escherichia coii и Bacillus sub>tilis.
В геноме бактериофагов присутствуют три типа генов: ранние, средние и поздние, классифицируемые на основании порядка их транскрипции в ходе развития фага. Ранние гены всегда транскрибируются РНКП клетки-хозяина, а в случае транскрипции средних и поздних генов возможно несколько вариантов:
среди продуктов ранних генов присутствует собственная фаговая РНКП, а также белки-ингибиторы РНКП хозяина. Таким образом, происходит смена РНКП;
среди продуктов ранних генов присутствуют новые а-факторы, взаимодействующие с «сердцевиной» РНКП хозяина и обеспечивающие транскрипцию средних генов. Среди продуктов средних генов, в свою очередь, присутствуют белки с функциями а-факторов, которые обеспечивают транскрипцию поздних генов, Таким образом, происходит смена о-факторов;
3) среди продуктов «ранних» генов присутствуют белки, модифицирующие «сердцевину» РНКП хозяина, тогда как а-фактор хозяина сохраняется.
Регуляция транскрипции путем образования специфических а-факторов широко распространена и в бактериальных системах.
Особый а-фактор контролирует транскрипцию ряда генов азотного метаболизма. Наконец, существуют промоторы, активирующиеся только при 50°, в их транскрипции участвует фактор аЕ.
Примером системной регуляции является и регуляция процесса спорообразования у бацилл. В этот процесс вовлекаются сотни локусов, организованные в специальные структуры и разбросанные по всей молекуле ДНК. Одним из способов переключения роста на спорообразование служит изменение РНКП и, в первую очередь, ее о-фактора. Такая РНКП преимущественно транскрибирует «ранние» гены спорообразования, важную роль в котором играет также регуляция на уровне трансляции.
Другой пример системной регуляции у прокариот: при воздействии на микроорганизмы нагревания до супраоптимальной температуры наступает тепловой шок, вызывающий координированную индукцию «белков теплового шока». Транскрипция их локусов и образование главного фактора а осуществляется под контролем минорного фактора с32.
Этот механизм обеспечивает координацию процессов транскрипции и трансляции и подробнее будет обсуждаться при рассмотрении способов управления скоростью роста клеток.
6. Регуляция на этапе терминации транскрипции
РНКП может «узнавать» специфические последовательности ДНК, сигнализирующие об окончании транскрипции. Эта ее способность усиливается или модифицируется под влиянием особого полипептида – р-фактора, обеспечивающего нормальную терминацию.
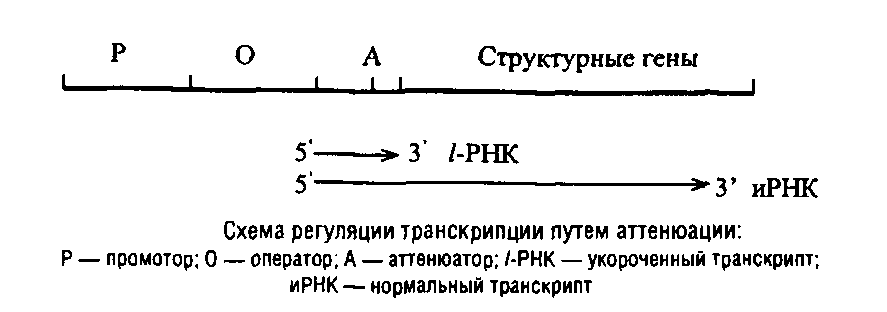
Наряду с этим в составе ряда регулонов обнаружены так называемые аттенюаторы, т.е. участки ДНК, вызывающие преждевременную терминацию, также р-зависимую. Эти генетические элементы располагаются между оператором и первым структурным геном в регулонах, контролирующих биосинтез ряда аминокислот. Все аминокислотные регулоны, управляемые с помощью аттенюации, характеризуются обогащением участка и РНК, соответствующего расстоянию до аттенюатора кодонами той аминокислоты, которая служит отрицательным эффектором.
Регуляция путем аттенюации основана на тесном сопряжении у прокариот транскрипции и трансляции, которые, как правило, протекают одновременно. В процессе трансляции синтезируемой иРНК рибосома, экранирующая около 10 нуклеотидов, может существенно влиять на конформацию транслируемой иРНК, а та, в свою очередь, на продвижение РНКП по матрице ДНК. При формировании «критической» конформации иРНК наступает диссоциация РНКП от ДНК.
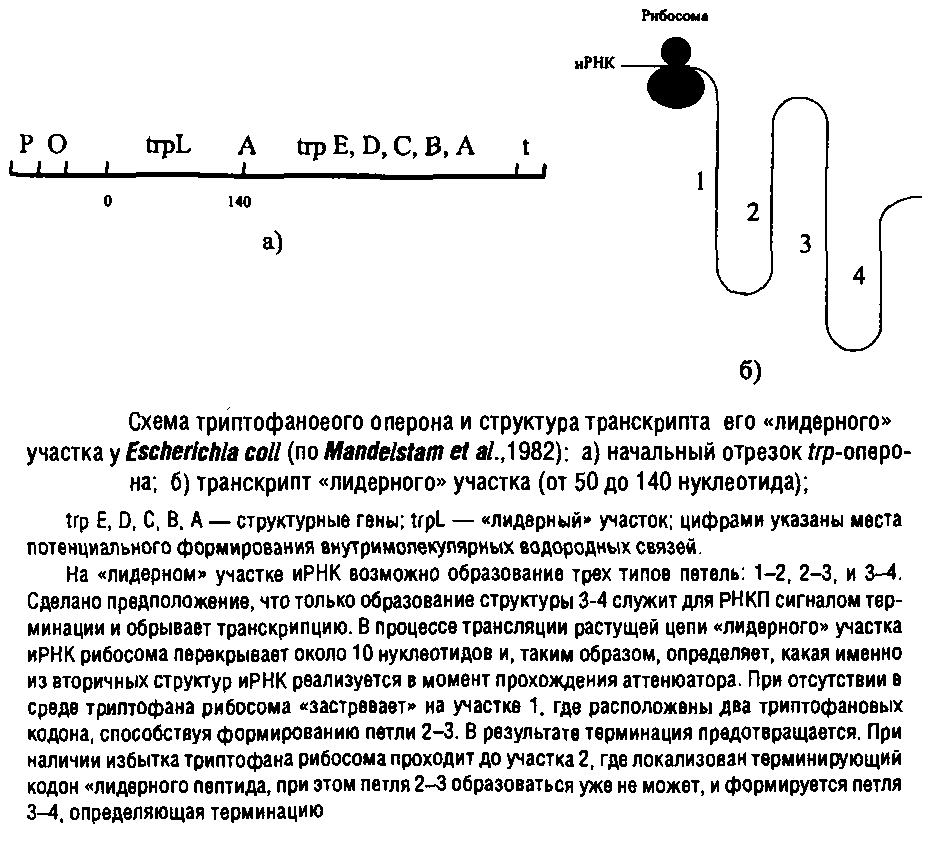
Триптофановый оперон у Escherichia coli регулируется одновременно двумя негативными механизмами: репрессией и аттенюа-цией. При избытке триптофана они действуют аддитивно, практически полностью подавляя транскрипцию. Но даже в отсутствие репрессии аттенюация снижает скорость транскрипции на 90%.
Гистидиновый оперон регулируется только посредством аттенюации. Аналогичная регуляция характерна для синтеза некоторых аминоацил-тРНК-синтетаз и, возможно, ряда экзоферментов. Аттенюатор присутствует также в геноме фага X. В этом случае эффект аттенюатора может быть нейтрализован специальным регуляторным белком антитерминатором. Получены данные о том, что комплекс белка БАК с сАМР также может выполнять функцию антитерминатора, например в опероне, и, таким образом, не только стимулировать инициацию транскрипции, но и предотвращать преждевременную терминацию.
Регуляция транскрипции путем изменения количества активной РНКП. РНКП может служить отрицательным регулятором своего собственного биосинтеза, и при относительном избытке в клетке ее дальнейшее образование тормозится. Координирование регулируется образование с переходом ее в латентное состояние. В определенных условиях латентная РНКП активируется, вызывая повышение скорости транскрипции. По некоторым данным, такая латентная РНКП может составлять до 50% всей РНКП клетки.
7. Регуляция транскрипции путем изменения конформации или структуры ДНК
Одним из способов системной регуляции транскрипции служит изменение степени сверхспирализации ДНК. В этом процессе участвует особый класс ферментов – топоизомеразы,
Релаксирующие топоизомеразы понижают степень сверхспирализации без затраты энергии и принимают участие в инициации репликации. По современной класификации их называют топоизомеразы I
Белки, повышающие степень сверхспирализации ДНК, зависят от АТР и участвуют в репликации ДНК, а также необходимы для осуществления рекомбинаций и конъюгативной передачи генетического материала. Их называют топоизомеразы II.
Косвенным указанием на возможность изменения специфичности транскрипции при изменении степени сверхспирализации ДНК служит тот факт, что в присутствии ингибиторов ДНК-гиразы изменяется спектр синтезируемых белков, хотя сами по себе эти ингибиторы не влияют на процессы транскрипции или трансляции. Например, у Escherichia coii снижается синтез около 20 белков, но одновременно стимулируется синтез других, а образование некоторых белков остается на прежнем уровне.
Перестройка транскрипции при изменении степени сверхспирализации ДНК объясняется по крайней мере двумя причинами. Во-первых, может изменяться специфичность узнавания промоторов РНК-полимеразой. Во-вторых, изменение сверхспирализованности должно приводить к изменению силы взаимодействия с ДНК регуляторных белков.
Поскольку АТР является необходимым компонентом для работы ДНК-гиразы, должна существовать связь степени сверхспирализации ДНК с энергетическим зарядом клетки, что открывает еще одну возможность для регуляции транскрипции.
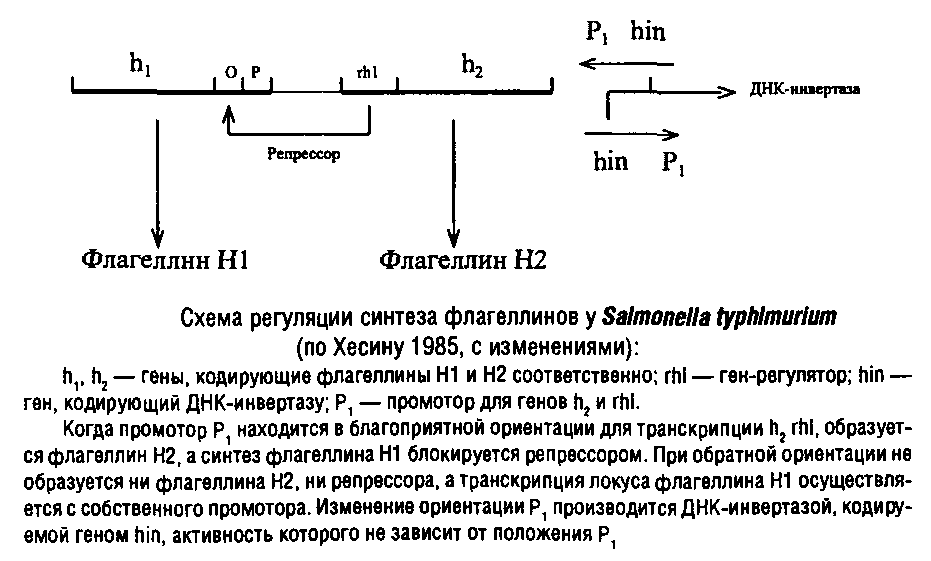
Принципиально иной способ регуляции может быть осуществлен путем обратимого изменения структуры ДНК за счет выщепления подвижных генетических элементов, а затем встраивания их в другие места генома. При этом может меняться характер регуляции транскрипции как генетических локусов, входящих в состав перемещаемых участков, так и соседних с ними локусов. Возможно также встраивание генетического элемента в тот же участок ДНК, откуда он был выщеплен, но в инвертированном виде. Такая инверсия используется иногда как способ регуляции развития фагов, а также образования жгутиковых антигенов у сальмонелл.